
The meat industry provides protein of high nutritional value to human populations worldwide. The meat industry is supported by the production of chicken (62 billion dollars), pork (1.5 billion), lamb (545 million), goats (444 million), and beef (301 million) (Adzitey et al. 2020). The meat industry, as a whole, expects a global growth of 76% by 2050, which is related to population growth (Mahanty et al. 2023). The constant growth in supply and demand is accompanied by higher production volumes and the diversification of products competing for the consumer’s final choice (Omer et al. 2018), which implies challenges to ensure the quality of meat products, mainly in aspects related to food safety certifications and quality certification labels (Aboah and Lees 2020). Food safety involves challenges related to the risks associated with diseases caused by consuming food contaminated by bacteria, parasites and viruses (Buncic 2015).
Bacterial pathogens such as Clostridium perfringens (Noor Mohammadi et al. 2022), Staphylococcus aureus, Salmonella spp., Shigella spp., Vibrio spp., and Escherichia coli (Bantawa et al. 2019; Pungpian et al. 2021) are associated with contamination of meat products and then, outbreaks in human populations (Wang et al. 2020), Other pathogenic bacteria associated with foodborne illnesses include Campylobacter, Yersinia enterocolitica, and Listeria monocytogenes (Chlebicz et al. 2018).
In particular, Shiga toxin E. coli (STEC) is responsible for diarrhea, hemolytic uremic syndrome, and hemorrhagic colitis (Bhoomika et al. 2016; Wang et al. 2017; Sánchez et al. 2021). E. coli (STEC) infections in humans are recurrently associated with the consumption of contaminated and undercooked meat (Beyi et al. 2017; Wang et al. 2017;). Foodborne infections by Shiga toxin-producing E. coli (STEC) account for one million infections per year worldwide; of these cases, 3,890 result in hemolytic uremic syndrome and up to 230 in death (Sánchez et al. 2021). In addition, E. coli is a microorganism listed by the World Health Organization (WHO) as a critical antimicrobial-resistant bacterium (Sonola et al. 2022) and, therefore, plays an important role in the spread and transmission of antimicrobial resistance between production animals and humans (de Been et al. 2014; Okubo et al. 2020). Given the relevance of pathogens such as E. coli in developing bacterial resistance, the food industry has proposed different alternative physical, chemical, and biological methods for bacterial control and their possible eradication in foodstuffs (Islam et al. 2019).
Control measures currently rely on antimicrobial agents, including disinfectants and antibiotics, substances that are not considered environmentally safe and sustainable green methods (Beier et al. 2013; Koga et al. 2015; Ajuwon et al. 2021). In addition, food preservation methods can negatively impacts the organoleptic quality and nutritional status of foods (Garvey 2022). In this scenario, the use of bacteriophages emerges as a biocontrol tool to increase the bactericidal efficiency in foods (Snyder et al. 2016). Bacteriophages or phages infect and kill bacterial populations; in this way, bacteriophages in the natural environment are key to maintaining microbial equilibrium (López-Cuevas et al. 2021). The utility of bacteriophages in the food industry is presented from three approaches: i) the use of phages to treat domesticated livestock to decrease source contamination, ii) treatments to decrease cross-contamination and decontaminate surfaces and fomites in food processing facilities, and, iii) post-production treatments involving direct applications of phages on foods (Sulakvelidze 2013).
The antimicrobial properties of bacteriophages derive from endolysin proteins, responsible for the last step of the lytic cycle of filamentous phages, which act at any point in the food chain, being a complementary and feasible alternative today (Gutiérrez Fernández et al. 2020).
Likewise, the use of bacteriophages presents important advantages in their application, such as high specificity, self-replication-auto limitation, adaptation to bacterial defense mechanisms, low inherent toxicity, relatively inexpensive, environmental resistance, and prolonged shelf life (Sillankorva et al. 2012). Most known phages are of the Caudovirales type, which have tail-like structures that they use to bind to receptors on the host surface and expel their genome from the virion head into the host cell. Infection may be virulent replication (lytic cycle) or passive replication in a lysogenic cycle (Maffei et al. 2021). For E. coli, mainly bacteriophages classified as T phages have been reported, including from T1 to T7 (Maffei et al. 2021; Nikulin et al. 2023). Likewise, the International Committee on Taxonomy of Viruses (ICTV) currently classifies the Caudovirales of Escherichia spp. into 37 genera and 157 species, although new findings in the phylogeny of these viruses continue to be presented (Korf et al. 2019), suggesting that their application in the food and meat industry is still a field of study with ample potential. This work aimed to perform a meta-analysis to evaluate the implementation of bacteriophages as a biocontrol strategy to prevent the contamination of meat products with E. coli.
A bibliographic search was conducted according to PRISMA guidelines (Moher et al. 2009). The first phase consisted of generating search entries (Table I), constructed by selecting the keywords E. coli, bacteriophage, meat product and biocontrol, as well as some synonyms for them (Table I). The established entries were entered in three specialized literature search engines, which were Science Direct (https://www.webof-science.com/wos) and PubMed (https://pubmed.ncbi.nlm.nih.gov).
Search entries established for the identification of bibliographic resources.
| No. | Input |
|---|---|
| 1. | (Escherichia coli OR E. coli) AND (Bacteriophage OR Phage) AND (Meat) AND (Biocontrol) |
| 2. | (Escherichia coli OR E. coli) AND (Bacteriophage OR Phage) AND (Meat product) AND (Control) |
| 3. | (Bacteriophage OR Phage) AND (Escherichia coli OR E. coli) AND (Meat) AND (CFU OR CFU’s) |
| 4. | (Bacteriophage OR Phage) AND (Escherichia coli OR E. coli) AND (Meat) AND (Safety OR Food Safety) |
The search was conducted during between the months of January and July 2024.
Articles included for this work were those that deal with the use of bacteriophages for the control of E. coli in meat matrices or meat products, following the characteristics of i) original articles, ii) published in the last 10 years, iii) English language, iv) articles with quantitative data for colony-forming units (CFU) of E. coli, even if the data were 0 CFU, and v) publications with standard deviation values reported means of E. coli CFU.
The elements identified in the search were filtered through three stages of exclusion according to PRISMA guidelines. During the first stage, articles were excluded that addressed the subject of this work but focused on bacterial species other than E. coli or focused on foods other than meat products; in addition, publications prior to 2014 in a language other than English and documents that corresponded to theses, reviews, meta-analyses, bibliometric analyses, short communications, conference proceedings, books, and book chapters were excluded. In the second stage, bibliographic resources that presented duplicity in the search were excluded. Finally, articles not presenting all the data required for this meta-analysis were excluded. Managing bibliographic resources was always a team effort, where discrepancies for inclusion or exclusion were subjected to peer review.
The bibliographic resources selected for statistical analysis by meta-analysis were loaded into the bibliographic manager Mendeley© in its version 1.19.8 for desktop, from where the articles were scanned for the extraction of data labeled as: Articles were reviewed for the following data extraction, author, year and country as identification data; then, manuscripts were classified by isolate origin, application method, phage application, phage taxonomy, E. coli pathotype, meat origin and meat product type as explanatory variables. Finaly the means ± standard deviation of CFUs reported were captured as the dependent variable for the meta analysis. The articles used up to this stage were 11. After applying the inclusion criteria, a total of 11 articles were selected to perform the meta-analysis (Fig. 1).
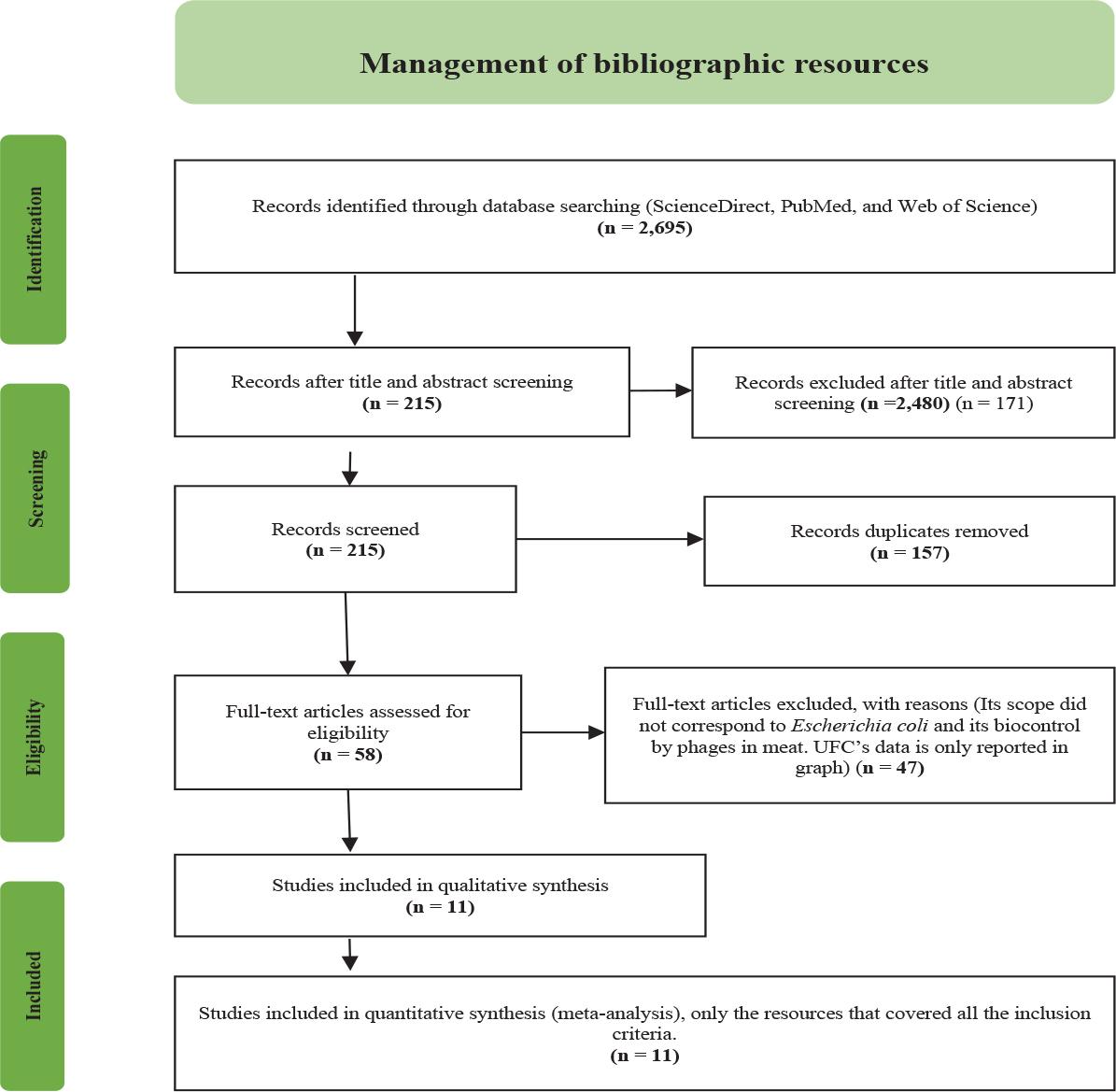
Management of bibliographic resources. Flow diagram describing the management phases for bibliographic resources according to the PRISMA guidelines.
The meta-analysis was developed using R-studio 2024.09.0+375 (Posit Software, PBC., USA), in a computational environment using the “metacont” function of the “meta” package, where the calculation of common and random effects estimates for meta-analysis is performed using continuous outcome data and where inverse variance weighting is used for clustering. For the execution of the “metacont” function, the “SMD” parameter was used to obtain the summary measures of the means of the continuous outcomes as expressed in formula (1) as reported by (Schwarzer 2007):
The approach of the four search entries allowed the identification of 2,695 bibliographic sources initially; this primary compendium was subjected to a scrutiny of information in which 2,480 resources that did not present the inclusion criteria for this meta-analysis were discarded, 157 were eliminated for presenting duplicity (≥ 2) and 47 were excluded because they did not present data in numerical format, including standard deviation values, these articles had the inclusion criteria, however, the data extraction for the meta-analysis did not present viability. Finally, 11 articles were selected for quantitative analysis.
The meta-analysis focused on the grouping of mean differences, where the values of logarithmic decrease of E. coli concentrations were expressed in applying bacterio-aphages as a control strategy in meat products. The 11 bibliographic sources analyzed are shown in Fig. 2, where the overall grouped mean difference was –1.72 Plogt C FU[CIe= –2.12; –1.33] on the logarithmic scale, evaluated through a total of 279 strains of E. coli, which f were challenged with different bacteriophage interventions. The reports showed statistical significance among them (p < 0.05), with heterogeneity by I = 99% among the interventions stand out of those reported by Seo et al. in 2016 with a mean difference of –3.72 log CFU [CI = –4.08; –3.36], followed by that reported by Wang et al. (2020), where a mean difference of –3.55 log CFU [CI = –4.21; –2.89] is observed, which counted 3 and 8 E. coli strains evaluated respectively. The studies presented a geographical distribution in four countries (p < 0.05), which included China (-2.20 log CFU [CI = –2.84; –1.56]), Indonesia (–1.66 log CFU [CI = –2.41; –0.91]), South Korea (–3.72 log CFU [CI = –4.08; –3.36]) and the United States of America (–0.82 log CFU [CI = –1.06; –0.59]).

Forest Plot of moderator analysis for authors. Differences in grouped means are observed in relation to the articles included in this meta-analysis study.
The moderator analysis for the explanatory variable of bacteriophage origin (Origin) showed that the differences in means were significant (p < 0.05) among the different categories (Fig. 3), where up to 202 of 279 observations were made on E. coli strains challenged with commercial bacteriophages; however, this category presented the most negligible effect on the log concentration of E. coli with –1.21 log CFU [CI = –1.59; –0.82]. In contrast, the strains challenged with bacteriophages isolated from excreta presented the most significant effect on the mean difference with a value of –3.55 log CFU [CI = –4.21; –2.89]. The heterogeneity of this category was I = 37%, related to a low number of observations, so the results should be taken with caution.
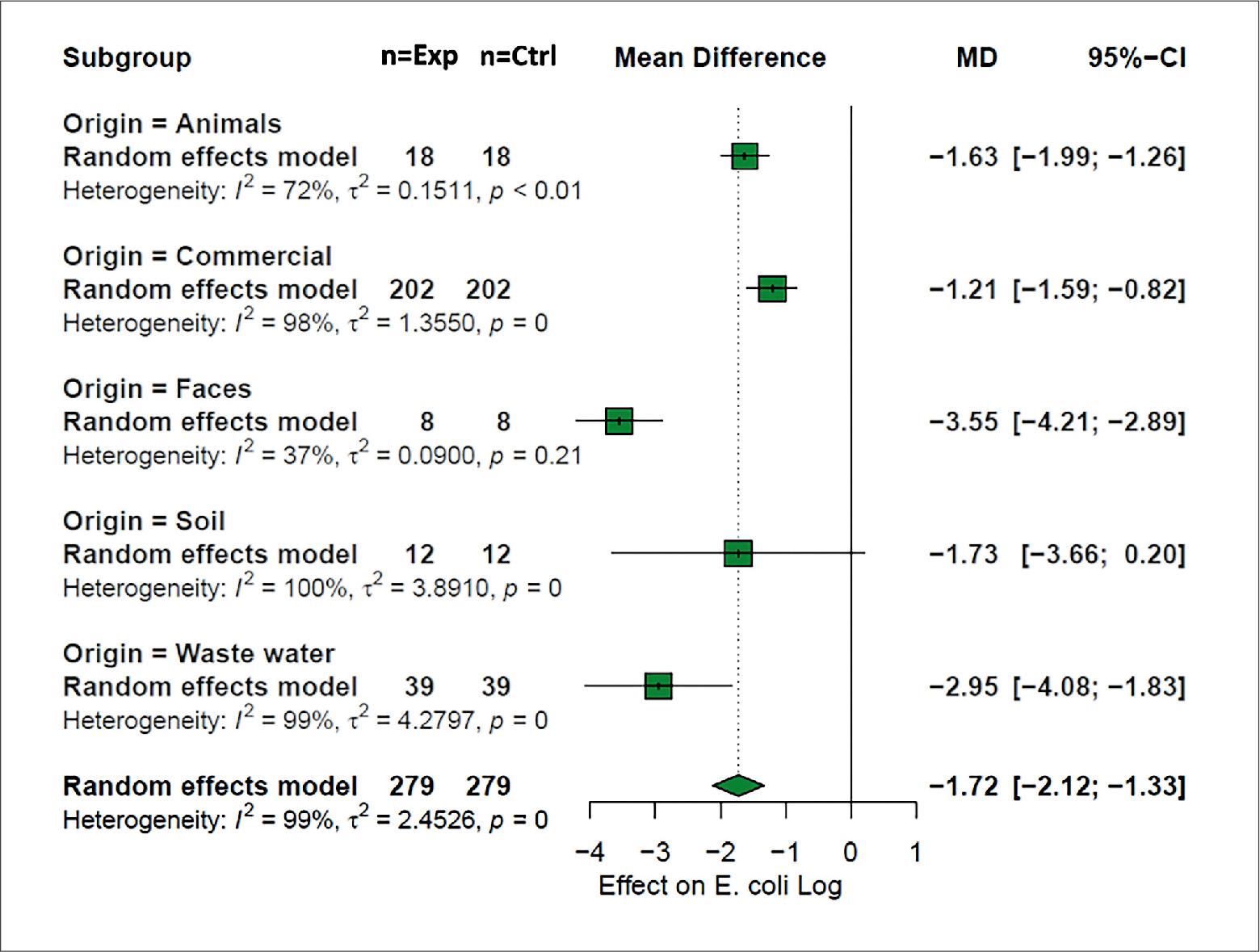
Bacteriophage origin. The difference of grouped means based on the moderator analysis applied to the variable origin of the bacteriophage.
The sort of bacteriophage application was another perspective approached using the moderator analysis (p < 0.05); the two routes of administration were cataloged as individual or in cocktails, which presented grouped mean differences of –2.02 log CFU [CI = –2.51; –1.52] and –0.85 log CFU [CI = –1.08; –0.61] respectively. This analysis was complemented concerning methods of bacteriophage application, Fig. 4 shows differences in grouped means, where the most frequent applications were by direct inoculum and drip inoculum; these categories evidenced a logarithmic decrease on E. coli of –1.13 log CFU [CI = –1.67; –0.58] and –0.96 log CFU [CI = –1.20; –0.73], which places them as those with the least effect. In contrast, the application of bacteriophages using vehicles allowed obtaining a greater effect in the mean difference, where the use of biofilms showed a grouped mean difference of –2.45 log CFU [CI = –3.91; –1.00], followed by the use of encapsulates with a value of –2.12 log CFU [CI = –2.84; –1.40]; however, the number of strains evaluated was low with 26 and 72 respectively, this in comparison with the application of direct inoculums that presented up to 123 observations.

Method of application of bacteriophages on meat products. Forest plot of the moderator analysis for the explanatory variable application method, grouped mean differences are observed.
The taxonomy of the applied bacteriophage was analyzed for mean difference comparison (p < 0.05). A total of (135 observations) were reported as commercial bacteriophages, where the taxonomic classification was not reported. In the other side, 45 observations were categorized as indeterminate cocktail and, 90 were indeterminate phage. These explanatory variables presented grouped mean differences of –0.32 log CFU [CI = –0.57; –0.07] and –2.12 log CFU [CI = –2.79; –1.45] respectively, between these two application categories the use of indeterminate bacteriophage cocktail presented the lowest effect on the log concentration of E. coli. In contrast, when the bacteriophage applied to the meat products belonged to the Myoviridae and Podoviridae families, they showed the greatest effects on the logarithmic concentration of E. coli, in particular the Podoviridae category showed the most significant effect with a value of –3.72 log CFU [CI = –4.08; –3.36], however, the number of strains did not allow the calculation of heterogeneity, so it is convenient to continue monitoring this explanatory variable in future studies (Fig. 5).
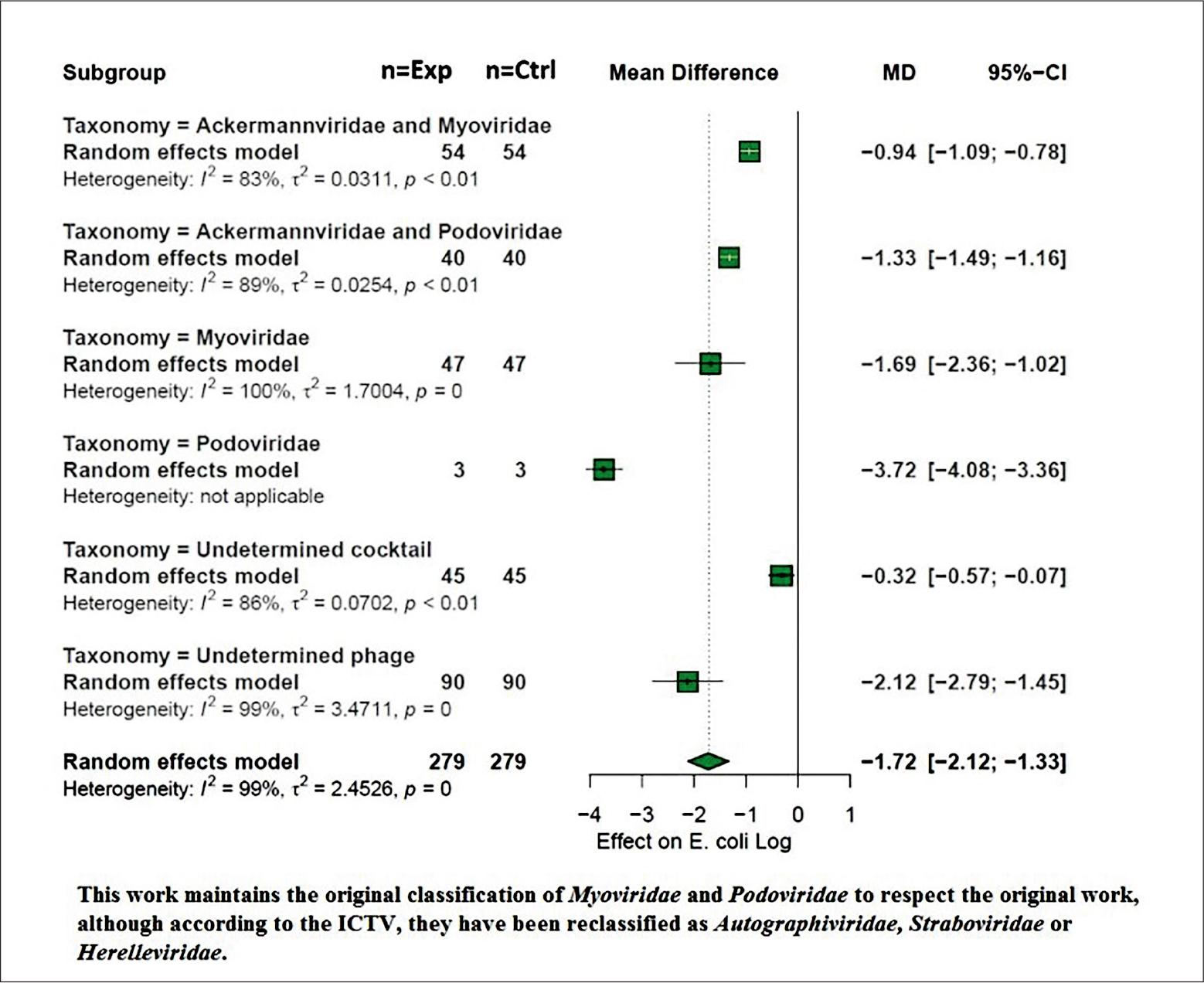
Bacteriophage taxonomy. Moderator analysis of the differences in means grouped according to the taxonomy of the bacteriophage applied in meat products.
Among the different strains of E. coli challenged with bacteriophages on meat matrices, four pathotypes were found (p < 0.05); their moderator analysis is shown in Fig. 6, where Shiga toxin-producing E. coli (STEC) is the most studied pathotype with 147 observations, followed by enterohemorrhagic E. coli with 114 observations. Enterohemorrhagic E. coli with 114 observations, although STEC presented a high number of observations, it showed less effect on the difference of means before the application of bacteriophages –1.11 log CFU [CI = –1.54; –0.68], which in this approach would result in the most resistant pathotype before the challenge with bacteriophages in meat matrices. In addition, enterotoxigenic E. coli was the pathotype with the most significant effect on the grouped mean differences with –2.55 log CFU [CI = –6.69; –1.60], which shows the pathotype with the greatest susceptibility to biocontrol with bacteriophages.
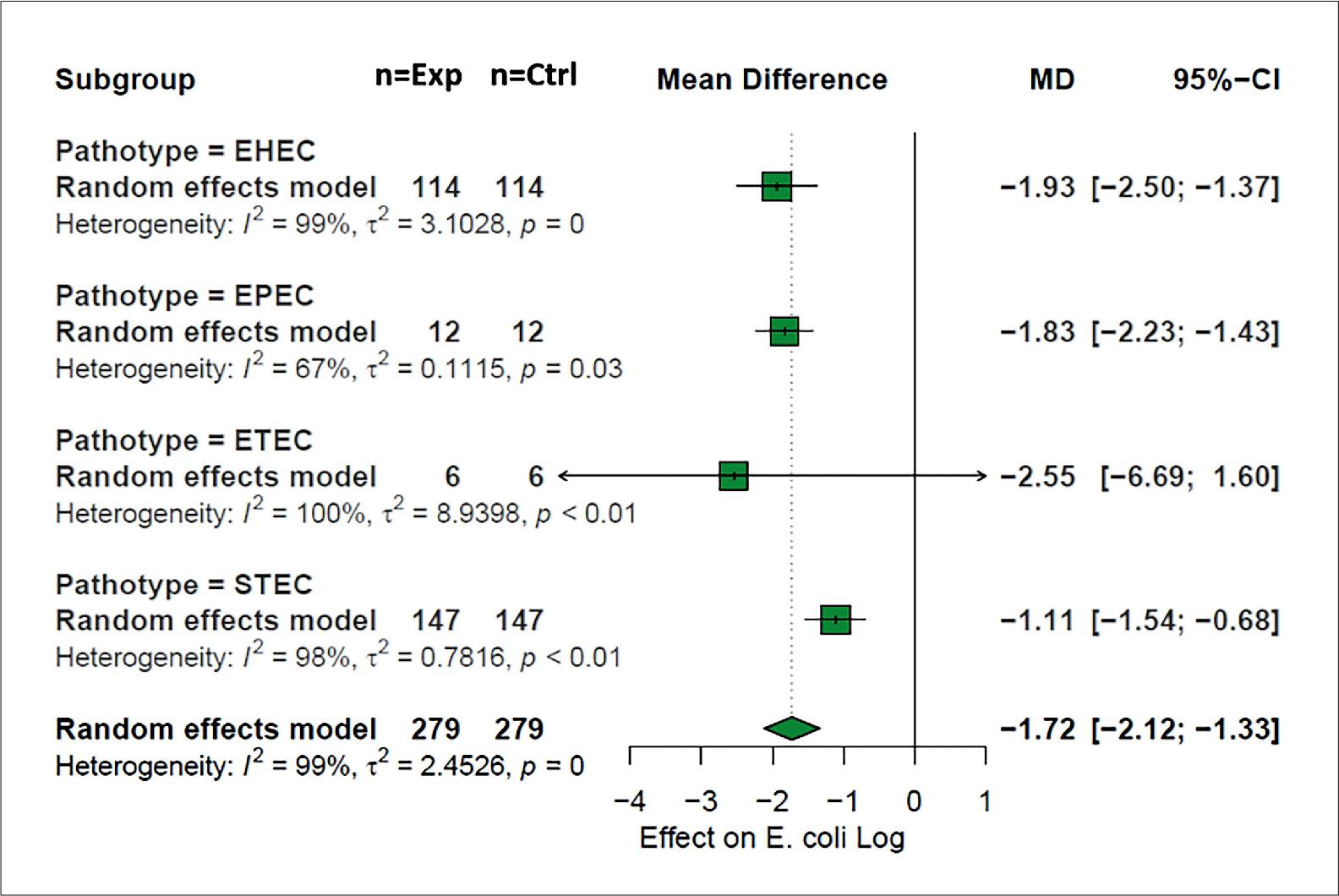
Escherichia coli pathotypes. Analysis of moderators for the explanatory variable pathotype of E. coli,EHEC – enterohemorrhagic E. coli, EPEC – enteropathogenic E. coli,ETEC – enterotoxigenic E. coli and STEC – Shiga toxin-producing E. coli.
Three sources of animal origin were identified, including mainly beef (n = 198), pork (n = 63), and chicken meat (n =18). This comparison presented statistical significance (p < 0.05), with values of –1.88 log CFU [CI = –2.45; –1.32], –1.47 log CFU [CI = –2.10; –0.84] and –1.71 log CFU [CI = –2.96; –0.45] respectively. In particular, when the types of meat products were evaluated, up to seven types of products were identified (Fig. 7). For this explanatory variable, it stood out that the evaluation of the bacteriophage on the concentration of E. coli was found to be limited in dry products such as skin –0.32 log CFU [CI = –0.57; –0.07] as well as in marinated products that presented a logarithmic decrease of –0.34 log CFU [CI = –1.96; –1.29], while primary products such as steaks and carcasses showed a greater effect of the application of bacteriophages with grouped mean differences of –3.55 log CFU [CI = –4.21; –2.89] and –1.92 log CFU [CI = –2.44; –1.41] respectively.
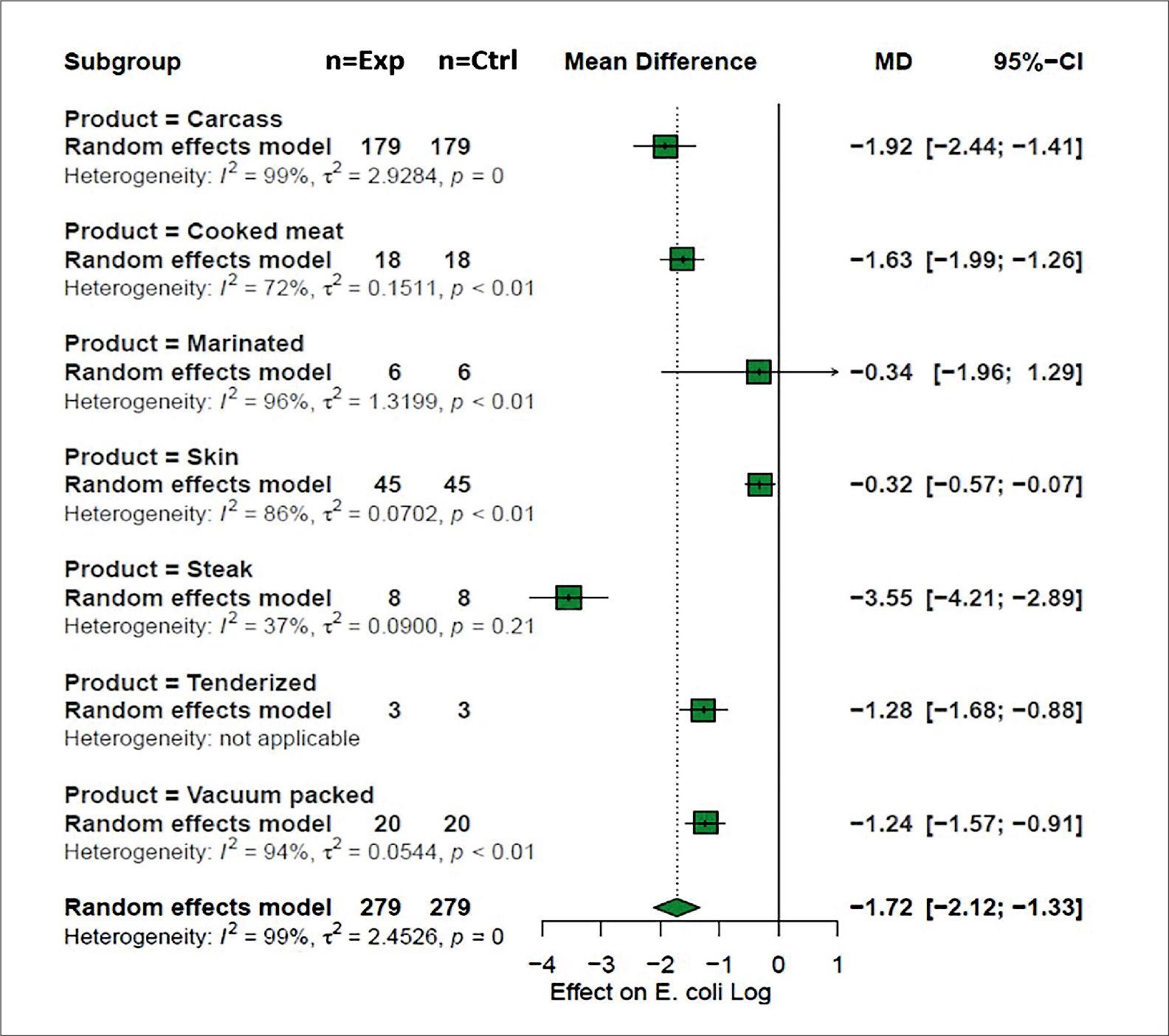
Meat products under analysis. Forest plot of the grouped mean difference of Escherichia coli in meat products that were treated with bacteriophages.
The use of bacteriophages has become a promising biocontrol strategy in the food industry to reduce the risk of gastrointestinal illness caused by pathogens such as E. coli, Salmonella spp., Campylobacter spp., L. monocytogenes, Shigella spp., Vibrio parahaemolyticus, and S. aureus (Ge et al. 2022). In this study, phage treatments targeting E. coli in meat products showed an average log reduction of –1.72 log CFU. Previous research has reported reductions of up to 3 log CFU in vitro tests using bacteriophages against E. coli (Manohar et al. 2019). Furthermore, in foods, such as mixed vegetables, a reduction of E. coli up to log CFU were reported (Kuek et al. 2023); while in meat products under storage conditions, a reduction of 1 log CFUs was reported after 6 days of bacteriophages exposure (Anany et al. 2011).
In this study, commercial phages were the most recurrent strategy for E. coli biocontrol; however, this strategy did not present the best effect in the logarithmic decrease of this pathogen (–1.22 log CFU). Hypothetically, is possible that effect is a consequence of empirical selection of bacteriophages, a situation that denotes the need to describe the underlying molecular mechanisms in the bacteriophage-host interaction (Maffei et al. 2021), recognition of these mechanisms (receptor adsorption and identification, injection of genetic material, phage replication and gene expression, synthesis of virion components, assembly and maturation of new phage, and release of new virions) may allow the design of bacteriophages, an example is the tail fibers and CRISPR-Cas machinery that has demonstrated greater specificity against E. coli in a mouse model, (Gencay et al. 2024). In contrast, bacteriophages isolated from feces showed a greater effect in our study (–3.55 log CFU); this may be related to the fact that feces are an excellent source of bacteriophages, especially those specific for E. coli; in addition, feces are a source of bacteriophages corresponding to three taxonomic groups including somatic coliphages, specific F coliphages and bacteriophages capable of infecting Bacteroides spp. (Toribio-Avedillo et al. 2021).
The form of application of bacteriophages is a key factor in evaluating their efficacy. In our study, we observed that cocktail application showed a lower effect (–1.08 log CFU) than single application of bacteriophages (–2.51 log CFU). This finding is in agreement with that reported by Niu et al. in 2021, who evaluated the efficacy of bacteriophages AKFV33 (Tequintavirus, T5), AHP24 (Rogunavirus, T1), wV7 (Tequatrovirus, T4) and AHP24S (Vequintavirus, rV5), together with 11 combinations of these, in the control of E. coli. The authors noted that the individual phage behavior does not necessarily replicate when combined in a cocktail. This could be because mixing phages with different life cycles causes a mismatch in the concentrations of each phage and favors the establishment of a dominant phage over the others (Niu et al. 2021). Therefore, the food industry is searching other technologies that present synergy with the application of phages (Nikolich and Filippov 2020). In our study, the use of films and encapsulation showed a greater effect of bacteriophage on E. coli (–2.12 to –2.45 log CFU), which can be related to the fact that the use of polymers can provide an environment of protection and controlled release for bacteriophages applied in foods (Weng et al. 2021). In addition, encapsulation with polymers, such as alginate and methacrylate, has the function of bacteriophage protection against pH when oral administration is sought for the control of E. coli (Nikulin et al. 2023). To improve the protection effect, the characteristics of phages should be evaluated to enhance their use in the meat and food industry.
In this work, it is highlighted that up to 135 interventions with phages did not report the corresponding taxonomic classification; however, when taxonomic identification was reported, viruses previously classified as Podoviridae (–3.72 log PFU) and Myoviridae (–1.69 log PFU) presented the best effect on the difference of grouped means. These phages, now reclassified within the class Caudoviricetes, belong to families such as Autographiviridae (formerly Podoviridae) and Straboviridae or Herelleviridae (formerly Myoviridae), which are among the most recurrent taxonomic groups in the development of phage cocktails. They are characterized by an icosahedral head (60 ± 3 nm) and a short, non-contractile tail (35 ± 1.8 nm) in the case of Autographiviridae, while in the case of Straboviridae and related families, they have an icosahedral head (82 ± 4.1 nm) and a contractile tail (199 ± 9.9 nm) (Kornienko et al. 2020). Specifically, the characterization of E. coli bacteriophages has shown that T-type phages are the most recurrent for their control. Among them, the T1 to T7 types stand out. Imke and collaborators in 2019 reported that, among 50 characterized phages, 70% belonged to Straboviridae (formerly Myoviridae), 22% to Drexlerviridae (formerly Siphoviridae), and 8% to Autographiviridae (formerly Podoviridae), including the well-characterized T4 and T5 phages (Korf et al. 2019). In addition, T4 bacteriophages have been reported as recurrent for the control of E. coli because they present non-canonical nucleotide bases, which limit phage transduction and the possibility of horizontal gene transfer, thus reducing undesirable consequences such as bacterial resistance (Nikulin et al. 2023). This is relevant because up to 21 conserved defense systems have been previously reported in E. coli, in a study where 71 strains evaluated showed these mechanisms as strategies against bacteriophages. These defense islands are based on restriction-modification systems and CRIS-PR-Cas, highlighting the continuous coevolution to which bacteria and viruses are subjected (Vassallo et al. 2022).
Another factor to take into account in the relationship between bacteriophage and E. coli is the pathotype that is sought to be subjected to biocontrol with phages; in this item, our meta-analysis showed that EHEC was the pathotype with the highest number of observations (114/279), this denotes the importance of this pathotype as a causal agent of hemorrhagic colitis and hemolytic uremic syndrome (Beyi et al. 2017). It was also observed that STEC was the pathotype on which bacteriophages had the least effect (–1.11 log CFU). This is the relevant situation in view of the toxic-infections caused by this pathotype upon infection in humans worldwide, causing one million cases and up to 100 deaths each year (Mangieri et al. 2021). The low effect of bacteriophages may be related to the fact that E. coli is a microorganism that can present metabolic responses through different cellular mechanisms, which indicates that exposure may lead to phage-mediated selection and may direct the phenotype of this pathogen (Mutalik et al. 2020). It has also been reported that the STEC pathotype can present synergy of pathogenicity in its exposure with bacteriophages that intervene as promoters of cell lysis and in the release of Shiga toxin. They can also act as prophages carrying genes related to pathogenicity in addition to the genes necessary for the phage cycle (Rodríguez-Rubio et al. 2021).
Finally, the effect of phages on E. coli when applied to meat products can present differences according to the animal origin of the meat and the type of meat product evaluated; the present study showed a greater effect on beef and meat products such as steak fillets and carcasses evaluated, the latter also having the most significant number of observations (179/279), in this context meat in general is characterized by being a recurrent vehicle of pathogens related to foodborne diseases (Fegan and Jenson 2018) in particular beef has reported a pH between 5.52 and 6.20 (Triki et al. 2018), in addition, the water activity (A) in beef is high with values ranging from 0.987 to 0.991 (Li et al. 2017), characteristics that favor bacterial growth, so emerging strategies for biocontrol become highly relevant. However, the application of phages cures its problems; among them, the application’s stability stands out (Ge et al. 2022). For example in our meta-analysis, meat matrices that are subjected to preservation processes such as marinades showed a low effect of bacteriophages on the concentration of E. coli (–0.34 log CFU), the marinating processes can generate an increase in lipid oxidation and biochemical and histological changes in the meat (Lebert and Daudin 2014). However, the modification of pH present tendencies towards acidity and a decrease in available water in the meat matrix (Harrison et al. 2006). It has been reported that their effect on bacterial concentration is dependent on several factors, including pH, temperature, exposure time, and the concentration applied (Litt and Jaroni 2017; Niu et al. 2021). Because of this, the meat industry faces other challenges implicit in the use of bacteriophages, among the main ones are the monitoring of phages in the practical applications’ environments and the low acceptance of the presence of a virus (phage) in food by consumers (Ge et al. 2022). The trends described in this work suggest continuing to monitor the specificity between the bacteriophage and E. coli, in search of improving the use of this biocontrol technology in the meat and food industry.
The use of bacteriophages as a biocontrol strategy has been shown to impact the bacterial concentration of E. coli, a pathogen with high public health importance worldwide. The global logarithmic decrease in this study indicates that the intervention can reduce the risk associated with FBD (Foodborne Diseases) in meat consumption; the analysis of moderators showed that some explanatory variables and their management can improve the expected effect on bacterial populations. Among them, isolation of phages from fecal samples, individual application and using polymeric vehicles, as well as application in primary meat products such as fillets and carcasses, may be characteristics that allow better results from the application of phages in meat products, in the search for reducing the risk of infection by E. coli infection. In this sense, the characteristics of the bacteriophage and the bacterium and their interaction mechanisms continue to be indispensable in their analysis to manage the intervention better. For all these reasons, it is highly relevant to continue the specific study of the bacteriophage-host bacterium relationship with E. coli and other pathogens of public health importance in order to improve the use of this emerging technology for bacterial control in the meat production chain, as well as in other foods.