
Monoterpene sind eine bedeutende Klasse flüchtiger Isoprenoide, die wesentlich zur Aromatik von Weintrauben und Weinen beitragen. Sie sind insbesondere in aromatischen Rebsorten wie Muskateller, Gewürztraminer und Riesling von Bedeutung, während neutrale Sorten wie Chardonnay oder Pinot Noir geringere Konzentrationen aufweisen (Mateo & Jiménez, 2000; Darriet et al., 2012). Die Hauptmonoterpene in Weintrauben umfassen Geraniol, Linalool, Nerol, Citronellol, α-Terpineol und Rosenoxide, die blumige, fruchtige und zitrusartige Aromen vermitteln (Matarese et al., 2014; Siebert et al., 2018).
Die Synthese von Monoterpenen erfolgt über den Desoxy-Xylulose-5-Phosphat/Methyl-Erythritol-4-Phosphat-Weg (DOXP/MEP) in Plastiden (Luan & Wüst, 2002; Schwab & Wüst, 2015). In der Rebe werden diese Verbindungen vorwiegend in der Beerenhaut synthetisiert, wobei Linalool und Rosenoxid auch im Mesokarp gebildet werden (Gunata et al., 1985). Die Monoterpenakkumulation verläuft je nach Sorte unterschiedlich: Während sie in vielen Rebsorten bereits während der frühen Fruchtentwicklung hoch ist und später abnimmt, zeigen einige Sorten eine verstärkte Produktion im späten Reifestadium (Rienth et al., 2021; Coelho et al., 2007). Monoterpene liegen in Trauben sowohl in freier als auch in glykosylierter Form vor. Während freie Monoterpene direkt zur sensorischen Wahrnehmung beitragen, dienen glykosylierte Monoterpene als Aroma-Vorläufer, die während der Weinherstellung durch enzymatische oder chemische Prozesse hydrolysiert werden (Park et al., 1991; Black et al., 2015). Glykosylierte Terpene können durch UDP-Glucosyltransferasen wie VvGT7, VvGT14 und VvGT15 modifiziert werden, wodurch ihre flüchtige Aktivität während der Reife beeinflusst wird (Bönisch et al., 2014a,b; Zhang et al., 2017).
Sonnenlicht ist einer der wichtigsten Faktoren für die Monoterpenbiosynthese. Die Expression von Terpensynthase-Genen wie VviDXS und VviPNLinNer1 wird durch Licht gefördert, während eine UV-B-Abschwächung die Linalool-Konzentration reduziert (Sasaki et al., 2016; Zhang et al., 2017). In Tempranillo-Trauben führte eine erhöhte UVB-Exposition zur Hochregulation von Linalool- und Cineol-Synthasen (Carbonell-Bejerano et al., 2014). Ein moderater UV-Einfluss fördert die Terpenbildung, während eine übermäßige UV-Strahlung oder vollständiger Lichtausschluss zu einem Rückgang führt (Martínez-Gil et al., 2013; Liu et al., 2024). Eine Beschattung verringert insbesondere glykosylierte Terpene, während Methoxypyrazin-Konzentrationen steigen, was sich negativ auf die aromatische Qualität auswirken kann (Marais et al., 1999; Bureau et al., 2000; Feng et al., 2012).
Die Terpenakkumulation zeigt ein breites Temperatur-Optimum zwischen 10 °C und 20 °C, wobei Temperaturen über 35 °C die Emission steigern, gleichzeitig aber auch zur Verflüchtigung beitragen (Loreto & Schnitzler, 2010; Marais et al., 2001). Übermäßige Hitze (>45 °C) kann die Monoterpensynthese jedoch stark hemmen (Mongélard et al., 2011). Wasserverfügbarkeit beeinflusst die Terpenproduktion ebenfalls erheblich. Während ein leichtes Wasserdefizit das Aromapotenzial erhöhen kann, indem es Monoterpenkonzentrationen steigert (Des Gachons et al., 2005; Savoi et al., 2016), kann starker Wasserstress die Terpenbildung limitieren.
Das Genom von Pinot Noir enthält mehrere funktionale Terpensynthase-Gene (VvTPS), jedoch produziert die Sorte nur geringe Mengen an aromatischen Monoterpenen (Fang & Qian, 2005). Dies deutet darauf hin, dass diese Gene primär für die Synthese nicht-aromatischer Terpene oder anderer pflanzlicher Verteidigungsstoffe genutzt werden (Martin et al., 2010). Obwohl Pinot Noir zu den neutralen Sorten gehört, könnten Monoterpene dennoch eine Rolle in der Aromawahrnehmung spielen. Durch säurekatalysierte Reaktionen können Monoterpene wie Linalool und Geraniol in Verbindungen wie 1,8-Cineol umgewandelt werden, die frische und minzige Noten in gereiften Rotweinen vermitteln (Slaghenaufi & Ugliano, 2018).
Die enzymatische Modifikation, insbesondere durch Cytochrom P450, trägt zur Vielfalt und Komplexität der Monoterpen-Zusammensetzung in Wein bei (Schwab & Wüst, 2015; Ilc et al., 2016). Cytochrom P450 katalysiert unter anderem die Umwandlung von Linalool in (E)-8-Carboxylinalool, aus dem während der Weingärung ein Weinlacton entsteht, ein wichtiger Geruchsstoff von Gewürztraminer-Weinen (Ilc et al., 2017).
In der vorliegenden Studie wurde die Entwicklung der gebundenen und freien Monoterpene in den Rebsorten Pinot Noir und Blaufränkisch während der Traubenreife untersucht. Zu diesem Zweck wurden repräsentative Traubenproben in definierten Entwicklungsstadien entnommen und hinsichtlich ihres Monoterpengehalts analytisch charakterisiert. Ziel der vorliegenden Studie war es, die Dynamik der Monoterpenakkumulation während der Traubenreife bei ausgewählten Rotweinsorten zu untersuchen. Es wurde angenommen, dass die Konzentration an Monoterpenen im Reifeverlauf zunimmt, jedoch nicht linear verläuft. Zudem wurde die Hypothese geprüft, dass Monoterpene größtenteils in glykosidisch gebundener Form vorliegen, während der Anteil freier Monoterpene vergleichsweise gering ist (Bönisch et al., 2014a; Park et al., 1991). Während zur Rebsorte Pinot Noir bereits Untersuchungen zur Monoterpenzusammensetzung in Trauben vorliegen (z. B. Yuan & Qian, 2016), fehlen vergleichbare Daten bislang für Blaufränkisch. Darüber hinaus ist die Forschung zu Monoterpenen in Rotweinrebsorten insgesamt noch begrenzt, da diese Aromastoffe primär mit aromatischen Weißweinsorten assoziiert werden. Die Ergebnisse dieser Untersuchung sollen daher einen Beitrag zur besseren Charakterisierung des Terpenstoffwechsels in roten Rebsorten leisten und potenzielle weinbauliche sowie önologische Implikationen aufzeigen. Ein weiteres Ziel war die methodische Adaptierung einer etablierten GC-MS-Analytik zur präzisen Quantifizierung von Monoterpenen, wobei die bestehende Probenvorbereitung um die Zugabe eines Antioxidans ergänzt wurde, um oxidativen Abbau sowohl während der Extraktion als auch während der Lagerung bei −26 °C zu minimieren.
Die für diese Untersuchung verwendeten Weingärten befinden sich in den österreichischen Weinbaugebieten Mittelburgenland (Blaufränkisch) und Thermenregion (Pinot Noir). Die durchgeführten Erhaltungsmaßnahmen wie Rebschnitt, Einstricken, Entblätterung und Unterstockpflege orientierten sich an den betriebsüblichen Praktiken, wobei keine Traubenausdünnung vorgenommen wurde. Darüber hinaus erfolgte der Pflanzenschutz gemäß integrierter Methoden. Weitere Details zu den Versuchsweingärten sind in Tab. 1 aufgeführt.
Charakterisierung der Versuchsweingärten
| Standort | Lutzmannsburg, Riede Alt Satz | Horitschon, Riede Laupil | Guntramsdorf, Riede Eichkogel | Oberwaltersdorf, Riede Kräutergarten |
|---|---|---|---|---|
| Rebsorte | Blaufränkisch | Pinot Noir | ||
| Koordinaten | 47°27′02″N | 47°34′52″N | 48°03′36″N | 47°58′02″N |
| 16°38′45″O | 16°33′11″O | 16°17′25″O | 16°19′51″O | |
| Pflanzjahr | 2003 | 1990 | 2000 | 2000 |
| Unterlage | Kober 5BB | Fercal | ||
| Klon | Selektion IBY | Dijon 115 | FR 52/86 | |
| Seehöhe | 261 m | 243 m | 295 m | 211 m |
| Reihenausrichtung | Nord-Süd | Nord-Süd | Nord-Süd | Ost-West |
An den Versuchsstandorten standen keine eigenen Wetterstationen zur Verfügung. Auf die Einbindung externer Klimadaten (z. B. von nahegelegenen Wetterstationen) wurde verzichtet, da diese aufgrund topographischer und mikroklimatischer Gegebenheiten nicht als repräsentativ für die jeweiligen Standorte angesehen wurden.
Die Daten der einzelnen Traubenproben-Entnahmen sind in Tab. 2 zusammengefasst. Die Proben wurden annähernd im Zwei-Wochen-Intervall entnommen. Um eine Wiederbeprobung der gleichen Rebstöcke zu vermeiden und so einen Ausdünnungseffekt zu verhindern, wurden bereits beprobte Rebstöcke bei jedem Entnahmezeitpunkt mit unterschiedlich farbigen Bändern markiert. Die Probenahme erfolgte zunächst getrennt von der Morgen- und Nachmittagssonnenseite der Reben, um potenzielle Unterschiede in der Sonnenexposition berücksichtigen zu können. Aufgrund eingeschränkter Ressourcen im analytischen Verarbeitungsschritt wurden die Trauben beider Seiten jedoch nach der Entnahme für die weitere Analyse gepoolt.
Zeitpunkte der Traubenentnahme (VR = Vorreife; NR = Normalreife; ÜR = Überreife)
| Rebsorte | Standort | Zeitpunkte | ||
|---|---|---|---|---|
| Blaufränkisch | Lutzmannsburg | 23.07.2023 | 13.08.2023 | 25.08.2023 |
| 15.09.2023 VR | 29.09.2023 NR | 13.10.2023 ÜR | ||
| Horitschon | 25.08.2023 | |||
| 15.09.2023 VR | 29.09.2023 NR | 13.10.2023 ÜR | ||
| Pinot Noir | Guntramsdorf | 30.07.2023 | 13.08.2023 | 25.08.2023 |
| 08.09.2023 VR | 22.09.2023 NR | 06.10.2023 ÜR | ||
| Oberwaltersdorf | 25.08.2023 | |||
| 08.09.2023 VR | 22.09.2023 NR | 06.10.2023 ÜR | ||
Die Auswahl der Rebstöcke und Trauben erfolgte unter Berücksichtigung spezifischer Ausschlusskriterien, die eine repräsentative Stichprobe sicherstellen sollten:
1) Gesundheitszustand der Rebstöcke: Es wurden ausschließlich Rebstöcke ohne Anzeichen von ESCA oder anderen Rebkrankheiten wie Bois noir ausgewählt.
2) Rebsortenreinheit und phänologische Entwicklung: Die Zugehörigkeit der Reben zur Sorte Blaufränkisch und Pinot Noir sowie die phänologische Repräsentativität der Trauben wurden verifiziert.
3) Ort der Probenahme: Die Reben der ersten Pfahl-zu-Pfahl-Abstände jeder Rebzeile, sowie die Reben der beiden äußersten Rebzeilen zur Gänze wurden von der Probenahme ausgeschlossen.
4) Proportionalität der Traubenprobenahme: Die Entnahme der Trauben erfolgte im Verhältnis von 1:1 zwischen basalen und distalen Trauben.
Durch die strikte Anwendung dieser Kriterien wurde die Repräsentativität der Proben maximiert. Für die Probenahme wurden sechs Trauben von der Morgensonnenseite und sechs von der Nachmittagssonnenseite getrennt entnommen. Diese Proben wurden bei −70 °C durch CO2(s) indirekt tiefgefroren (FA Linde, Wien, Österreich), um biochemische Reaktionen in den Beeren zu verhindern und ihre chemische Zusammensetzung zu bewahren. Im Anschluss wurden die Trauben beider Seiten gepoolt und in geeigneten Beuteln vakuumversiegelt. Es wurde festgestellt, dass das CO2(s) die Stiele der Trauben so stark spröde machte, dass sich die Beeren nahezu gänzlich von den Stielen lösten, was dazu beitrug, die Proben zusätzlich in ihrer Repräsentativität zu steigern. Die vakuumversiegelten und gepoolten Beeren wurden schließlich bei −26 °C bis zur chromatographischen Analyse zwischengelagert.
Die Quantifizierung der freien und gesamten Monoterpene fand in der Abteilung Chemie-Forschung und Isotopenanalyse der HBLAuBA für Wein- und Obstbau Klosterneuburg statt. Die Traubenproben wurden zwischen Mai 2024 und Juli 2024 analysiert.
Zur Quantifizierung der freien und gesamten Monoterpene wurde eine repräsentative Beerenprobe aus 20 Einzelbeeren mittels flüssigem Stickstoff versprödet, und 5,0 g des resultierenden Pulvers in ein 20-mL-Glasfläschchen überführt. Zu der Probe wurden 5,0 mL 80 % Methanol sowie 50,0 μL des Hydrolysekontrollstandards Octyl-β-D-glucopyranosid (Zielkonzentration 1-Octanol = 445,41 μg/L; Sigma-Aldrich, St. Louis, USA) hinzugefügt. Der Kontrollstandard diente ausschließlich der Kontrolle der Ausbeute der Hydrolyse, die Quantifizierung der freien Monoterpene wurde nicht tangiert. Zum Schutz vor Oxidation wurde der Probe 20 μL einer Butylhydroxyanisol (BHA)-Lösung (Carl Roth, Karlsruhe, Deutschland; Konzentration: 125 mg/10 mL Ethanol) hinzugefügt. Nach einem 1-minütigen Vortex-Vorgang wurde die Probe für 4 Stunden auf einem Laborschüttler (Typ KS 15 B, VWR, Darmstadt, Deutschland) extrahiert und anschließend bei 6000 U/min für 10 min zentrifugiert. Insgesamt wurden je Standort und Zeitpunkt drei biologische Replikate angefertigt. Bis zur chromatographischen Analyse wurden die methanolischen Extrakte bei −26 °C tiefgefroren.
Zur Bestimmung der freien Monoterpene wurde 1,0 mL des methanolischen Extrakts im Verhältnis 1:10 mit McIlvaine-Puffer (pH 5) verdünnt. Für die Bestimmung der gesamten Monoterpene wurden 2,5 mL des Extrakts mit McIlvaine-Puffer auf ein Endvolumen von 25,0 mL (1:10) aufgefüllt, anschließend 14 mg Rapidase® Revelation Aroma (DSM Food & Beverage, Leeuwarden, Niederlande) zugesetzt und die Probe für 32 Stunden bei 35 °C enzymatisch hydrolysiert. Der Gasraum der Messkolben wurde unmittelbar nach der Verdünnung unter Zugabe von Stickstoff (N2) inertisiert.
Für die Analyse der freien und gesamten Monoterpene wurde ein 7890A Gaschromatograph (GC) von Agilent Technologies in Kombination mit einem 5975C Inert Massenspektrometer (MSD) mit Triple Axis Detector sowie einem Autosampler von CTC Analytics verwendet. Zur Trennung der Analyten kam eine ZB-5MS-Gaschromatographiesäule (Länge: 60 m, Innendurchmesser: 0,25 mm, Filmdicke: 0,25 μm) von Phenomenex zum Einsatz.
Die Quantifizierung der freien und gesamten Monoterpene erfolgte gemäß der Methode von Philipp et al. (2020) unter Anwendung einer Headspace-Solid-Phase-Microextraction-Gaschromatographie-SIM-Massenspektrometrie (HS-SPME-GC-SIM-MS) Technik. Dabei wurde eine 65 μm Polydimethylsiloxan/Divinylbenzol (PDMS/DVB)-beschichtete Faser (SUPELCO, Bellefonte, PA, USA) verwendet.
Für die Probenvorbereitung wurden 1,5 g Natriumchlorid (NaCl), 5 mL der Probe und 10 μL eines internen Standards (3,4-Dimethylanisol in Methanol, nominale Konzentration: 35 μg/L) in ein 20-mL-Headspace-Fläschchen gegeben. Nach einer kurzen Vorinkubation von 0,1 min wurde die Probe bei 50 °C für 30 min unter kontinuierlichem Rühren (500 U/min) extrahiert.
Nach der Extraktion wurde die Faser im „splitless“-Verfahren in den Gaschromatographen injiziert. Die Desorption der extrahierten Substanzen erfolgte bei 250 °C über eine Dauer von fünf min. Die Injektionstemperatur wurde auf 250 °C eingestellt, und Helium wurde als Trägergas mit einer Flussrate von 1,2 mL/min verwendet.
Das Ofenprogramm begann bei 50 °C und wurde für 3 min gehalten. Danach folgte eine lineare Erhöhung der Temperatur mit 1 °C/min bis auf 92 °C (10 min gehalten), gefolgt von einer Erhöhung mit 5 °C/min auf 127 °C und abschließend einer schnellen Erhöhung mit 40 °C/min bis zu einer Temperatur von 260 °C, die für 4,6 min gehalten wurde. Die Transferleitung zum Massenspektrometer wurde auf 250 °C temperiert. Die Analyse der Verbindungen erfolgte im Einzelionenmodus (EI+, 70 eV) mit einer Gesamtlaufzeit von 70 min. Die Massen der Monoterpene können der Arbeit von Philipp et al. (2024) entnommen werden. Der prozentuale Anteil der gebundenen Monoterpene in den Trauben wurden durch die Differenz zwischen den gesamten und den freien Monoterpenen berechnet. Die spezifischen Validierungsparameter sind in Philipp et al. (2024) dokumentiert.
Die statistische Analyse der Daten erfolgte mit SPSS 29.0 (IBM, USA) und XLSTAT 2023.3.1 (Lumivero, Denver, USA). Zunächst wurden die Daten einer logarithmischen Transformation gemäß Leydesdorff & Bensman (2006) unterzogen. Im Anschluss wurde die Normalverteilung der Daten mittels Shapiro-Wilk-Test und die Homogenität der Varianzen mit dem Levene-Test überprüft. Für die Analyse der Varianzen wurde eine einfaktorielle Varianzanalyse (ANOVA) in SPSS 29.0 durchgeführt, wobei ein Signifikanzniveau von p < 0,05 zugrunde gelegt wurde. Signifikante Unterschiede wurden mittels Tukey-B-Post-hoc-Test ermittelt. Zur Visualisierung der Konzentrationsverläufe wurde eine Heatmap-Darstellung auf Basis der Mittelwerte (n = 3) je Monoterpen und Erhebungszeitpunkt erstellt. Die Farbcodierung entspricht den absoluten Konzentrationswerten in μg/kg Frischgewicht, wobei niedrigere Konzentrationen rötlich und höhere Konzentrationen grünlich dargestellt sind. Die Farbskala wurde jeweils individuell auf den Konzentrationsbereich des jeweiligen Monoterpens (xmin–xmax) bezogen. In Fällen, in denen die Differenz zwischen minimalem und maximalem Konzentrationswert eines Monoterpens ≤ 1 μg/kg Frischgewicht betrug, wurden alle zugehörigen Zellen einheitlich grau eingefärbt, um eine visuelle Überbetonung geringfügiger Unterschiede zu vermeiden. Zusätzlich wurden die exakten Mittelwerte direkt in die Felder der Heatmap integriert, um eine transparente und quantitativ nachvollziehbare Darstellung zu gewährleisten. Statistisch signifikante Unterschiede innerhalb der jeweiligen Monoterpenverläufe sind durch Sternchensymbole gekennzeichnet: *** = p < 0,001 (hochsignifikant), ** = p < 0,01 (signifikant), kein Stern = nicht signifikant. Ergänzend sind die entsprechenden Mittelwerte in tabellarischer Form im Anhang dokumentiert.
In dieser Studie wurde die Entwicklung der freien und gesamten Monoterpene in den Rebsorten Blaufränkisch und Pinot Noir während der Beerenentwicklung untersucht. Ziel war es, die zeitliche Dynamik der Monoterpenakkumulation zu erfassen, die Verteilung zwischen freien und gebundenen Formen zu differenzieren sowie mögliche Standorteffekte zu identifizieren. Es wurde hypothesiert, dass die Monoterpenkonzentrationen im Verlauf der Reife nicht linear, jedoch insgesamt ansteigen. Zudem wurde angenommen, dass Monoterpene überwiegend in gebundener Form in den Trauben vorliegen und nur ein geringer Anteil als freie Monoterpene nachweisbar ist. Da bisher nur wenige Studien zur Monoterpenzusammensetzung in Rotweinrebsorten, insbesondere für Blaufränkisch, vorliegen, sollte diese Untersuchung zur besseren Charakterisierung des Terpenstoffwechsels in roten Rebsorten beitragen.
Im Kontext der Reifeentwicklung ist zu berücksichtigen, dass sich das Verhältnis von Beerenoberfläche zu Fruchtmasse verändert, was die Konzentration schalenassoziierter Verbindungen wie Monoterpenen potenziell beeinflussen kann. In der vorliegenden Studie erfolgte – analog zur Vorgehensweise von Yuan & Qian (2016) – die Probenaufarbeitung auf Frischmassebasis, ohne explizite Korrektur nach Beerenoberfläche.
Die Ergebnisse zu den quantifizierten Monoterpenen an den Blaufränkisch-Standorten Lutzmannsburg (Riede Alt Satz) und Horitschon (Riede Lauspil) werden in Abb. 1 und Abb. 2 als Heatmaps visualisiert und in Tab. 3 und Tab. 4 im Anhang durch Mittelwerttabellen ergänzt.
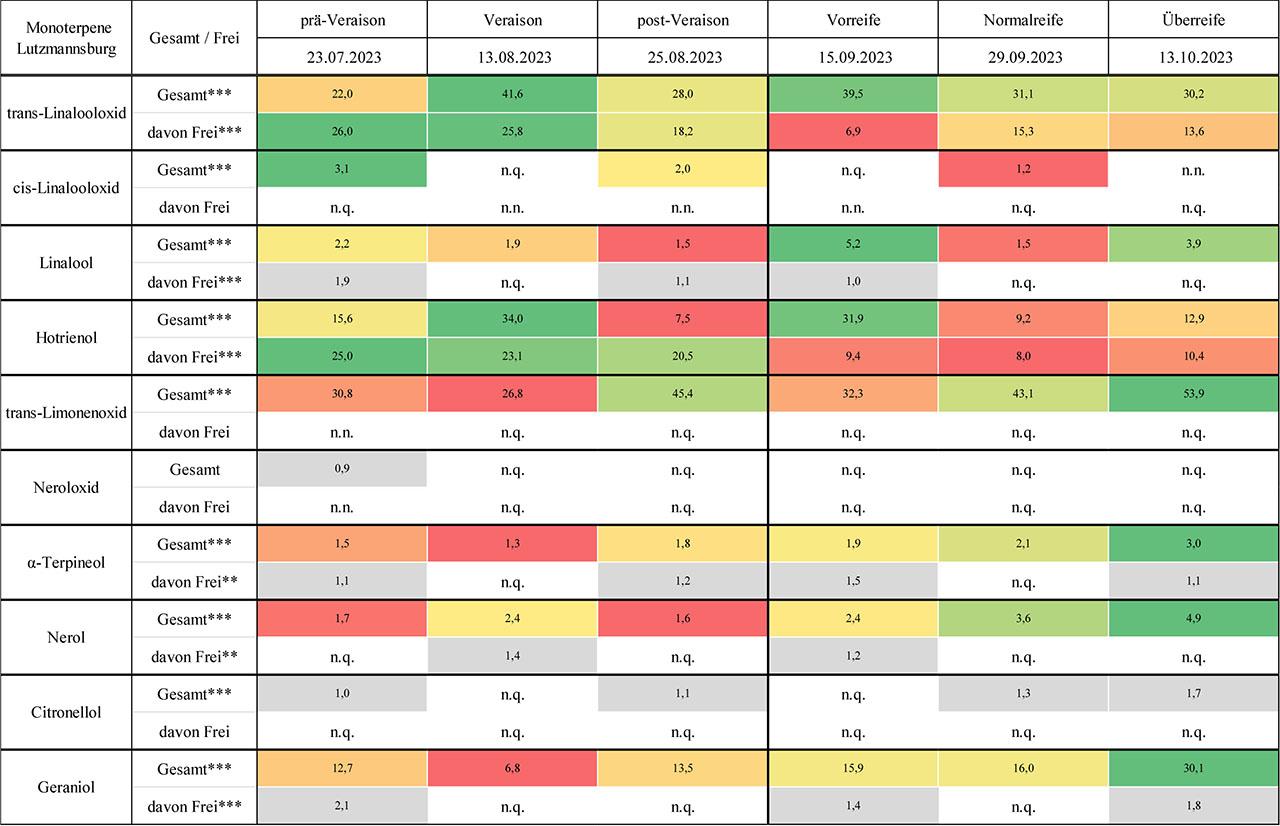
Konzentration [µg/kg Frischgewicht; n =3] und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Blaufränkisch-Trauben am Standort Lutzmannsburg, Riede Alt Satz („n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Hochsignifikante Unterschiede zwischen den Zeitpunkten sind mit *** und signifikante Unterschiede mit ** gekennzeichnet.)
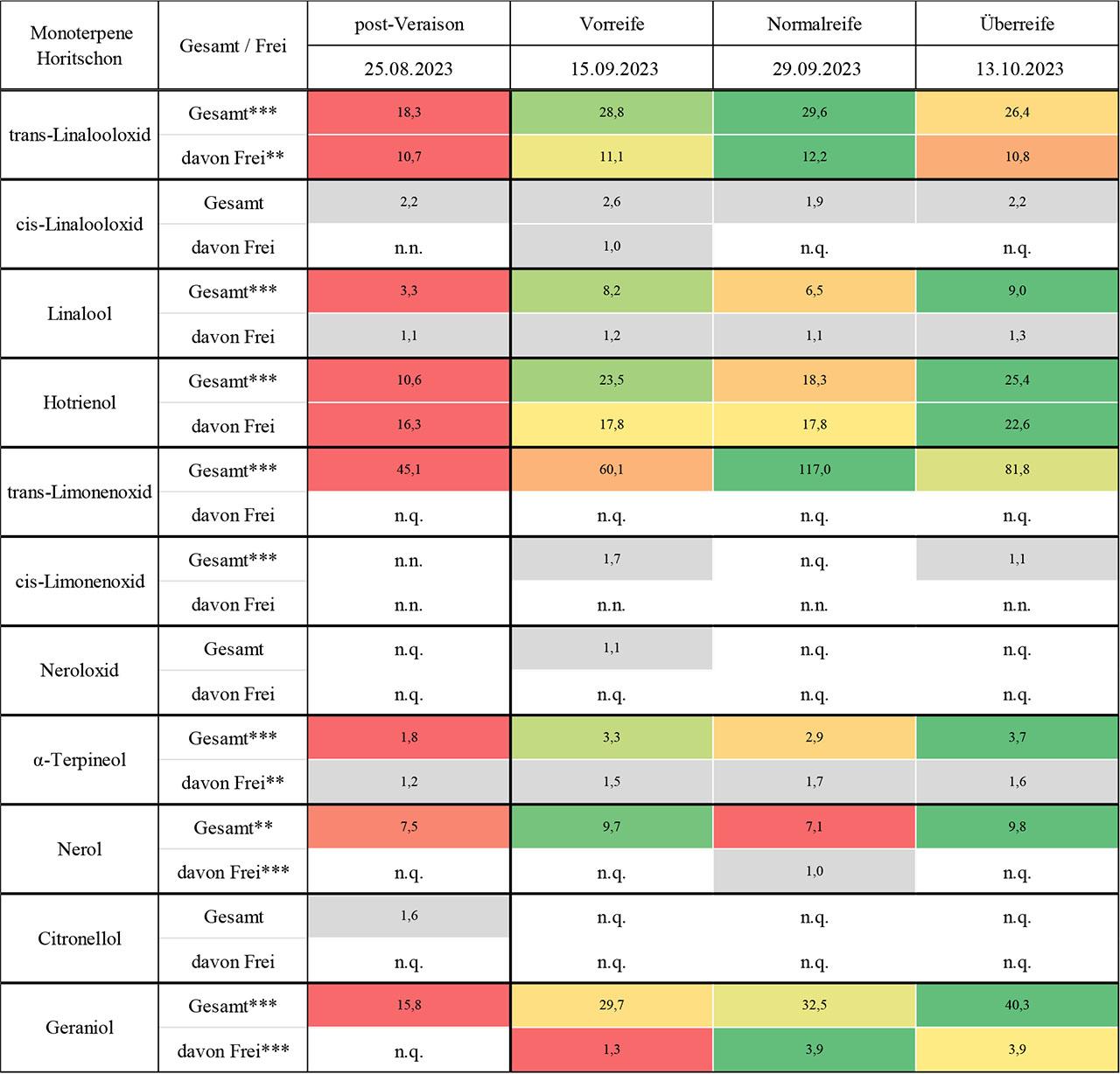
Konzentration [µg/kg Frischgewicht; n =3] und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Blaufränkisch-Trauben am Standort Horitschon, Riede Lauspil („n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Hochsignifikante Unterschiede zwischen den Zeitpunkten sind mit *** und signifikante Unterschiede mit ** gekennzeichnet.)
Konzentration und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Blaufränkisch-Trauben am Standort Lutzmannsburg, Riede Alt Satz
| µg/kg Frischgewicht | Blaufränkisch Lutzmannsburg, Riede Alt Satz | ||||||
|---|---|---|---|---|---|---|---|
| 23.07.2023 | 13.08.2023 | 25.08.2023 | 15.09.2023 Vorreife | 29.09.2023 Normalreife | 13.10.2023 Überreife | ||
| trans-Linalooloxid | Fr. | 26,0c ± 2,3 | 25,8c ± 2,7 | 18,2b ± 3,3 | 6,9a ± 1,0 | 15,3b ± 1,4 | 13,6b ± 0,9 |
| Ges. | 22,0a ± 5,2 | 41,6c ± 4,6 | 28,0a ± 1,4 | 39,5bc ± 4,7 | 31,1bc ± 2,2 | 30,2ab ± 1,4 | |
| Geb. | 0% | 38% | 35% | 82% | 51% | 55% | |
| cis-Linalooloxid | Fr. | n.q. | n.n. | n.n. | n.n. | n.q. | n.q. |
| Ges. | 3,1c ± 0,6 | n.q. | 2,0b ± 0,3 | n.q. | 1,2a ± 0,2 | n.n. | |
| Geb. | 100% | - | 100% | - | 100% | - | |
| Linalool | Fr. | 1,9b ± 0,2 | n.q. | 1,1a ± 0,1 | 1,0a ± 0,1 | n.q. | n.q. |
| Ges. | 2,2b ± 0,5 | 1,9ab ± 0,1 | 1,5a ± 0,1 | 5,2d ± 0,3 | 1,5a ± 0,3 | 3,9c ± 0,1 | |
| Geb. | 14% | 100% | 25% | 80% | 100% | 100% | |
| Hotrienol | Fr. | 25,0b ± 2,3 | 23,1b ± 2,5 | 20,5b ± 1,9 | 9,4a ± 1,5 | 8,0a ± 1,1 | 10,4a ± 1,3 |
| Ges. | 15,6b ± 3,9 | 34,0c ± 3,3 | 7,5a ± 1,3 | 31,9c ± 2,6 | 9,2a ± 0,5 | 12,9b ± 1,8 | |
| Geb. | 0% | 32% | 0% | 70% | 13% | 20% | |
| cis-Rosenoxid | Fr. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. | - | - | - | - | - | - | |
| trans-Rosenoxid | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. | |
| Geb. | - | - | - | - | - | - | |
| trans-Limonenoxid | Fr. | n.n. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 30,8a ± 4,5 | 26,8a ± 5,0 | 45,4b ± 7,8 | 32,3a ± 3,1 | 43,1b ± 0,2 | 53,9b ± 3,0 | |
| Geb. | 100% | 100% | 100% | 100% | 100% | 100% | |
| cis-Limonenoxid | Fr. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. | - | - | - | - | - | - | |
| Neroloxid | Fr. | n.n. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 0,9 ± 0,2 | n.q. | n.q. | n.q. | n.q. | n.q. | |
| Geb. | 100% | - | - | - | - | - | |
| Lavandulol | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.q. | n.n. | n.q. | n.n. | n.n. | |
| Geb. | - | - | - | - | - | - | |
| α-Terpineol | Fr. | 1,1a ± 0,2 | n.q. | 1,2ab ± 0,2 | 1,5b ± 0,2 | n.q. | 1,1ab ± 0,1 |
| Ges. | 1,5ab ± 0,3 | 1,3a ± 0,1 | 1,8bc ± 0,1 | 1,9bc ± 0,2 | 2,1c ± 0,1 | 3,0d ± 0,2 | |
| Geb. | 28% | 100% | 35% | 22% | 100% | 61% | |
| γ-Terpineol | Fr. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. | - | - | - | - | - | - | |
| Nerol | Fr. | n.q. | 1,4b ± 0,2 | n.q. | 1,2ab ± 0,2 | n.q. | n.q. |
| Ges. | 1,7a ± 0,4 | 2,4b ± 0,2 | 1,6a ± 0,1 | 2,4b ± 0,5 | 3,6c ± 0,5 | 4,9c ± 0,8 | |
| Geb. | 100% | 40% | 100% | 47% | 100% | 100% | |
| Citronellol | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 1,0ab ± 0,0 | n.q. | 1,1ab ± 0,2 | n.q. | 1,3b ± 0,1 | 1,7c ± 0,3 | |
| Geb. | 100% | - | 100% | - | 100% | 100% | |
| Geraniol | Fr. | 2,1c ± 0,3 | n.q. | n.q. | 1,4ab ± 0,3 | n.q. | 1,8bc ± 0,2 |
| Ges. | 12,7b ± 1,8 | 6,8a ± 0,3 | 13,5b ± 0,9 | 15,9b ± 2,2 | 16,0b ± 0,9 | 30,1c ± 2,6 | |
| Geb. | 84% | 100% | 100% | 91% | 100% | 94% | |
| Σ Monoterpene | Fr. | 56,0d ± 2,1 | 50,3d ± 3,7 | 40,9c ± 4,0 | 21,5a ± 2,2 | 23,3ab ± 0,8 | 26,9b ± 1,2 |
| Ges. | 91,6a ± | 114,7cd ± | 102,4ab ± | 129,0d ± | 109,2bc ± 2,0 | 140,5d ± 2,1 | |
| Geb. | 39% | 56% | 60% | 83% | 79% | 81% | |
Die Zeilen zeigen die quantifizierten Konzentrationen [μg/kg Frischgewicht] der einzelnen Monoterpene (x̄ ± σ), differenziert nach freier, gesamter und gebundener Fraktion. Signifikante Unterschiede zwischen den Zeitpunkten sind durch unterschiedliche Buchstaben gekennzeichnet. „n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Die Nachweisgrenze (LOD) und die Bestimmungsgrenze (LOQ) der analysierten Monoterpene sind in der Studie von Philipp et al. (2024) dokumentiert.
Konzentration und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Blaufränkisch-Trauben am Standort Horitschon, Riede Lauspil
| [µg/kg Frischgewicht] | Blaufränkisch Horitschon, Riede Lauspil | ||||
|---|---|---|---|---|---|
| 25.08.2023 | 15.09.2023 Vorreife | 29.09.2023 Normalreife | 13.10.2023 Überreife | ||
| trans-Linalooloxid | Fr. | 10,7a ± 0,3 | 11,1a ± 0,5 | 12,2b ± 1,3 | 10,8a ± 0,1 |
| Ges. | 18,3a ± 1,1 | 28,8bc ± 0,8 | 29,6c ± 1,5 | 26,4b ± 0,5 | |
| Geb. [%] | 42% | 61% | 59% | 59% | |
| cis-Linalooloxid | Fr. | n.n. | 1,0 ± 0,1 | n.q. | n.q. |
| Ges. | 2,2a ± 0,4 | 2,6a ± 0,4 | 1,9a ± 0,2 | 2,2a ± 0,1 | |
| Geb. [%] | 100% | 60% | 100% | 100% | |
| Linalool | Fr. | 1,1a ± 0,2 | 1,2a ± 0,1 | 1,1a ± 0,0 | 1,3a ± 0,2 |
| Ges. | 3,3a ± 0,2 | 8,2bc ± 1,4 | 6,5b ± 0,5 | 9,0c ± 0,2 | |
| Geb. [%] | 67% | 85% | 83% | 86% | |
| Hotrienol | Fr. | 16,3a ± 1,1 | 17,8a ± 1,8 | 17,8a ± 1,2 | 22,6a ± 4,1 |
| Ges. | 10,6a ± 1,5 | 23,5bc ± 4,3 | 18,3b ± 1,7 | 25,4c ± 1,3 | |
| Geb. [%] | 0% | 24% | 3% | 11% | |
| cis-Rosenoxid | Fr. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| trans-Rosenoxid | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.q. | n.q. | n.q. | |
| Geb. [%] | - | - | - | - | |
| trans-Limonenoxid | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 45,1a ± 3,4 | 60,1b ± 2,4 | 117,0d ± 20,3 | 81,8c ± 8,7 | |
| Geb. [%] | 100% | 100% | 100% | 100% | |
| cis-Limonenoxid | Fr. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | 1,7b ± 0,2 | n.q. | 1,1a ± 0,1 | |
| Geb. [%] | - | 100% | - | 100% | |
| Neroloxid | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | 1,1 ± 0,1 | n.q. | n.q. | |
| Geb. [%] | - | 100% | - | - | |
| Lavandulol | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| α-Terpineol | Fr. | 1,2a ± 0,2 | 1,5b ± 0,1 | 1,7b ± 0,2 | 1,6ab ± 0,3 |
| Ges. | 1,8a ± 0,1 | 3,3c ± 0,2 | 2,9b ± 0,2 | 3,7c ± 0,1 | |
| Geb. [%] | 32% | 54% | 41% | 57% | |
| γ-Terpineol | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| Nerol | Fr. | n.q. | n.q. | 1,0 ± 0,1 | n.q. |
| Ges. | 7,5a ± 0,6 | 9,7b ± 1,7 | 7,1a ± 0,5 | 9,8b ± 0,3 | |
| Geb. [%] | 100% | 100% | 87% | 100% | |
| Citronellol | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 1,6 ± 0,2 | n.q. | n.q. | n.q. | |
| Geb. [%] | 100% | - | - | - | |
| Geraniol | Fr. | n.q. | 1,3a ± 0,2 | 3,9b ± 0,3 | 3,9b ± 0,5 |
| Ges. | 15,8a ± 2,8 | 29,7b ± 1,5 | 32,5bc ± 0,9 | 40,3c ± 2,3 | |
| Geb. [%] | 100% | 96% | 88% | 90% | |
| Σ Monoterpene | Fr. | 29,2a ± 0,8 | 34,0ab ± 2,4 | 37,7b ± 2,6 | 40,2b ± 4,6 |
| Ges. | 106,1a ± 7,2 | 168,7ab ± 5,5 | 131,3ab ± 55,4 | 199,7b ± 8,5 | |
| Geb. [%] | 72% | 80% | 71% | 80% | |
Die Zeilen zeigen die quantifizierten Konzentrationen [μg/kg Frischgewicht] der einzelnen Monoterpene (x̄ ± σ), differenziert nach freier, gesamter und gebundener Fraktion. Signifikante Unterschiede zwischen den Zeitpunkten sind durch unterschiedliche Buchstaben gekennzeichnet. „n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Die Nachweisgrenze (LOD) und die Bestimmungsgrenze (LOQ) der analysierten Monoterpene sind in der Studie von Philipp et al. (2024) dokumentiert.
Die beiden Linalooloxid-Isomere trans- und cis-Linalooloxid zeigten in der Rebsorte Blaufränkisch deutlich unterschiedliche Konzentrationsverläufe zwischen den Standorten Lutzmannsburg und Horitschon. Während trans-Linalooloxid an beiden Standorten regelmäßig nachgewiesen wurde, wies Lutzmannsburg eine wesentlich dynamischere Entwicklung auf. Die freie Fraktion wies zu Beginn vergleichsweise hohe Werte auf, fiel zur Vorreife deutlich ab und verblieb anschließend auf einem niedrigeren Niveau. Die Gesamtfraktion folgte keinem linearen Muster, sondern zeigte ein unregelmäßiges Profil mit mehreren Peaks. In Horitschon verlief die Entwicklung von trans-Linalooloxid hingegen gleichmäßiger: Die freie Fraktion blieb auf konstant niedrigem Niveau, während die Gesamtfraktion kontinuierlich bis zur Normalreife anstieg und erst zur Überreife leicht zurückging. Diese Unterschiede lassen sich möglicherweise auf das Rebalter und kleinklimatische Faktoren zurückführen: Die jüngere Rebanlage in Lutzmannsburg könnte für die größere Volatilität der Monoterpenverläufe verantwortlich sein (Lin & Cantu, 2019; Riffle et al., 2022), während das potenziell leicht kühlere Mikroklima dort zusätzliche Enzymaktivität beeinflusst haben könnte (Bönisch et al., 2014a; Bönisch et al., 2015). In beiden Lagen dominierte die gebundene Fraktion über den gesamten Verlauf hinweg, was mit bisherigen Erkenntnissen zur glykosidischen Speicherung oxidierter Monoterpene übereinstimmt (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991). Das cis-Isomer hingegen trat insgesamt deutlich seltener auf. In Lutzmannsburg war es lediglich zu Beginn nachweisbar, in Horitschon wurde es zur Vorreife einmalig in freier Form bestimmt. Auch hier dominierte die gebundene Fraktion, wobei die Gesamtwerte deutlich niedriger blieben als beim trans-Isomer. Diese Beobachtung deckt sich mit Literaturangaben, nach denen cis-Linalooloxid in wirtschaftlich relevanten Rebsorten oft nur in Spuren vorkommt (Ferreira & Lopez, 2019; Gunata et al., 1985; Schwab & Wüst, 2015). Die erste Hypothese eines nicht-linearen Konzentrationsverlaufs wurde damit insbesondere für trans-Linalooloxid in Lutzmannsburg klar bestätigt. Die zweite Hypothese zur Dominanz gebundener Formen wurde von beiden Isomeren erfüllt. Ein Standorteffekt zeigte sich ebenfalls, jedoch in isomerspezifischer Weise – mit deutlich ausgeprägteren Differenzen beim trans-Isomer.
Linalool konnte in der Rebsorte Blaufränkisch an beiden Standorten kontinuierlich nachgewiesen werden, wobei sich deutliche Unterschiede im zeitlichen Verlauf sowie in der Nachweisbarkeit der freien Fraktion zeigten. In Lutzmannsburg war die freie Fraktion nur zu einzelnen Zeitpunkten messbar, während sie in Horitschon über alle Reifestadien hinweg durchgehend nachweisbar war. Die Gesamtfraktion in Lutzmannsburg wies ein unregelmäßiges Muster auf, mit einem Rückgang zu Beginn, einem Peak zur Vorreife, einem erneuten Rückgang und einem weiteren Anstieg zur Überreife. In Horitschon hingegen zeigte sich ein gleichmäßiger, kontinuierlicher Anstieg bis zur Überreife, wobei die gebundene Fraktion von Beginn an dominierte und sich stetig erhöhte. Diese Unterschiede könnten auf verschiedene Standortfaktoren zurückzuführen sein: Die ältere Rebanlage in Horitschon spricht für eine konstantere Aromaausprägung (Riffle et al., 2022), während in Lutzmannsburg möglicherweise stärkere äußere Einflüsse – wie Temperaturschwankungen oder UV-Belastung – eine volatilere Terpenentwicklung begünstigt haben (Lin & Cantu, 2019; Zhang et al., 2017; Song et al., 2015). Linalool ist besonders empfindlich gegenüber Licht- und Temperatureinflüssen, wobei moderate UV-Strahlung die Synthese fördern kann (Gil et al., 2012; Sasaki et al., 2016), während übermäßige Belastung hemmend wirkt (Liu et al., 2024). Die Tatsache, dass die gebundene Fraktion in beiden Lagen dominant war, ist konsistent mit bisherigen Beobachtungen bei nicht-aromatischen Rebsorten (Mateo & Jiménez, 2000; Darriet et al., 2012). Die erste Hypothese eines nicht-linearen Konzentrationsverlaufs wurde insbesondere in Lutzmannsburg bestätigt, während die zweite Hypothese – die Dominanz der gebundenen Form – an beiden Standorten klar erfüllt wurde.
Hotrienol war an beiden Blaufränkisch-Standorten über den gesamten Verlauf der Beerenentwicklung hinweg nachweisbar und stellte im Vergleich zu anderen Monoterpenen eine Besonderheit dar: Die Verbindung trat überwiegend in freier Form auf (70–100 %) und zeigte – im Gegensatz zu den meisten Monoterpenen – nur eine geringe Tendenz zur glykosidischen Bindung. Dementsprechend wurde im Rahmen der Probenvorbereitung – insbesondere während der enzymatischen Hydrolyse – nur sehr wenig Hotrienol freigesetzt. Besonders in Lutzmannsburg zeigte sich ein ausgeprägter, dynamischer Verlauf, mit einem Peak der freien Fraktion zu Beginn der Reife und einem weiteren Maximum der Gesamtfraktion zur Vorreife. Lediglich zu diesem Zeitpunkt war dort ein erheblicher Anteil an gebundenem Hotrienol (70 %) messbar, während die übrigen Zeitpunkte durch eine klare Dominanz der freien Fraktion geprägt waren. In Horitschon hingegen blieb der Hotrienolverlauf konstanter: Sowohl die freie als auch die gesamte Fraktion stiegen kontinuierlich und gleichmäßig an, wobei der gebundene Anteil durchgehend niedrig blieb. Die auffällige Dynamik in Lutzmannsburg könnte durch das jüngere Rebalter und eine damit verbundene höhere Enzymreaktivität auf Umweltreize erklärt werden (Lin et al., 2019; Riffle et al., 2022; Bönisch et al., 2014a; Bönisch et al., 2015). Zudem ist Hotrienol bekanntlich ein Oxidationsprodukt von Linalool (Luan et al., 2005). In Lutzmannsburg könnten stärkere kleinklimatische Schwankungen – z. B. durch höhere Sonneneinstrahlung oder Temperaturvariabilität – diese Umwandlung begünstigt haben (Gil et al., 2012; Skinkis et al., 2010; Song et al., 2015; Zhang et al., 2017). In der Literatur wird Hotrienol zudem häufiger als freie Verbindung beschrieben, was die Beobachtungen in dieser Studie stützt (Bönisch et al., 2014b; Mateo & Jiménez, 2000; Darriet et al., 2012). Die erste Hypothese eines nicht-linearen Konzentrationsverlaufs wurde durch die deutlich unterschiedlichen Kurvenverläufe in Lutzmannsburg bestätigt. Die zweite Hypothese, wonach gebundene Fraktionen dominieren, trifft auf Hotrienol hingegen nicht zu und stellt hier eine relevante Ausnahme dar. Auch ein Standorteffekt zeigte sich: Während Lutzmannsburg eine stärkere Dynamik und deutlichere Peaks aufwies, verlief die Entwicklung in Horitschon konstanter und ruhiger – was den Einfluss des Rebalters wie auch mikroklimatische Effekte in der Terpenbiosynthese unterstreicht (Marais et al., 2001; Riffle et al., 2022; Skinkis et al., 2010; Reynolds & Wardle, 1989).
Die beiden Rosenoxid-Isomere cis- und trans-Rosenoxid konnten in der Rebsorte Blaufränkisch an keinem der beiden untersuchten Standorte in quantifizierbarer Konzentration nachgewiesen werden und sind daher in den Abb. 1 und 2 nicht enthalten. Sowohl in Lutzmannsburg als auch in Horitschon blieben alle Werte – unabhängig vom Reifestadium und von der Fraktionsform – unterhalb der analytischen Nachweisgrenze. Entsprechend war keine Differenzierung zwischen den Standorten oder eine Beobachtung eines Reifeverlaufes möglich. Die Abwesenheit von Rosenoxid ist im Kontext der Rebsorte Blaufränkisch nicht überraschend: Als Vertreter der sogenannten neutralen Rebsorten enthält Blaufränkisch im Vergleich zu aromatischen Sorten wie Muskateller oder Gewürztraminer deutlich geringere Mengen an Monoterpenen insgesamt (Mateo & Jiménez, 2000; Gunata et al., 1985; Reynolds & Wardle, 1989). Rosenoxid, das über Citronellol biosynthetisiert wird, tritt vor allem in stark aromatischen Weißweinsorten auf, während es in Rotweinsorten entweder gar nicht oder nur in Spuren detektiert wurde (Ferreira & Lopez, 2019; Gunata et al., 1985; Schwab & Wüst, 2015). Dass Citronellol in den vorliegenden Proben ebenfalls nur in sehr geringen Mengen nachweisbar war, unterstützt diese Einschätzung. Auch genetische und epigenetische Faktoren könnten eine Rolle spielen: Studien zeigen, dass sowohl die Expression der notwendigen Enzyme als auch die Glykosylierungsaktivität (z. B. durch VvGT14) stark sorten- und klonabhängig sind (Bönisch et al., 2014a; Li et al., 2017). Die beiden Hypothesen der Studie konnten in Bezug auf Rosenoxid nicht überprüft werden, da mangels quantifizierbarer Werte weder ein Verlauf noch eine Fraktionsverteilung beobachtbar war.
Trans-Limonenoxid konnte in der Rebsorte Blaufränkisch an beiden Standorten ausschließlich in gebundener Form nachgewiesen werden. Während in Lutzmannsburg ein insgesamt moderater, unregelmäßiger Konzentrationsverlauf beobachtet wurde, zeigte sich in Horitschon eine deutlich dynamischere Entwicklung, mit einem klaren Anstieg der Gesamtfraktion bis zur Normalreife und einem Rückgang zur Überreife. Über alle Zeitpunkte hinweg war der gebundene Anteil bei 100 %, was auf eine ausgeprägte Glykosylierungskapazität hinweist. Der Vergleich der Standorte zeigt, dass in Horitschon durchweg höhere Konzentrationen auftraten, insbesondere im Bereich der Normalreife. Die enzymatische Umwandlung von Limonen zu Limonenoxid erfolgt wahrscheinlich über Cytochrom-P450-vermittelte Reaktionen, wobei das trans-Isomer in der Literatur häufiger auftritt (Ilc et al., 2017; Bosman & Lashbrooke, 2023; Schwab & Wüst, 2015). Das vollständige Fehlen des cis-Isomers in beiden Lagen – sowohl in freier als auch in gebundener Form – lässt auf eine nur sehr geringe Aktivität der zugehörigen Synthesewege oder auf Sortenbeschränkungen schließen. In der Literatur wird Limonenoxid überwiegend in aromatischen Sorten beobachtet, während es in neutralen Rebsorten meist unterhalb der Nachweisgrenze bleibt (Kalua & Boss, 2010). Die erste Hypothese eines nicht-linearen Reifeverlaufs wurde für das trans-Isomer erfüllt, insbesondere in Horitschon. Die zweite Hypothese der Dominanz gebundener Monoterpene wurde durch den vollständigen Bindungsstatus bestätigt. Für das cis-Isomer war mangels Nachweisbarkeit keine Hypothesenprüfung möglich.
Neroloxid konnte in der Rebsorte Blaufränkisch an den beiden untersuchten Standorten Horitschon und Lutzmannsburg nur in Einzelfällen in der gesamten Fraktion nachgewiesen werden, während die freie Fraktion durchgehend unterhalb der Nachweisgrenze lag. In Lutzmannsburg war Neroloxid lediglich zu Beginn der Beerenentwicklung messbar, in Horitschon hingegen ausschließlich zur Vorreife. Die Nachweise erfolgten in beiden Fällen ausschließlich für die gebundene Form, was auf eine hohe Glykosylierungskapazität dieses oxidierten Monoterpens schließen lässt. Aufgrund der sehr lückenhaften Nachweisbarkeit lässt sich weder ein zeitlicher Verlauf noch ein systematischer Unterschied zwischen den Standorten erkennen. Die geringe Präsenz von Neroloxid steht im Einklang mit den ebenfalls niedrigen Konzentrationen seines Vorläufers Nerol und lässt auf eine insgesamt geringe Aktivität des zugehörigen oxidativen Stoffwechselwegs schließen. In der Literatur wird Neroloxid fast ausschließlich in aromatischen Sorten wie Muskateller oder Gewürztraminer detektiert, während es in neutralen Sorten kaum vorkommt (Mateo & Jiménez, 2000; Schwab & Wüst, 2015; Reynolds & Wardle, 1989; Yuan & Qian, 2016). Die enzymatische Umwandlung von Nerol zu dessen Oxidationsprodukten setzt eine aktive Beteiligung von Cytochrom-P450-Enzymen voraus, deren Expression ebenfalls stark sortenabhängig ist (Ilc et al., 2017). Die Hypothese, dass gebundene Fraktionen dominieren, wurde im Fall von Neroloxid bestätigt, da die Verbindung nur in glykosylierter Form nachweisbar war. Ein nichtlinearer Verlauf sowie standortspezifische Unterschiede konnten aufgrund der wenigen Datenpunkte hingegen nicht beurteilt werden. Insgesamt bestätigt das Beispiel Neroloxid die Limitierung des Terpenpotenzials in neutralen Rotweinsorten wie Blaufränkisch und zeigt, dass einige oxidierte Monoterpene in diesen Sorten analytisch keine relevante Rolle spielen.
Lavandulol konnte in der Rebsorte Blaufränkisch an keinem der beiden untersuchten Standorte – Lutzmannsburg und Horitschon – zu keinem der Erhebungszeitpunkte in quantifizierbarer Konzentration nachgewiesen werden und ist daher in den Abb. 1 und 2 nicht dargestellt. Konzentrationen dieses Monoterpens waren über den gesamten Reifeverlauf hinweg weder in freier noch in gebundener Form quantifizierbar. Auch an den drei potenziellen Lesezeitpunkten Vorreife, Normalreife und Überreife blieb Lavandulol vollständig unterhalb der Nachweisgrenze. Das völlige Fehlen dieser Verbindung steht im Einklang mit der Charakteristik von neutralen Rebsorten, die allgemein nur sehr geringe Mengen an Monoterpenen synthetisieren (Mateo & Jiménez, 2000). Lavandulol ist ein seltener Monoterpenalkohol, der insbesondere in floralen Pflanzen und in hoch aromatischen Rebsorten wie Muskateller auftritt (Gunata et al., 1985; Schwab & Wüst, 2015). Seine Biosynthese setzt das Vorhandensein spezifischer Terpensynthasen voraus, deren Expression in Blaufränkisch offenbar nicht erfolgt (Ilc et al., 2016). Auch die Umwandlung verwandter Monoterpene wie Linalool oder α-Terpineol zu Lavandulol scheint hier nicht stattzufinden. Da an beiden Standorten identisches Klonmaterial verwendet wurde, erscheinen genetisch-klonale Unterschiede als Ursache unwahrscheinlich. Epigenetische Effekte, die durch standortspezifische Umweltfaktoren induziert werden können, lassen sich jedoch nicht vollständig ausschließen (Carrier et al., 2012; Pecinka et al., 2013). Die Hypothesen der Studie konnten für Lavandulol nicht geprüft werden, da keinerlei quantifizierbare Daten vorlagen. Auch ein Standorteffekt war nicht erkennbar.
α-Terpineol war in der Rebsorte Blaufränkisch an beiden untersuchten Standorten über den gesamten Zeitraum der Beerenentwicklung hinweg in der freien und der gebundenen Fraktion regelmäßig nachweisbar, wobei sich zwischen den beiden Lagen deutliche Unterschiede im Verlauf der freien Fraktion zeigten. Während in Horitschon die freie Fraktion durchgehend quantifizierbar blieb und nur leichte Schwankungen aufwies, war sie in Lutzmannsburg zu zwei Zeitpunkten nicht nachweisbar und zeigte insgesamt ein unregelmäßigeres Muster. Die Gesamtfraktion stieg an beiden Standorten kontinuierlich an und erreichte ihren Höhepunkt zur Überreife, wobei in Horitschon ein etwas konstanterer Verlauf beobachtet wurde. Die gebundene Fraktion dominierte in beiden Lagen, wobei sie in Lutzmannsburg deutlich stärkere Schwankungen aufwies – der Anteil reichte dort von 22 % bis 100 %, während er in Horitschon in einem konstanteren Bereich zwischen 32 % und 57 % lag. Diese Unterschiede könnten unter anderem auf das Rebalter zurückzuführen sein: Ältere Anlagen wie in Horitschon tendieren zu einer konstanteren Aromaausprägung (Riffle et al., 2022). α-Terpineol entsteht meist aus Linalool und wird überwiegend in gebundener Form gespeichert (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991; Gunata et al., 1985; Schwab & Wüst, 2015), was durch die gemessenen Daten bestätigt wurde. Das Isomer γ-Terpineol hingegen war zu keinem Erhebungszeitpunkt nachweisbar, was auf eine fehlende Enzymaktivität oder genetische Nicht-Expression der entsprechenden Synthesewege in Blaufränkisch schließen lässt (Ilc et al., 2016). Es ist daher in den Abb. 1 und 2 nicht dargestellt. Die Hypothese eines nicht-linearen Konzentrationsverlaufs konnte für die freie Fraktion insbesondere in Lutzmannsburg bestätigt werden, während die zweite Hypothese, die Dominanz der gebundenen Form, an beiden Standorten erfüllt wurde. Der Standortvergleich zeigte konstantere Verläufe in Horitschon und größere Variabilität in Lutzmannsburg. Für γ-Terpineol war aufgrund vollständiger Nichtnachweisbarkeit keine Hypothesenprüfung möglich.
Nerol konnte in der Rebsorte Blaufränkisch an beiden untersuchten Standorten regelmäßig in der Gesamtfraktion nachgewiesen werden, wobei in Horitschon durchgehend höhere Konzentrationen gemessen wurden als in Lutzmannsburg. Die freie Fraktion war insgesamt nur vereinzelt bestimmbar: In Lutzmannsburg lediglich zur zweiten und vierten Erhebung, in Horitschon ausschließlich zur Normalreife. In Lutzmannsburg zeigte sich über die gesamte Beerenentwicklung hinweg ein kontinuierlicher Anstieg der Gesamtfraktion, während in Horitschon ein gleichmäßiger Verlauf auf höherem Ausgangsniveau mit leichten Anstiegen zur Vor- und Überreife beobachtet wurde. Die gebundene Fraktion dominierte über alle Zeitpunkte hinweg, insbesondere in Lutzmannsburg, wo teils vollständige Bindung (100 %) festgestellt wurde. In Horitschon blieb der gebundene Anteil zwischen 87 % und 100 %. Nerol tritt als typischer glykosidisch gebundener Monoterpenalkohol auf, dessen Bildung eng mit dem Reifeprozess verknüpft ist und der als Vorläufer für weitere Terpene wie Citronellol fungieren kann (Gunata et al., 1985; Schwab & Wüst, 2015). Der deutlich höhere Gehalt in Horitschon könnte durch das höhere Rebalter sowie durch günstigere kleinklimatische Bedingungen bedingt sein (Marais et al., 2001; Riffle et al., 2022; Reynolds, & Wardle, 1989). Der Standort Lutzmannsburg zeigte hingegen eine größere Variabilität und punktuelle Ausprägung der freien Fraktion. Die erste Hypothese, wonach die Monoterpenkonzentrationen über die Reife nicht-linear verlaufen, wurde durch die diskontinuierliche Präsenz der freien Fraktion sowie den kontinuierlichen Gesamtanstieg weitgehend bestätigt. Die zweite Hypothese der Dominanz gebundener Fraktionen wurde durch alle Daten eindeutig gestützt. Auch ein Standorteffekt lässt sich feststellen, wobei Horitschon nicht nur höhere Gesamtgehalte aufwies, sondern auch konstantere Verläufe zeigte. Insgesamt bekräftigen die Ergebnisse die Relevanz der gebundenen Form von Nerol für das aromatische Potenzial in Blaufränkisch-Weinen und unterstreichen den Einfluss von Standortbedingungen auf die Terpenentwicklung.
Im Zuge der Vinifikation verändern sich Monoterpenprofile durch chemische und physikalische Prozesse. Monoterpene – die in den Beeren überwiegend in geruchloser Form vorliegen (Mateo & Jiménez, 2000; Ilc et al., 2017; Guth, 1997) – können durch enzymatische oder saure Hydrolyse teilweise freigesetzt und damit sensorisch wirksam werden (Michlmayr et al., 2012; Ilc et al., 2017). Die zunehmende Ethanolkonzentration während der Gärung erleichtert zudem die Extraktion, während gleichzeitig flüchtige Anteile durch CO2 reduziert werden können. Trotz meist geringer Konzentrationen in Rotweinen können einzelne Monoterpene mit niedriger Geruchsschwelle – wie etwa Linalool – dennoch einen subtilen Beitrag zum Bukett leisten (Slaghenaufi et al., 2022).
Citronellol konnte in der Rebsorte Blaufränkisch an beiden untersuchten Standorten ausschließlich in der gebundenen Form nachgewiesen werden. Während in Lutzmannsburg über den Reifeverlauf hinweg ein moderater, aber erkennbarer Anstieg der Gesamtfraktion zu beobachten war – mit einem Peak zur Überreife –, blieb Citronellol in Horitschon abgesehen von der ersten Erhebung durchgehend unterhalb der Nachweisgrenze. In allen quantifizierbaren Fällen lag Citronellol vollständig in gebundener Form vor, was die bekannte Speicherform dieses Monoterpens bestätigt (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991). Der unterschiedliche Verlauf zwischen den Standorten könnte auf mikroklimatische Bedingungen zurückzuführen sein: Während in Lutzmannsburg möglicherweise kühlere Temperaturen und spätere Reife eine verzögerte, aber gesteigerte Synthese ermöglichten, könnten in Horitschon äußere Einflüsse – etwa hohe UV-Belastung – eine frühzeitige oder geringere Produktion bewirkt haben (Gil et al., 2012; Liu et al., 2024; Marais et al., 2001; Song et al., 2015; Skinkis et al., 2010; Reynolds & Wardle, 1989). Die erste Hypothese eines nicht-linearen Konzentrationsverlaufs wurde durch den sprunghaften Anstieg in Lutzmannsburg erfüllt. Die zweite Hypothese, wonach Monoterpene überwiegend in gebundener Form vorliegen, wurde durch die ausgeprägte Glykosylierung von Citronellol klar bestätigt. Auch ein Standorteffekt lässt sich erkennen, wobei Lutzmannsburg eine höhere Citronellol-Akkumulation zeigte als Horitschon. Insgesamt bestätigt sich, dass Citronellol in Blaufränkisch nur in geringen Mengen vorkommt, vorrangig in späteren Reifestadien auftritt und vollständig gebunden vorliegt – typisch für nicht-aromatische Rebsorten.
Geraniol konnte in der Rebsorte Blaufränkisch an beiden untersuchten Standorten regelmäßig nachgewiesen werden, sowohl in freier als auch in gebundener Form. In Lutzmannsburg zeigte die freie Fraktion einen unregelmäßigen Verlauf mit Nachweisen zur ersten, vierten und letzten Erhebung, während sie zu den übrigen Zeitpunkten unterhalb der Nachweisgrenze lag. In Horitschon hingegen war die freie Fraktion ab der Vorreife durchgehend bestimmbar und stieg insbesondere bis zur Normalreife kontinuierlich an; in der darauffolgenden Überreife blieb sie auf konstant hohem Niveau. Die Gesamtfraktion entwickelte sich in Lutzmannsburg unregelmäßig, mit einem Minimum zur zweiten Erhebung und einem klaren Anstieg zur Überreife. In Horitschon verlief die Entwicklung der Gesamtfraktion gleichmäßiger, mit einem kontinuierlichen Anstieg über alle Erhebungspunkte hinweg bis zum Höchstwert zur Überreife. Der gebundene Anteil dominierte in beiden Lagen deutlich: In Lutzmannsburg lag er stets über 84 %, in Horitschon sogar über 88 %. Die kontinuierlichere Akkumulation in Horitschon lässt sich möglicherweise auf das höhere Rebalter zurückführen (Riffle et al., 2022). Die Biosynthese von Geraniol erfolgt über den plastidären MEP-Weg und ist licht-, temperatur- und entwicklungsabhängig (Schwab & Wüst, 2015; Song et al., 2015; Zhang et al., 2017). Die Tatsache, dass Geraniol überwiegend in gebundener Form vorliegt, stimmt mit der Literatur überein, wonach UDP-Glykosyltransferasen wie VvGT7 und VvGT14 für die Glykosidbildung verantwortlich sind (Bönisch et al., 2014a; Bönisch et al., 2014b; Bönisch et al., 2015). Die erste Hypothese, die einen nicht-linearen Verlauf der Konzentration über die Reife annimmt, wurde durch die diskontinuierliche freie Fraktion in Lutzmannsburg sowie die stufenweise Anreicherung in Horitschon bestätigt. Auch die zweite Hypothese wurde erfüllt, da der gebundene Anteil zu allen Zeitpunkten dominierte. Die Unterschiede zwischen den Standorten deuten zudem auf einen Standorteffekt hin, der sowohl im Verlauf als auch im absoluten Niveau der Geraniolkonzentrationen sichtbar wurde.
Die Ergebnisse zu den quantifizierten Monoterpenen an den Pinot Noir-Standorten Guntramsdorf (Riede Eichkogel) und Oberwaltersdorf (Riede Kräutergarten) werden in Abb. 3 und Abb. 4 als Heatmaps visualisiert und in Tab. 5 und Tab. 6 im Anhang durch Mittelwerttabellen ergänzt.
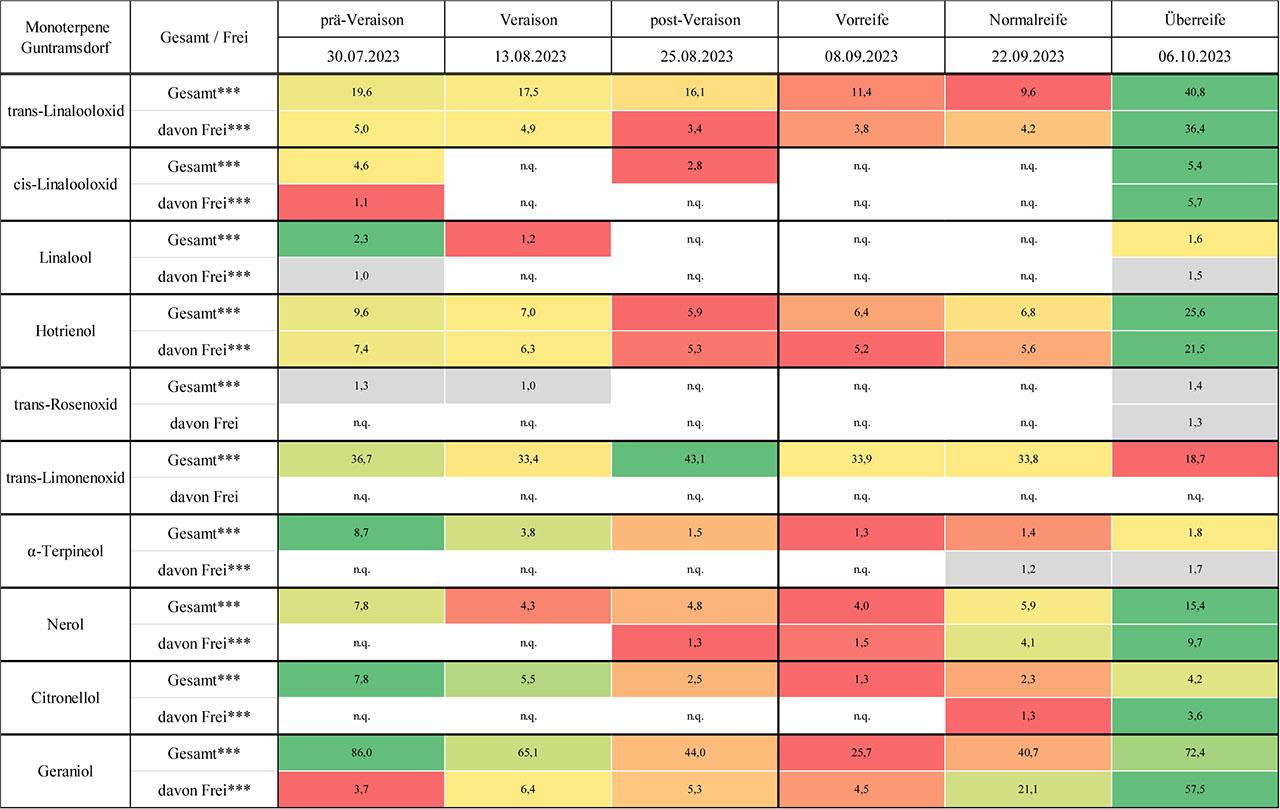
Konzentration [μg/kg Frischgewicht; n =3] und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Pinot Noir-Trauben am Standort Guntramsdorf, Riede Eichkogel („n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Hochsignifikante Unterschiede zwischen den Zeitpunkten sind mit *** und signifikante Unterschiede mit ** gekennzeichnet.)
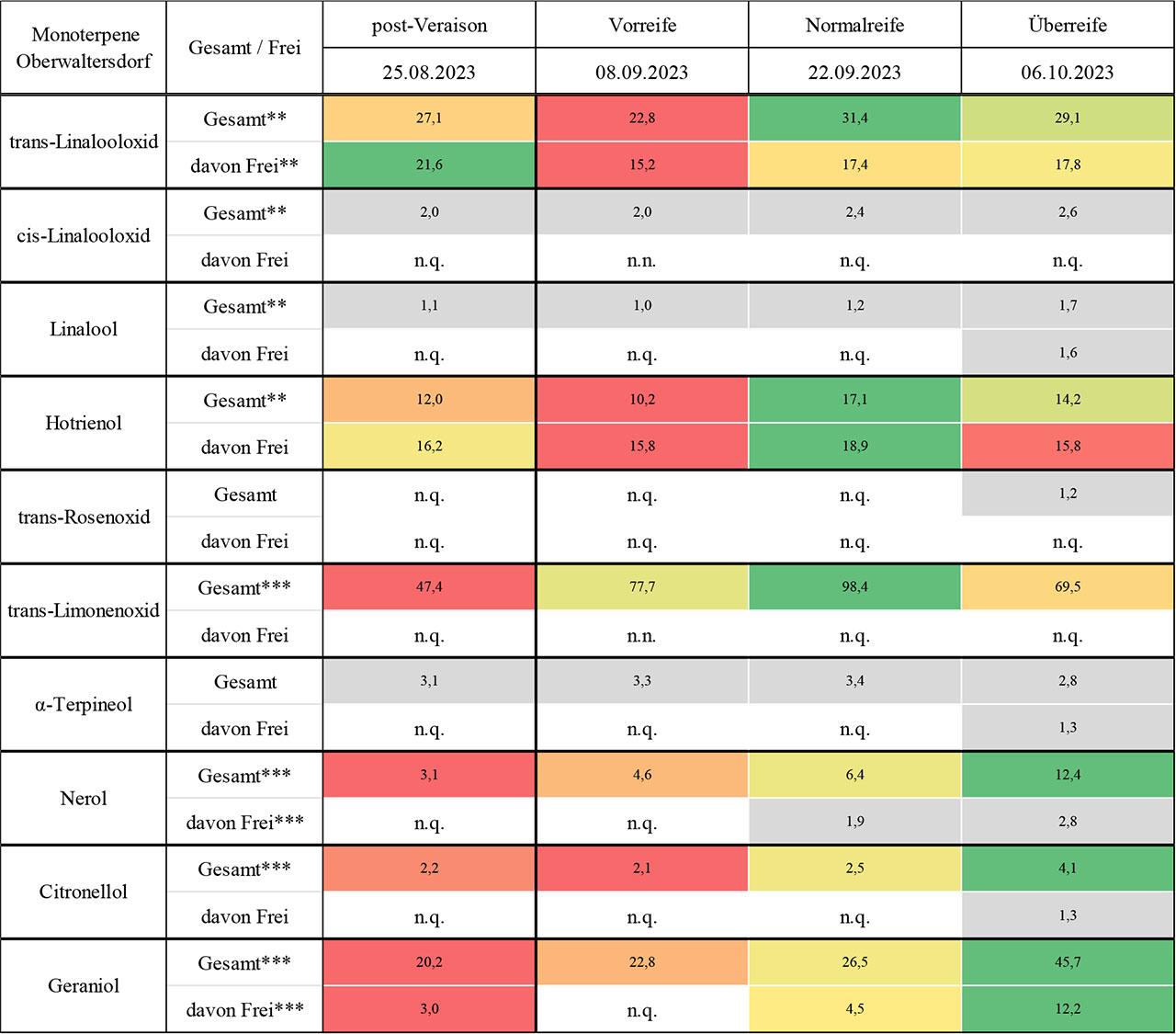
Konzentration [μg/kg Frischgewicht; n =3] und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Pinot Noir-Trauben am Standort Oberwaltersdorf, Riede Kräutergarten („n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Hochsignifikante Unterschiede zwischen den Zeitpunkten sind mit *** und signifikante Unterschiede mit ** gekennzeichnet.)
Konzentration und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Pinot Noir-Trauben am Standort Guntramsdorf, Riede Eichkogel
| [µg/kg Frischgewicht] | Pinot Noir Guntramsdorf, Riede Eichkogel | ||||||
|---|---|---|---|---|---|---|---|
| 23.07.2023 | 13.08.2023 | 25.08.2023 | 08.09.2023 Vorreife | 22.09.2023 Normalreife | 06.10.2023 Überreife | ||
| trans-Linalooloxid | Fr. | 5,0b ± 0,2 | 4,9b ± 0,3 | 3,4a ± 0,4 | 3,8a ± 0,4 | 4,2a ± 0,3 | 36,4c ± 0,8 |
| Ges. | 19,6b ± 2,2 | 17,5b ± 1,5 | 16,1b ± 2,8 | 11,4a ± 1,7 | 9,6a ± 1,2 | 40,8c ± 0,7 | |
| Geb. [%] | 75% | 72% | 79% | 67% | 56% | 11% | |
| cis-Linalooloxid | Fr. | 1,1a ± 0,2 | n.q. | n.q. | n.q. | n.q. | 5,7b ± 0,2 |
| Ges. | 4,6b ± 0,1 | n.q. | 2,8a ± 0,4 | n.q. | n.q. | 5,4c ± 0,2 | |
| Geb. [%] | 76% | - | 100% | - | - | 0% | |
| Linalool | Fr. | 1,0a ± 0,0 | n.q. | n.q. | n.q. | n.q. | 1,5b ± 0,1 |
| Ges. | 2,3c ± 0,0 | 1,2a ± 0,2 | n.q. | n.q. | n.q. | 1,6b ± 0,0 | |
| Geb. [%] | 58% | 100% | - | - | - | 8% | |
| Hotrienol | Fr. | 7,4a ± 0,5 | 6,3a ± 0,1 | 5,3a ± 0,8 | 5,2a ± 0,4 | 5,6a ± 0,1 | 21,5b ± 4,4 |
| Ges. | 9,6b ± 1,6 | 7,0a ± 0,5 | 5,9a ± 0,6 | 6,4a ± 1,0 | 6,8a ± 0,8 | 25,6c ± 3,4 | |
| Geb. [%] | 23% | 9% | 11% | 19% | 18% | 16% | |
| cis-Rosenoxid | Fr. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | - | - | |
| trans-Rosenoxid | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | 1,3 ± 0,0 |
| Ges. | 1,3b ± 0,1 | 1,0a ± 0,1 | n.q. | n.q. | n.q. | 1,4b ± 0,2 | |
| Geb. [%] | 100% | 100% | - | - | - | 5% | |
| trans-Limonenoxid | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | 36,7bc ± 1,1 | 33,4b ± 3,4 | 43,1c ± 5,4 | 33,9bc ± 2,4 | 33,8bc ± 3,9 | 18,7a ± 1,6 | |
| Geb. [%] | 100% | 100% | 100% | 100% | 100% | 100% | |
| cis-Limonenoxid | Fr. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | - | - | |
| Neroloxid | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. | |
| Geb. [%] | - | - | - | - | - | - | |
| Lavandulol | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. | |
| Geb. [%] | - | - | - | - | - | - | |
| α-Terpineol | Fr. | n.q. | n.q. | n.q. | n.q. | 1,2a ± 0,2 | 1,7b ± 0,1 |
| Ges. | 8,7d ± 1,5 | 3,8c ± 0,5 | 1,5ab ± 0,2 | 1,3a ± 0,1 | 1,4ab ± 0,1 | 1,8b ± 0,2 | |
| Geb. [%] | 100% | 100% | 100% | 100% | 19% | 7% | |
| γ-Terpineol | Fr. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.n. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | - | - | |
| Nerol | Fr. | n.q. | n.q. | 1,3ab ± 0,2 | 1,5b ± 0,1 | 4,1c ± 0,6 | 9,7d ± 1,0 |
| Ges. | 7,8c ± 0,1 | 4,3a ± 0,5 | 4,8ab ± 0,4 | 4,0a ± 0,7 | 5,9b ± 0,3 | 15,4d ± 1,2 | |
| Geb. [%] | 100% | 100% | 73% | 64% | 30% | 37% | |
| Citronellol | Fr. | n.q. | n.q. | n.q. | n.q. | 1,3a ± 0,2 | 3,6b ± 0,0 |
| Ges. | 7,8e ± 1,0 | 5,5d ± 0,2 | 2,5b ± 0,1 | 1,3a ± 0,1 | 2,3b ± 0,3 | 4,2c ± 0,3 | |
| Geb. [%] | 100% | 100% | 100% | 100% | 41% | 14% | |
| Geraniol | Fr. | 3,7a ± 0,3 | 6,4b ± 1,0 | 5,3ab ± 1,1 | 4,5ab ± 0,5 | 21,1c ± 3,2 | 57,5d ± 2,4 |
| Ges. | 86,0d ± 5,9 | 65,1c ± 1,2 | 44,0b ± 2,4 | 25,7a ± 1,5 | 40,7b ± 3,2 | 72,4c ± 5,4 | |
| Geb. [%] | 96% | 90% | 88% | 83% | 48% | 21% | |
| Σ Monoterpene | Fr. | 18,2b ± 0,4 | 17,6ab ± | 15,2ab ± 1,5 | 14,9a ± 1,1 | 37,4c ± 4,2 | 138,8d ± 5,4 |
| Ges. | 184,4d ± 9,4 | 138,6c ± | 120,6c ± 9,2 | 83,9a ± 5,7 | 100,6b ± 8,1 | 187,2d ± 10,8 | |
| Geb. [%] | 90% | 87% | 87% | 82% | 63% | 26% | |
Die Zeilen zeigen die quantifizierten Konzentrationen [μg/kg Frischgewicht] der einzelnen Monoterpene (x̄ ± σ), differenziert nach freier, gesamter und gebundener Fraktion. Signifikante Unterschiede zwischen den Zeitpunkten sind durch unterschiedliche Buchstaben gekennzeichnet. „n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Die Nachweisgrenze (LOD) und die Bestimmungsgrenze (LOQ) der analysierten Monoterpene sind in der Studie von Philipp et al. (2024) dokumentiert.
Konzentration und dynamische Entwicklung einzelner Monoterpene während der Traubenreife von Pinot Noir-Trauben am Standort Oberwaltersdorf, Riede Kräutergarten
| [µg/kg Frischgewicht] | Pinot Noir Oberwaltersdorf, Riede Kräutergarten | ||||
|---|---|---|---|---|---|
| 25.08.2023 | 08.09.2023 Vorreife | 22.09.2023 Normalreife | 06.10.2023 Überreife | ||
| trans-Linalooloxid | Fr. | 21,6b ± 1,2 | 15,2a ± 1,4 | 17,4ab ± 2,5 | 17,8ab ± 1,3 |
| Ges. | 27,1ab ± 3,2 | 22,8a ± 3,2 | 31,4b ± 1,0 | 29,1b ± 1,5 | |
| Geb. [%] | 20% | 34% | 45% | 39% | |
| cis-Linalooloxid | Fr. | n.q. | n.n. | n.q. | n.q. |
| Ges. | 2,0ab ± 0,5 | 2,0a ± 0,2 | 2,4ab ± 0,1 | 2,6b ± 0,1 | |
| Geb. [%] | 100% | 100% | 100% | 100% | |
| Linalool | Fr. | n.q. | n.q. | n.q. | 1,6 ± 0,2 |
| Ges. | 1,1a ± 0,2 | 1,0a ± 0,1 | 1,2a ± 0,1 | 1,7b ± 0,1 | |
| Geb. [%] | 100% | 100% | 100% | 4% | |
| Hotrienol | Fr. | 16,2a ± 2,6 | 15,8a ± 1,4 | 18,9a ± 1,5 | 15,8a ± 1,7 |
| Ges. | 12,0ab ± 1,8 | 10,2a ± 1,7 | 17,1c ± 2,1 | 14,2bc ± 1,1 | |
| Geb. [%] | 0% | 0% | 0% | 0% | |
| cis-Rosenoxid | Fr. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| trans-Rosenoxid | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | n.q. | n.q. | 1,2 ± 0,1 | |
| Geb. [%] | - | - | - | 100% | |
| trans-Limonenoxid | Fr. | n.q. | n.n. | n.q. | n.q. |
| Ges. | 47,4a ± 11,3 | 77,7bc ± 5,5 | 98,4c ± 11,4 | 69,5b ± 3,6 | |
| Geb. [%] | 100% | 100% | 100% | 100% | |
| cis-Limonenoxid | Fr. | n.n. | n.n. | n.n. | n.n. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| Neroloxid | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.q. | n.q. | n.q. | n.q. | |
| Geb. [%] | - | - | - | - | |
| Lavandulol | Fr. | n.q. | n.q. | n.q. | n.q. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| α-Terpineol | Fr. | n.q. | n.q. | n.q. | 1,3 ± 0,2 |
| Ges. | 3,1a ± 0,4 | 3,3a ± 0,4 | 3,4a ± 0,5 | 2,8a ± 0,3 | |
| Geb. [%] | 100% | 100% | 100% | 54% | |
| γ-Terpineol | Fr. | n.q. | n.n. | n.n. | n.q. |
| Ges. | n.n. | n.n. | n.n. | n.n. | |
| Geb. [%] | - | - | - | - | |
| Nerol | Fr. | n.q. | n.q. | 1,9a ± 0,3 | 2,8b ± 0,7 |
| Ges. | 3,1a ± 0,5 | 4,6b ± 0,8 | 6,4c ± 0,1 | 12,4d ± 0,4 | |
| Geb. [%] | 100% | 100% | 70% | 78% | |
| Citronellol | Fr. | n.q. | n.q. | n.q. | 1,3 ± 0,2 |
| Ges. | 2,2a ± 0,3 | 2,1a ± 0,1 | 2,5a ± 0,3 | 4,1b ± 0,1 | |
| Geb. [%] | 100% | 100% | 100% | 68% | |
| Geraniol | Fr. | 3,0a ± 0,5 | n.q. | 4,5b ± 0,2 | 12,2c ± 0,5 |
| Ges. | 20,2a ± 2,2 | 22,8a ± 1,8 | 26,5a ± 1,9 | 45,7b ± 1,9 | |
| Geb. [%] | 85% | 100% | 83% | 73% | |
| Σ Monoterpene | Fr. | 40,8b ± 2,3 | 30,9a ± 2,2 | 42,7b ± 1,9 | 52,8c ± 3,4 |
| Ges. | 118,2a ± 19,4 | 146,6a ± 8,7 | 152,1a ± 53,8 | 183,2a ± 3,5 | |
| Geb. [%] | 66% | 79% | 72% | 71% | |
Die Zeilen zeigen die quantifizierten Konzentrationen [μg/kg Frischgewicht] der einzelnen Monoterpene (x̄ ± σ), differenziert nach freier, gesamter und gebundener Fraktion. Signifikante Unterschiede zwischen den Zeitpunkten sind durch unterschiedliche Buchstaben gekennzeichnet. „n.n.“ bezeichnet nicht nachweisbare Werte, während „n.q.“ für nicht quantifizierbare Werte steht. Die Nachweisgrenze (LOD) und die Bestimmungsgrenze (LOQ) der analysierten Monoterpene sind in der Studie von Philipp et al. (2024) dokumentiert.
Die Analyse von trans- und cis-Linalooloxid in Pinot Noir offenbarte deutliche Unterschiede im Reifeverlauf und zwischen den beiden untersuchten Standorten. Für trans-Linalooloxid zeigte sich in Guntramsdorf eine klare Dynamik mit einem auffälligen Konzentrationsanstieg zur Überreife sowie einer ausgeprägten Umverlagerung von der gebundenen zur freien Fraktion. Dieses Muster verweist auf eine verstärkte enzymatische Freisetzung gebundener Vorstufen im späten Reifestadium, wie sie auch in der Literatur für nicht-aromatische Sorten beschrieben wurde (Park et al., 1991; Bönisch et al., 2014b; Mateo & Jiménez, 2000; Darriet et al., 2012). Oberwaltersdorf hingegen zeigte bei diesem Isomer über den gesamten Reifeverlauf ein deutlich konstanteres Profil mit einer durchgehend dominanten freien Fraktion, was auf eine andere Regulation der Terpenfreisetzung hinweisen könnte. Die Hypothesen der Studie konnten somit für trans-Linalooloxid bestätigt werden: Der Verlauf war nicht linear, sondern durch ausgeprägte Veränderungen im Verlauf der Reife gekennzeichnet, insbesondere zur Überreife. Zudem dominierte die gebundene Fraktion zunächst klar und wurde erst im Spätstadium von der freien Form abgelöst, was die zweite Hypothese ebenfalls stützt. Die Unterschiede zwischen den Standorten lassen sich möglicherweise durch die Höhenlage und unterschiedliche Rebzeilenausrichtung erklären. Guntramsdorf liegt deutlich höher, was potenziell kühlere Nachttemperaturen und eine verzögerte Reifung mit sich bringen könnte – Faktoren, die für eine verzögerte Terpenfreisetzung sprechen (Zhang et al., 2017; Friedel et al., 2016). Hinzu kommt die unterschiedliche klonale Ausstattung der Anlagen, die genetisch unterschiedliche Potenziale zur Monoterpenbildung beinhalten kann (Carrier et al., 2012; Martin et al., 2010). Cis-Linalooloxid konnte dagegen nur in sehr geringem Umfang nachgewiesen werden. In Guntramsdorf war die Verbindung intermittierend vorhanden, insbesondere zur Überreife, während sie in Oberwaltersdorf ausschließlich gebunden und konstant auf niedrigem Niveau messbar war. Die geringe Nachweisbarkeit dieses Isomers stimmt mit Literaturberichten überein, wonach cis-Linalooloxid in nicht-aromatischen Rebsorten nur in Spuren vorkommt und eine hohe Labilität aufweist (Ferreira & Lopez, 2019; Gunata et al., 1985; Schwab & Wüst, 2015).
Die Analyse von Linalool in Pinot Noir offenbart eine klare Dominanz der gebundenen Fraktion in beiden Lagen, insbesondere in den frühen und mittleren Reifestadien. Zur Überreife kam es jedoch in beiden Fällen zu einem deutlichen Anstieg der freien Fraktion, was mit einem Rückgang der gebundenen Form einherging – ein Muster, das auf eine späte enzymatische Freisetzung hindeutet, wie sie für Monoterpene in nicht-aromatischen Sorten beschrieben wurde (Park et al., 1991; Bönisch et al., 2014b; Mateo & Jiménez, 2000; Darriet et al., 2012). In Guntramsdorf zeigte sich der zeitliche Verlauf insgesamt unregelmäßiger, mit Lücken in der Gesamtfraktion und einer späten, aber ausgeprägten Zunahme der freien Fraktion zur Überreife. Dagegen verlief die Entwicklung in Oberwaltersdorf gleichmäßiger: Die Gesamtfraktion nahm über den Reifezeitraum kontinuierlich zu, während die freie Fraktion bis zur Überreife weitgehend fehlte. Diese Unterschiede lassen sich möglicherweise auf standortspezifische Faktoren zurückführen: Die höhere Seehöhe und Nord–Süd-Ausrichtung in Guntramsdorf könnten ein kühleres und tendenziell schattigeres Mikroklima bedingen, das die Reifung verzögert oder dazu führen, dass die Freisetzung gebundener Monoterpene später einsetzt (Reynolds & Wardle, 1989; Reynolds et al., 1996). In Oberwaltersdorf, mit flacherer Lage und Ost–West-Ausrichtung, könnten gleichmäßigere Lichtverhältnisse und stabilere Temperaturen zur kontinuierlicheren Akkumulation beigetragen haben. Diese Ergebnisse stehen im Einklang mit Erkenntnissen, wonach Linalool – insbesondere in Pinot Noir – spät akkumuliert und stark durch Umweltfaktoren wie Sonnenexposition beeinflusst wird (Sasaki et al., 2016; Song et al., 2015; Gil et al., 2012; Zhang et al., 2017). Die Hypothese eines nicht-linearen Verlaufs kann durch den Peak zur Überreife in Guntramsdorf klar bestätigt werden. Auch die zweite Hypothese – Dominanz der gebundenen Fraktion – findet in beiden Lagen Unterstützung, wobei zur Überreife eine Umkehr des Fraktionsverhältnisses erfolgt. Diese Umwandlung gebundener Vorläufer in flüchtige Formen ist auch als Reifeindikator bekannt (Bönisch et al., 2014b; Yuan & Qian, 2016).
Die Analyse von Hotrienol in Pinot Noir zeigt ein spannendes und atypisches Muster im Vergleich zu anderen Monoterpenen: An beiden Standorten dominiert die freie Fraktion, wobei vor allem in Guntramsdorf zur Überreife ein markanter Konzentrationsanstieg sowohl in der freien als auch in der Gesamtfraktion verzeichnet wurde. Dieses sprunghafte Verhalten lässt auf eine reifestadiumsabhängige Freisetzung aus gebundenen Vorstufen schließen, möglicherweise durch verstärkte β-Glucosidase-Aktivität. In Oberwaltersdorf hingegen zeigte sich ein gleichmäßiger Verlauf ohne erkennbare Akkumulationsspitzen, und es konnte zu keinem Zeitpunkt eine gebundene Fraktion detektiert werden. Dies könnte auf eine geringe Glykosylierungskapazität oder eine frühzeitige vollständige Freisetzung hinweisen. Die Hypothese eines nicht-linearen Verlaufs wurde in Guntramsdorf durch den klaren Peak zur Überreife bestätigt, während in Oberwaltersdorf eine eher gleichmäßige Entwicklung vorlag. Die zweite Hypothese, wonach Monoterpene in der gebundenen Form dominieren, konnte für Hotrienol nicht bestätigt werden – im Gegenteil: die freie Fraktion war durchgehend die prägende Komponente, was auf einen besonderen Synthese- oder Speicherweg dieses Linalool-Derivats in Pinot Noir hinweist. Die Unterschiede zwischen den Standorten lassen sich möglicherweise durch topographische und mikroklimatische Faktoren erklären. Der Standort Guntramsdorf, mit seiner höheren Seehöhe, Nord–Süd-Ausrichtung und potenziell kühleren Nachttemperaturen könnte eine spätere, aber intensivere Enzymaktivität fördern, was zu einer konzentrierten Freisetzung gegen Ende der Reife führt (Reynolds & Wardle, 1989). In Oberwaltersdorf scheint die Ost–West-Ausrichtung zu einer konstanteren Terpenentwicklung beizutragen, unterstützt durch die in der Literatur beschriebene Wirkung gleichmäßiger Lichtverhältnisse auf die Terpenbiosynthese (Zhang et al., 2017; Gil et al., 2012; Song et al., 2015). Hotrienol als flüchtiger Monoterpenalkohol mit floralen Noten wird in Pinot Noir trotz insgesamt geringer Mengen offensichtlich spät aktiviert, ein Vorgang, der in der Literatur für nicht-aromatische Sorten ebenfalls beschrieben ist (Mateo & Jiménez, 2000; Yuan & Qian, 2016). Seine Präsenz in freier Form – entgegen der allgemeinen Erwartung – macht ihn zu einem potenziell bedeutenden Aromaträger in der späten Reifephase.
Die beiden Isomere cis- und trans-Rosenoxid zeigten in Pinot Noir insgesamt eine äußerst geringe Nachweisbarkeit. Während cis-Rosenoxid an keinem der beiden Standorte nachgewiesen werden konnte und daher in den Abb. 3 und 4 nicht dargestellt ist, wurde trans-Rosenoxid nur in wenigen Einzelfällen bestimmt – in Guntramsdorf vor allem punktuell über den Reifeverlauf verteilt, in Oberwaltersdorf ausschließlich zur Überreife. Eine systematische Betrachtung der Reifedynamik war damit kaum möglich. Die Hypothesen zur nicht-linearen Entwicklung und zur Dominanz der gebundenen Fraktion konnten im Fall von Rosenoxid nicht belastbar geprüft werden. Zwar lag die gebundene Form in den frühen Stadien am Standort Guntramsdorf in einzelnen Fällen vor, doch fehlte ein konsistenter Verlauf. Zur Überreife trat in Guntramsdorf erstmals die freie Fraktion auf, während in Oberwaltersdorf ausschließlich die gebundene Form bestimmbar war – ein Hinweis auf potenzielle Unterschiede in den Aktivitätsprofilen der Glykosidasen oder in der Verfügbarkeit der Vorläuferstoffe wie Geraniol und Citronellol. Die äußerst geringe Nachweisbarkeit steht im Einklang mit der Literatur, die Rosenoxid primär mit stark aromatischen Sorten wie Muskateller oder Gewürztraminer in Verbindung bringt, während neutrale Sorten wie Pinot Noir nur Spuren produzieren (Gunata et al., 1985; Mateo & Jiménez, 2000; Schwab & Wüst, 2015; Ferreira & Lopez, 2019). Hinzu kommt, dass Rosenoxide als labil gelten und sowohl enzymatisch als auch nicht-enzymatisch weiterverarbeitet werden können (Ilc et al., 2016). Auch wenn die Unterschiede zwischen den Standorten nur minimal erscheinen, weist der späte Nachweis der freien Form in Guntramsdorf gegenüber der ausschließlich gebundenen Form in Oberwaltersdorf auf potenzielle klonale Unterschiede hin (Carrier et al., 2012; Pecinka et al., 2013).
Die Analyse der Limonenoxid-Isomere in Pinot Noir zeigt ein klares Muster: trans-Limonenoxid konnte in beiden untersuchten Lagen ausschließlich in gebundener Form nachgewiesen werden, während das cis-Isomer gänzlich unterhalb der Nachweisgrenze blieb und daher in den Abb. 3 und 4 nicht dargestellt ist. In Guntramsdorf zeigte die Gesamtfraktion über den Reifeverlauf hinweg nur moderate Schwankungen und einen leichten Rückgang zur Überreife, wohingegen Oberwaltersdorf eine ausgeprägtere Reifekurve mit einem deutlichen Anstieg bis zur Normalreife aufwies, bevor auch hier ein Rückgang einsetzte. Ein möglicher Erklärungsansatz für diese Unterschiede sind standortspezifische Einflüsse, auf die in den vorangegangenen Absätzen bereits eingegangen wurde. Auch genetische Unterschiede zwischen den verwendeten Pinot-Noir-Klonen (Dijon 115 vs. FR 52/86) könnten eine Rolle in der Synthese oder Glykosylierung von Limonenoxid spielen (Carrier et al., 2012; Bönisch et al., 2014a; Reynolds & Wardle, 1989). Die vollständige Abwesenheit freier Fraktionen bei trans-Limonenoxid sowie die Nichtexistenz quantifizierbarer Mengen von cis-Limonenoxid entsprechen der Literaturlage. In neutralen Sorten wie Pinot Noir treten Monoterpene generell in sehr geringen Konzentrationen auf und liegen zumeist gebunden vor (Mateo & Jiménez, 2000; Fang & Qian, 2005). Die Hypothese der Dominanz gebundener Monoterpene wird durch die Ergebnisse eindeutig gestützt, während ein nicht-linearer Verlauf nur in Oberwaltersdorf erkennbar war. Insgesamt unterstreichen diese Ergebnisse die geringe aromatische Relevanz von Limonenoxid in Pinot Noir, verweisen jedoch auf subtile standortspezifische Unterschiede in der Akkumulation gebundener Vorläufer.
Für Neroloxid konnte in der Rebsorte Pinot Noir an keinem der beiden untersuchten Standorte eine quantifizierbare Konzentration festgestellt werden. Weder in Guntramsdorf noch in Oberwaltersdorf war diese Verbindung in freier oder gebundener Form nachweisbar. Es ist daher in den Abb. 3 und 4 nicht dargestellt. Diese vollständige Abwesenheit verhinderte sowohl die Darstellung eines Reifeverlaufs als auch eine Bewertung der Fraktionsverhältnisse oder eine standortbezogene Differenzierung. Die Hypothesen der Studie – ein nicht-linearer Verlauf sowie die Dominanz der gebundenen Fraktion – konnten für dieses Monoterpen nicht überprüft werden. Neroloxid ist ein oxidiertes Derivat von Nerol, dessen Bildung auf enzymatische Umwandlung zurückgeht und das selbst in aromatischen Sorten nur in geringen Mengen nachgewiesen wird (Gunata et al., 1985; Luan et al., 2005). Entsprechend ist es nicht überraschend, dass Neroloxid in Pinot Noir unterhalb der Nachweisgrenze blieb. Die vollständige Abwesenheit könnte auch durch ungünstige klimatische Bedingungen am Standort bedingt sein. Technisch denkbar ist ferner, dass die Verbindung während der Probenverarbeitung abgebaut wurde – was insbesondere für labilere, oxidierte Monoterpenstrukturen wie Neroloxid plausibel erscheint (Skouroumounis & Sefton, 2000).
Lavandulol konnte in der Rebsorte Pinot Noir an keinem der beiden untersuchten Standorte nachgewiesen werden und ist daher in den Abb. 3 und 4 nicht dargestellt. Diese vollständige Abwesenheit sowohl in der freien als auch in der gesamten Fraktion verhinderte eine Darstellung des zeitlichen Verlaufs, eine Bewertung der Fraktionsverteilung sowie jegliche Differenzierung zwischen den Standorten. Lavandulol ist ein Hydroxylierungsprodukt von Linalool und seine Bildung ist stark von spezifischen Cytochrom-P450-Enzymen abhängig, deren Expression in Pinot Noir schwach ausgeprägt ist (Ilc et al., 2017; Schwab & Wüst, 2015; Bönisch et al., 2015). Darüber hinaus könnte die geringe Stabilität dieser Verbindung – etwa durch enzymatischen Abbau oder Verflüchtigung – zur analytischen Nichtdetektion beigetragen haben (Skouroumounis & Sefton, 2000). Zudem ist Lavandulol auch in aromatischen Sorten wie Muskateller oder Gewürztraminer nur in Spuren nachweisbar und hat in diesen Fällen kaum sensorische Relevanz (Gunata et al., 1985; Matarese et al., 2014).
α-Terpineol konnte in der Rebsorte Pinot Noir an beiden Standorten regelmäßig in der gesamten Fraktion nachgewiesen werden, wobei sich vor allem in Guntramsdorf ein dynamischer Reifeverlauf mit deutlichem Minimum in der mittleren Phase und einem Anstieg zur Überreife zeigte. In Oberwaltersdorf verlief die Entwicklung gleichmäßiger, mit geringeren Schwankungen in der Gesamtfraktion. Die freie Fraktion war in beiden Lagen erst zur Überreife messbar, was für eine späte Freisetzung durch enzymatische Umwandlung spricht. Über weite Teile des Reifeverlaufs dominierte die gebundene Fraktion klar, insbesondere in den frühen und mittleren Phasen, was mit bisherigen Erkenntnissen zur glykosidischen Speicherung von Monoterpenen übereinstimmt (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991). Der beobachtete Anstieg freier Fraktionen zur Überreife passt zur Literatur, die für Pinot Noir einen späten Anstieg freier Monoterpenalkohole beschreibt (Yuan & Qian, 2016). α-Terpineol wird aus Linalool gebildet und ist in zahlreichen Rebsorten, auch in nicht-aromatischen, nachweisbar – meist jedoch in gebundener Form (Ilc et al., 2016; Schwab & Wüst, 2015). γ-Terpineol hingegen war in keiner Probe nachweisbar und ist daher in den Abb. 3 und 4 nicht dargestellt – ein Befund, der mit der Literatur übereinstimmt, in der diese Verbindung in neutralen Sorten häufig als nicht detektiert erwähnt wurde (Mateo & Jiménez, 2000). Ein möglicher Erklärungsansatz für diese Unterschiede sind standortspezifische Einflüsse, auf die in den vorangegangenen Abschnitten bereits eingegangen wurde. Die Hypothesen dieser Untersuchung – ein nicht-linearer Reifeverlauf und die Dominanz der gebundenen Fraktion – lassen sich für α-Terpineol bestätigen, während γ-Terpineol in Pinot Noir weder analytisch noch aromatisch eine Rolle zu spielen scheint.
Nerol konnte in der Rebsorte Pinot Noir an beiden Standorten regelmäßig in der gesamten Fraktion nachgewiesen werden, wobei sich insbesondere in der freien Fraktion deutliche Unterschiede zwischen den Lagen ergaben. Während in Guntramsdorf ein unregelmäßiger Verlauf mit einem frühen Maximum, einem Rückgang in der mittleren Reifephase und einem Anstieg zur Überreife festgestellt wurde, zeigte sich in Oberwaltersdorf ein kontinuierlicher, gleichmäßiger Anstieg über den gesamten Reifeverlauf hinweg. In beiden Lagen blieb die freie Fraktion in den frühen Phasen unterhalb der Quantifizierungsgrenze und wurde erst ab der mittleren bis späten Reife messbar, was auf eine späte Freisetzung aus der gebundenen Form hindeutet. Die Dominanz der gebundenen Fraktion zu Beginn der Beerenentwicklung entspricht der typischen Speicherung von Monoterpenalkoholen in Form von Glykosiden, wie sie insbesondere für Nerol beschrieben ist (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991). Im weiteren Verlauf der Reife kam es in Guntramsdorf zu einer zunehmenden Freisetzung, sodass zur Überreife ein starker Überhang der freien Fraktion entstand. Oberwaltersdorf hingegen wies auch zur Überreife noch eine klar dominierende gebundene Fraktion auf, was auf einen konservativeren Verlauf der Umwandlung hinweist. Ein möglicher Erklärungsansatz für diese Unterschiede sind standortspezifische Einflüsse, auf die in den vorangegangenen Abschnitten bereits eingegangen wurde. Die Hypothesen der Studie lassen sich für Nerol weitgehend bestätigen: Der Verlauf ist nicht linear, sondern zeigt insbesondere in Guntramsdorf eine deutliche Dynamik. Auch die Dominanz der gebundenen Fraktion über weite Strecken der Beerenentwicklung hinweg konnte an beiden Standorten nachgewiesen werden. Literaturquellen berichten übereinstimmend von einer erhöhten Akkumulation und möglichen Freisetzung von Nerol in späten Reifestadien (Yuan & Qian, 2016; Salinas et al., 2004), was sich in den hier präsentierten Daten widerspiegelt.
Citronellol konnte in der Rebsorte Pinot Noir an beiden Standorten über die gesamte Reifeent-wicklung hinweg in der Gesamtfraktion regelmäßig nachgewiesen werden. Die freie Fraktion blieb dagegen in beiden Lagen über weite Teile des Beobachtungszeitraums unterhalb der Nachweisgrenze und wurde erst zur Überreife quantifizierbar. In Guntramsdorf zeichnete sich die Entwicklung durch ein ausgeprägtes Absinken der Gesamtfraktion zur mittleren Reife und einen anschließenden Anstieg zur Überreife aus, während in Oberwaltersdorf ein gleichmäßigerer Anstieg über alle Erhebungszeitpunkte hinweg beobachtet wurde. Ein möglicher Erklärungsansatz für diese Unterschiede sind standortspezifische Einflüsse, auf die in den vorangegangenen Abschnitten bereits eingegangen wurde. Die Fraktionsverteilung war klar durch die Dominanz der gebundenen Form geprägt, besonders in den frühen und mittleren Reifestadien. Erst zur Überreife kam es zu einer merklichen Freisetzung, was vermutlich auf enzymatische Aktivitäten – insbesondere von Glykosidasen – zurückzuführen ist (Reynolds & Wardle, 1989; Reynolds et al., 1996). Dieses Verhalten deckt sich mit der Literatur, wonach Monoterpene wie Citronellol bevorzugt in glykosylierter Form gespeichert und erst spät freigesetzt werden (Bönisch et al., 2014a; Bönisch et al., 2014b; Park et al., 1991). Citronellol entsteht typischerweise durch Reduktion von Geraniol und ist in aromatischen Sorten wie Muskateller ein relevanter Aromastoff, während er in neutralen Sorten wie Pinot Noir meist unterhalb sensorischer Schwellen liegt (Mateo & Jiménez, 2000; Gunata et al., 1985). Die Hypothesen der Studie lassen sich für Citronellol bestätigen: Die Konzentrationen zeigen einen nicht-linearen Verlauf mit deutlichem Anstieg zur Überreife, und die gebundene Form dominierte über weite Strecken der Reife. Auch wenn die sensorische Bedeutung in Pinot Noir begrenzt ist, dokumentieren die Ergebnisse die potenzielle Rolle von Citronellol als Reifeindikator und als Bestandteil des Glykosidpools, aus dem während der Weinbereitung aromaaktive Verbindungen freigesetzt werden können.
Geraniol, als einer der zentralen Monoterpenalkohole mit floralen und zitrusartigen Duftnoten, wurde in Pinot Noir an beiden Standorten regelmäßig sowohl in der freien als auch in der gesamten Fraktion nachgewiesen. Dabei zeigten sich deutliche Unterschiede im zeitlichen Verlauf und in der Fraktionsverteilung zwischen den Lagen. In Guntramsdorf war die freie Fraktion bereits zu Beginn der Beerenentwicklung messbar und nahm im weiteren Verlauf kontinuierlich zu, mit einem ausgeprägten Anstieg zur Überreife. Die Gesamtfraktion hingegen verlief dort zunächst rückläufig, bevor sie in der Spätphase erneut stark anstieg. In Oberwaltersdorf zeigte sich ein konstanterer Verlauf mit einem kontinuierlichen Anstieg der Gesamtfraktion und einer freien Fraktion, die erst ab der Normalreife messbar war und ebenfalls zur Überreife zunahm. Die gebundene Form war in beiden Lagen in den frühen Reifestadien dominierend und wurde im weiteren Verlauf zunehmend durch die freie Fraktion abgelöst – ein Verhalten, das bereits in früheren Studien für Geraniol beschrieben wurde (Gunata et al., 1985; Park et al., 1991). Diese Umverlagerung lässt sich auf eine zunehmende enzymatische Freisetzung glykosidisch gebundener Monoterpene in der späten Reife zurückführen, möglicherweise begünstigt durch Temperatur- und Lichtreize (Zhang et al., 2017; Gambetta et al., 2014). Geraniol entsteht biosynthetisch über den plastidären DOXP/MEP-Weg und ist stark licht- und temperaturabhängig exprimiert (Luan & Wüst, 2002; Dudareva et al., 2013). In neutralen Rebsorten wie Pinot Noir liegt es überwiegend in gebundener Form vor und trägt weniger direkt zur primären Aromawahrnehmung bei (Mateo & Jiménez, 2000; Schwab & Wüst, 2015). Ein möglicher Erklärungsansatz für diese Unterschiede sind standortspezifische Einflüsse, auf die in den vorangegangenen Abschnitten bereits eingegangen wurde. Die Hypothesen der Studie lassen sich für Geraniol bestätigen: Die Entwicklung verläuft nicht linear, sondern zeigt besonders in Guntramsdorf dynamische Veränderungen. Auch die Dominanz der gebundenen Form in der frühen Reife sowie deren Rückgang zur Überreife ist klar belegt. Diese Ergebnisse stehen im Einklang mit aktuellen Studien zur Geraniolbiosynthese und -freisetzung, die eine verstärkte Aktivität der beteiligten Enzyme in der Spätphase der Reifung vermuten lassen (Ilc et al., 2016; Salinas et al., 2004; Yuan & Qian, 2016).
Die vorliegenden Ergebnisse belegen, dass Monoterpene auch in neutralen Rotweinsorten wie Blaufränkisch und Pinot Noir regelmäßig auftreten und differenzierte Reifeverläufe sowie standortspezifische Ausprägungen zeigen. Insgesamt dominierten gebundene Fraktionen, insbesondere in den frühen und mittleren Stadien der Beerenentwicklung, während zur Überreife hin vermehrt freie Monoterpene messbar wurden. Unter den analysierten Verbindungen erwiesen sich insbesondere Linalool, α-Terpineol und Geraniol als konstant nachweisbar, wohingegen andere Substanzen, wie Neroloxid oder Lavandulol, durchgehend unterhalb der Nachweisgrenze lagen. Die Dynamik der Monoterpenentwicklung war stärker ausgeprägt an Standorten mit Nord-Süd-Ausrichtung, was auf einen Einfluss von Mikroklima und Rebzeilengeometrie schließen lässt. Zudem bestätigen die Ergebnisse die Hypothesen eines überwiegend gebundenen Vorkommens und eines nicht-linearen Konzentrationsverlaufs während der Reife. Die Beobachtungen unterstreichen die Rolle gebundener Monoterpene als aromatisches Potenzial und deuten darauf hin, dass auch in klassischen Rotweinsorten ein differenziertes Verständnis des Terpenstoffwechsels von weinbaulichem und önologischem Interesse sein kann.