
The genus Streptomyces, first described by Waksman and Henrici, is currently recognized as comprising 901 active species. In addition to Streptomyces, the family includes other members, such as Actinosporangium and Microellobosporia. Actinobacteria, known for their role as producers of active secondary metabolites, have been utilized in producing various natural products (Bundale et al. 2019; Lee et al. 2020a; 2020b), including antibiotics, vitamins, and enzymes, which hold significant value in both agriculture and industry. Their significance has been increasingly recognized in biotechnology, pharmacy, and agriculture in recent years (Sedeek et al. 2023).
Streptomyces is a Gram-positive bacterium that produces spores. Streptomyces are widely distributed in the environment (Olanrewaju and Babalola 2019; Sedeek et al. 2022). Different species of Streptomyces have been found in different soils and marine habitats, as well as in extreme environments such as deserts and volcanic areas (Jia et al. 2015). A large number of bioactive compounds have been previously extracted from Streptomyces. Streptomyces produces secondary metabolites with various pharmacological properties, including antibacterial, antifungal, antitumour, anti-inflammatory, and immunosuppressive effects (Risdian et al. 2019). To obtain these metabolites, culture conditions are critical and Bode et al. (2002) proposed the One Stain Many Compounds (OSMAC) strategy in 2002, which has been playing an essential role since then. OSMAC is a method to fully utilize the ability of microorganisms to synthesize secondary metabolites by changing the nutrient medium, co-culture conditions, and epigenetic modulation, thus obtaining a wider variety of structurally novel and active secondary metabolites (Gao et al. 2024). Since 2015, more than 70 Streptomyces species have been isolated from terrestrial ecosystems and yielded over 170 novel bioactive metabolites. Many of these compounds had antimicrobial activity even against multidrug-resistant pathogens. These secondary metabolites exhibit antimicrobial activity, for example, tirandamycins A and B, originally isolated from Streptomyces tirandis in 1971, are antibiotics that exhibit antimicrobial activity against a wide range of bacteria (Mo et al. 2011). Rejectinomycin has antagonistic activity against Staphylococcus epidermidis (Kim et al. 2022). Streptomyces species have since been recognized as a significant source of various antibiotics, such as streptomycin, tetracycline, chloramphenicol, clindamycin, lincomycin, erythromycin, and kanamycin, among others (Quinn et al. 2020). Examples include krisynomycin (B and C), picolinamycin, nybomycin D, puromycin B-E, ulleungmycin (A and B), streptoone A, and other compounds (Sedeek et al. 2023). In 1970, Reusser et al. discovered tirandamycin, a compound known for its inhibitory effect on ribonucleic acid polymerase. Subsequently, Carlson et al. (2011) reported the homologues tirandamycin C, D, and E, while Yu et al. (2011) also reported tirandamycin F and G in the same year. Collectively, these compounds exhibit enhanced activity.
In this study, we conducted an in-depth exploration of the metabolic potential of Streptomyces gossypiisoli TRM 44567. We identified 35 potential gene clusters through bioinformatics analysis, with particular emphasis on the tirandamycin synthesis gene cluster. Utilizing the OSMAC strategy and R5 medium for fermentation experiments, we successfully isolated and purified tirandamycin A, which exhibits inhibitory effects on Pseudomonas aeruginosa. This study highlights the rich metabolic capacity of S. gossypiisoli TRM 44567 and establishes a comprehensive set of strategies that combine genome mining, mass spectrometry analysis, and optimization of fermentation conditions. These strategies can be applied to the future exploitation of the metabolic potential of other strains, thereby providing valuable insights and methodologies.
Strain TRM 44567 was isolated from continuous cropping cotton fields in Xinjiang, northwest PR China (40°22’N 80°30’E) (Zhang et al. 2021). The sample was isolated on GJ medium by using a serial dilution method, at 37°C for 1 week.
The microbial strains used in this study for antimicrobial activity testing included Staphylococcus aureus (ATCC® 25923™), Escherichia coli (ATCC® 25922™), Klebsiella pneumoniae (ATCC® 10031™), Acinetobacter baumannii (ATCC® 19606™), Candida albicans (ATCC® 64550™), Pseudomonas aeruginosa (ATCC® 27853™), Salmonella enterica subsp. enterica (ATCC® 700623™), Shigella sp. (ATCC® 23354™) and Enterococcus faecalis (ATCC® 29212™) (Honorato et al. 2024).
Extraction and splicing of strain TRM 44567 and the genome were completed in the preliminary stage. The relevant literature can be referred to (Zhang et al. 2021). The accession number of its sequence is JACAOI000000000.
The study on biosynthetic gene clusters responsible for secondary metabolites in the genomes of strain TRM 44567 employed the antiSMASH 6.0-web tool (Blin et al. 2021). Identifying potential products from these gene clusters involved analyzing gene sequences, gene syntenies in established compound BGCs, and the structures domain of PKSs and NRPSs (Komaki et al. 2014).
The “One Strain Multiple Compound” (OSMAC) strategy, which involves altering microbial culture conditions, has proved to be particularly effective in mining many novel secondary metabolites over the past few years (Zhang et al. 2024). In order to optimize the fermentation conditions and to improve the capability of strain TRM 44567 to produce antimicrobial metabolites, 13 solid media (01-12 and R5, as described in Table SI) were tested in 100 ml plates. Cultures were incubated at 30°C for 10 days and then extracted with methanol to investigate the effect of different media on the metabolic potential of strain TRM 44567.
The analysis was conducted using an Agilent 6545 QTOF mass spectrometer coupled with a Waters HSS T3 column (2.1 × 100 mm, 1.8 μm). The mobile phase consisted of water with 0.1% formic acid (Phase A) and acetonitrile (Phase B), delivered at a 0.2 ml/min flow rate. A gradient elution from 10% to 100% was applied over 40 minutes. The samples, dissolved in methanol, were analyzed using an ESI-MS with a linear ion trap mass spectrometer, employing an electrospray ionization source and an injection volume of 10 μL. The mass spectrometer operated in positive ion mode with a scanning range of 100-3,000 m/z, and the gas flow rate was set to 8 l/min.
Strain TRM 44567 was inoculated into 20 l of R5 solid fermentation medium and incubated at 30°C for 10 d. At the end of fermentation, the medium was cut into pieces according to the area of 1 cm × 1 cm. An equal volume of methanol was added to ultrasonic extraction four times. The extracts were combined and filtered through a Büchner funnel to remove impurities, and then, they were rotary evaporated and dried to obtain the crude product.
The obtained crude extract was dissolved in 1 l of ultrapure water and filtered to remove insoluble impurities. Then, the water-soluble crude extract sample was loaded on a column filled with macroporous resin (AB-8). The column was rinsed with about 5 times the volume of the column of purified water to remove the significant polar substances until the effluent was colorless. The column was slowly eluted with 30%, 50%, 70%, and 100% methanol in a 5-fold column volume, and the eluates were collected and tested by HPLC-fractions of detected peaks (at which wavelength 213 nm) were collected and combined. The fractionated products were further separated by liquid preparative chromatography Waters™ (Prep 150 system; Waters™, USA). Column type: Waters Sunfire Prep OBDTM C18 10.0 μm, 19 × 250 mm (Waters™, USA). Mobile phase A was 0.1% formic acid in water, and mobile phase B was methanol at a flow rate of 0.4 ml/min; the column temperature was 40°C, and the injection volume was 20 pl; the elution methods were 0-20 min: 10-100% methanol, 20-30 min: 100% methanol, 30-35 min: 10% methanol.
Compounds are identified using NMR (Pannakal et al. 2023). The isolated compound was dissolved in a suitable deuterated solvent and subsequently analyzed by 1H and 13C NMR on a Bruker Ascend 700 MHz spectrometer (Bruker, USA) at frequencies of 700 and 125 MHz, respectively. The chemical shifts (δ) were given in parts per million (ppm), the coupling constants in Hz. Structure elucidation of the compounds was carried out using 2D NMR experiments 1H-1H COSY, 1H-13C HMBC, 1H-13C HSQC, 1H-15N HMBC.
The activity assay was performed using a plate standoff assay with a concentration of 10 mg/ml, and three treatments were set up to measure the size of the ring of inhibition. To determine the Minimum Inhibitory Concentration (MIC) of compound 44567-1 for its antibacterial effects, the 96-well microtiter plate method outlined by Wiegand et al. in 2008 was utilized. The activity was determined using 96-well plates, with the bacterial solution as the treatment group and the addition of an equal amount of blank LB medium as the test group. The treatments were carried out at 256, 128, 64, 32, 16, 8, 4, 2, 1, 0.5, 0.25 and 0 μg/ml. The test bacteria were cultured overnight and adjusted to an OD600 value of 0.1 before being dispensed into the wells. Each treatment was set up with three replicates after 17 hours of incubation at 37°C to determine the MIC value of P. aeruginosa.
Following the extraction of S. gossypiisoli TRM44567 strain genome, 16 contigs spanning 9,331,940 bp were acquired resulting in annotations for 8,414 CDS, 132 transfer RNA (tRNA) genes. Additional genomic information can be found in Zhang et al. (2021).
A total of 35 BGCs were discovered within the genome of S. gossypiisoli TRM 44567 using the antiSMASH 6.0 tool (Table I). Some of these gene clusters displayed significant correlations with established antibacterials, such as tirandamycin, scabichelin, alkylresorcinol. Furthermore, there were clusters associated with bioactive metabolites like thiocoraline, hopene, desferrioxamin B/E, melanin, spore pigment, and ectoine. A few clusters exhibited minimal to no links with known compounds. The identified clusters encompassed diverse categories such as Terpenes, CDPS, T2PKS, T3PKS, or Hybrids. Based on antiSMASH predictions of the strain TRM 44567 genome sequence, it was determined that the 81.1 kb PKS-NRPS gene cluster 20 in Scaffold 1 shares 100% similarity with the tirandamycin A biosynthetic gene cluster, displaying nearly identical gene composition and alignment. By comparing the gene composition and sequence, the boundaries of cluster 20 were defined, estimating its size to be 56.6 kb. This cluster was further compared to the tirandamycin BGC in Streptomyces sp. 307-9 (Carlson et al. 2009), as illustrated in Fig. 1. Analysis of the functional proteins in the synthesis gene cluster (Table II) indicated a high similarity of around 95% between the synthesis enzymes in strain TRM 44567 and those in Streptomyces sp. 307-9. The gene cluster of strain TRM 44567 contains all the necessary genes for tirandamycin A synthesis, suggesting a high likelihood of the production of tirandamycin A and its natural analogs.
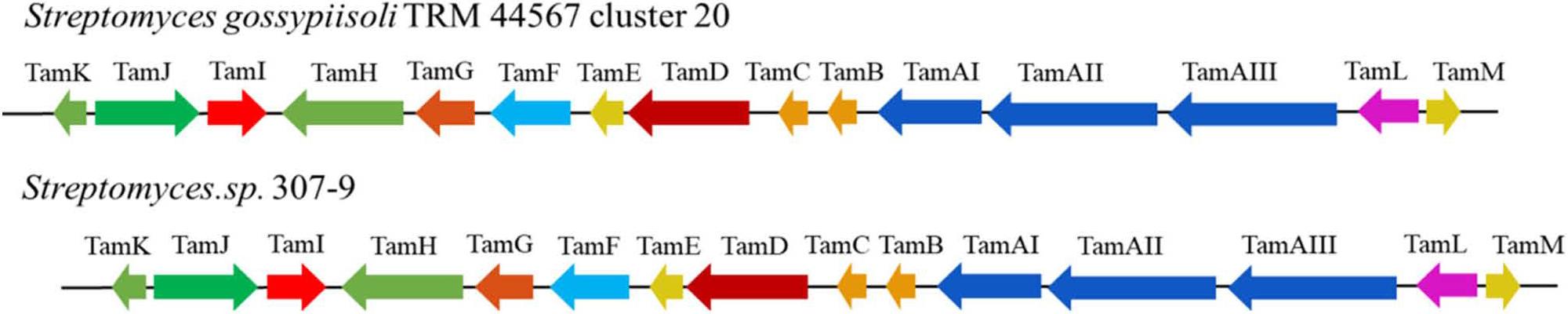
A) Comparison of genetic organization of tirandamycin biosynthetic gene clusters in strain TRM44567 and Streptomyces sp.307–9.
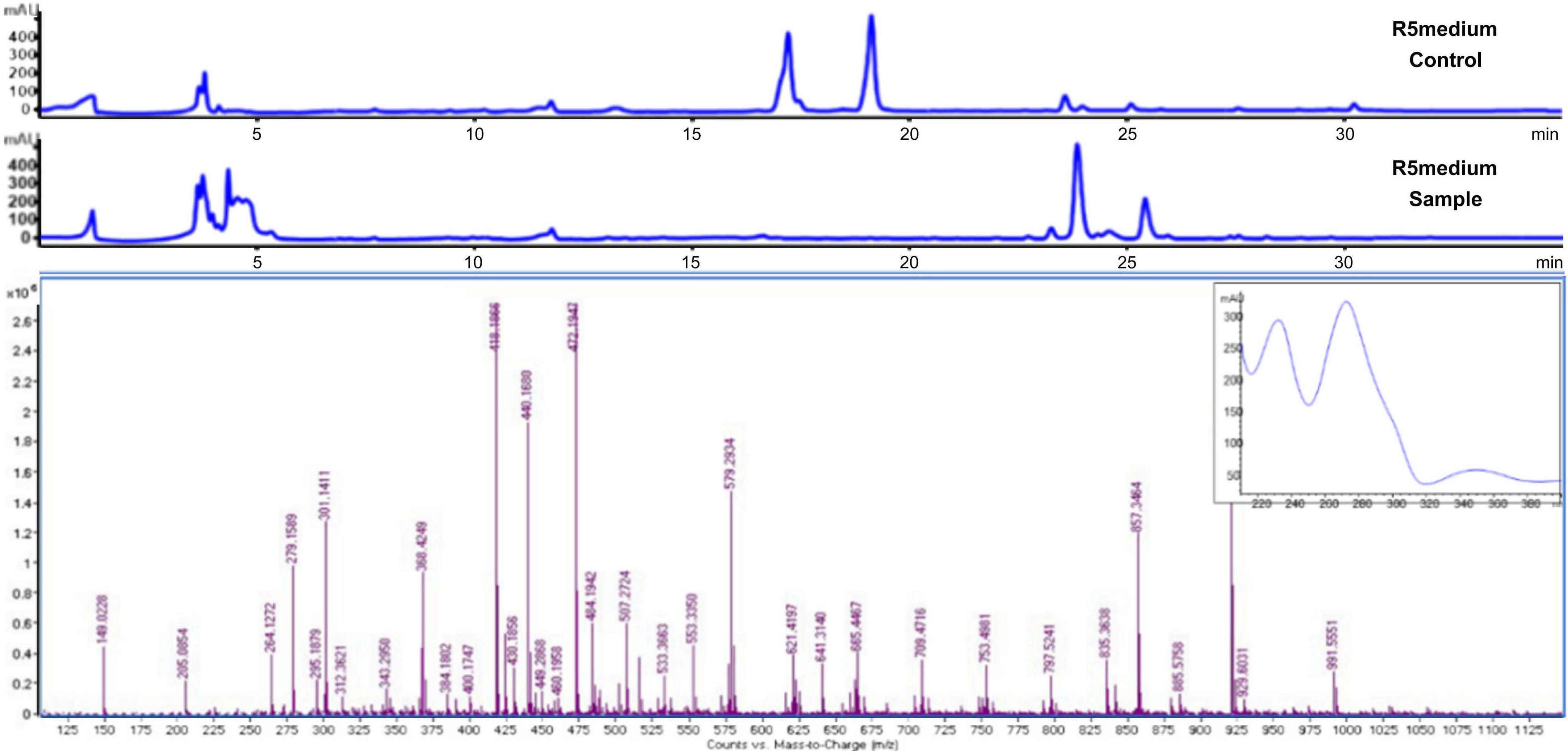
B) UV spectrum and MS peak in the crude extract of strain TRM44567 in R5 medium.
TRM44567 secondary metabolite gene cluster prediction.
| Number | Gene cluster position | Lengths (bp) | Typology | Similar gene clusters | Resemblance (%) |
|---|---|---|---|---|---|
| 1 | 8,773-35,720 | 26,948 | butyrolactone, terpene | 2-methylisoborneol | 75 |
| 2 | 214,388-239,491 | 25,104 | lanthipeptide-class-i | - | - |
| 3 | 667,293-708,315 | 41,023 | other | 7-deoxypactamycin | 11 |
| 4 | 744,740-767,544 | 22,805 | lanthipeptide-class-iv | venezuelin | 50 |
| 5 | 811,611-837,822 | 26,212 | amglyccycl, tyrolactone | acarbose | 14 |
| 6 | 1,098,443-1,154,095 | 55,653 | T1PKS, NRPS | jomthonic acid A/B/C | 5 |
| 7 | 1,329,308-1,339,462 | 10,155 | RiPP-like | - | - |
| 8 | 1,355,940-1,397,127 | 41,188 | T3PKS | gobichelin A/B | 16 |
| 9 | 1,541,462-1,565,687 | 24,226 | lanthipeptide-class-i | - | - |
| 10 | 2,467,823-2,476,751 | 8,929 | RiPP-like | - | - |
| 11 | 2,621,822-2,641,748 | 19,927 | Trpene | albaflavenone | 100 |
| 12 | 3,407,363-3,468,045 | 60,683 | siderophore, lipolanthine, T1PKS, lanthipeptide-class-iii, linaridin | kinamycin | 22 |
| 13 | 3,603,223-3,671,199 | 67,977 | NRPS, NRPS-like | thiocoraline | 60 |
| 14 | 3,715,948-3,744,688 | 28,741 | RiPP-like, terpene | ashimides | 12 |
| 15 | 3,772,034-3,793,450 | 21,417 | butyrolactone | γ-butyrolactone | 100 |
| 16 | 3,859,629-3,900,359 | 40,731 | NRPS, NAPAA | stenothricin | 13 |
| 17 | 4,043,123-4,056,246 | 13,124 | siderophore | atakemycin | 6 |
| 18 | 4,127,840-4,168,065 | 40,226 | NRPS-like | lobosamide A/B/C | 4 |
| 19 | 4,646,047-4,672,724 | 26,678 | terpene | hopene | 92 |
| 20 | 4,773,585-4,854,740 | 81,156 | NRPS, T1PKS | tirandamycin | 100 |
| 21 | 5,069,238-5,079,453 | 10,216 | Ripp-like | informatipeptin | 42 |
| 22 | 276,872-347,373 | 70,502 | T2PKS | prejadomycin/gaudimyc in | 29 |
| 23 | 776,140-787,912 | 11,773 | siderophore | desferrioxamin B/E | 83 |
| 24 | 905,539-914,564 | 9,026 | melanin | melanin | 60 |
| 25 | 1,263,341-1,304,207 | 40,867 | T3PKS | s56-p1 | 11 |
| 26 | 1,631,756-1,704,267 | 72,512 | T2PKS | spore pigment | 83 |
| 27 | 2,082,866-2,093,270 | 10,405 | ectoine | ectoine | 100 |
| 28 | 2,295,368-2,314,558 | 19,191 | CDPS | - | - |
| 29 | 2,369,855-2,402,869 | 33,015 | NAPAA | meilingmycin | 2 |
| 30 | 2,970,457-3,010,999 | 40,543 | T3PKS | herboxidiene | 7 |
| 31 | 3094,635-3,115,642 | 21,008 | terpene | desotamide | 9 |
| 32 | 3,430,099-3,508,231 | 78,133 | Melanin, T1PKS, NRPS | scabichelin | 100 |
| 33 | 3,579,258-3,620,409 | 41,152 | T3PKS | alkylresorcinol | 100 |
| 34 | 3,887,970-3,908,680 | 20,711 | CDPS | - | - |
| 35 | 3,920,922-3,964,188 | 43,267 | NRPS, NAPAA | stenothricin | 13 |
Tirandamycin biosynthesis-related gene function prediction and consistency analysis.
| Number | Gene lengths (bp) | Coding protein | Homologous strain | Resemblance (%) |
|---|---|---|---|---|
| 4283 | 645 | TamK (TetR family regulator) | Streptomyces sp. 307-9 | 99 |
| 4284 | 1494 | TamJ (efflux pump) | Streptomyces sp. 307-9 | 99 |
| 4285 | 1236 | TamI (cytochrome P450) | Streptomyces sp. 307-9 | 99 |
| 4286 | 2814 | TamH (LuxR family regulator) | Streptomyces sp. 307-9 | 97 |
| 4287 | 2094 | TamG (DNA helicase) | Streptomyces sp. 307-9 | 99 |
| 4288 | 1539 | TamF (Prenyltransferase) | Streptomyces sp. 307-9 | 97 |
| 4289 | 831 | TamE (glucosidase) | Streptomyces sp. 307-9 | 98 |
| 4290 | 3165 | TamD (NRPS) | Streptomyces sp. 307-9 | 97 |
| 4291 | 831 | TamC (hypothetical protein) | Streptomyces sp. 307-9 | 97 |
| 4292 | 777 | TamB (alpha/beta fold hydrolase) | Streptomyces sp. 307-9 | 98 |
| 4293 | 8400 | TamAIII | Streptomyces sp. 307-9 | 95 |
| 4294 | 10131 | TamAII | Streptomyces sp. 307-9 | 94 |
| 4295 | 17490 | TamAI | Streptomyces sp. 307-9 | 95 |
| 4296 | 1503 | TamL (FAD-binding oxidoreductase) | Streptomyces sp. 307-9 | 98 |
| 4297 | 234 | TamM (4’-phosphopantetheinyl transferase) | Streptomyces. sp. 307-9 | 97 |
| 4291 | 831 | TamC (hypothetical protein) | Streptomyces sp. 307-9 | 97 |
Upon examination of the reference, it was noted that tirandamycin displayed distinct UV absorptions at 213 nm and 351 nm (Rateb et al. 2014), with a theoretical high-resolution mass spectral signal [M + H] + of 418.1792. The identifiable UV absorption and mass spectral peaks of the desired compounds were specifically detected in the 01, 08, 09, and R5 mediums, with the highest concentrations observed in the R5 fermentation medium, and it was determined by mass spectrometry that R5 can produce tandamycin, as shown in Fig. 1 and Fig. S2. Analysis using Q-Tof results revealed a peak with similar UV absorption and mass spectral signal [M + H] + of 418.1866 appearing in the liquid phase around 24 minutes, suggesting a potential identification of tirandamycin A.
Pre-fermentation using R5 medium and obtaining metabolites 44567-1 using prepared liquid phase. The compound 44567-1 was found as yellow solids. 1H NMR (700 MHz, Methanol-d4) δ 7.56 (d, J = 15.7 Hz, 1H, H-3), 7.20 (d, J = 15.6 Hz, 1H, H-2), 6.20 (d, J = 10.1 Hz, 1H, H-5), 3.98 (d, J = 6.1 Hz, 1H, H-9), 3.80 (s, 2H, H-5’), 3.76 (dd, J = 11.5, 2.1 Hz, 1H, H-7), 3.32 (s, 1H, H-11), 2.95 (m, 1H, H-6), 1.95 (m, 1H, H-8), 1.92 (s, 3H, H-15), 1.52 (s, 3H, H-14), 1.46 (s, 3H, H-18), 1.15 (d, J = 6.9 Hz, 3H, H-16), 0.72 (d, J = 7.0 Hz, 3H, H-17); 13C NMR (700 MHz, Methanol-d4) δ203.77(C-10), 176.36(C-2’), 176.27(C-1), 150.23(C-3), 144.98(C-5), 136.15(C-4), 117.84(C-2), 98.07(C-13), 80.16(C-9), 78.02(C-7), 62.13(C-11), 58.15(C-12), 49.85(C-5’), 35.86(C-8), 35.72(C-6), 22.95(C-14), 17.30(C-16), 15.84(C-18), 12.40(C-15), 11.67(C-17). Related results for Table SII and Fig. S3-S4, for other relevant results, in comparison to tirandamycins D, tirandamycins A was found to possess a C-11/C-12 Z double bond, as indicated by the chemical shift values observed at position 11 (δH 3.32, δC 62.125) and position 12 (δC 58.15), along with the adjacent carbonyl at C-10 (δC 203.77). This assignment is further supported by various correlations, including a long-range COSY coupling between H-11 and H-18, as well as HMBC correlations from H-11 to C-13 and C-18, from H-18 to C-11 and C-12, and from H-14 to C-12. Additionally, HSQC mapping data align with the characteristics of tirandamycins A, distinguishing them from tirandamycins C, as illustrated in Fig. S5 and S7. The mapping tool used was MestReNova (Mestrelab Research S.L.U., Spain). Based on the spectral characteristics obtained and the data from the literature (Reusser et al. 1970; Reusser et al. 1976), it was deduced that the isolated compound 44567-1 is tirandamycin. The chemical nature of compound 44567-1 was confirmed to be that of tirandamycin with molecular formulae C22H27NO7. The compounds were drawn using ChemDraw 19.0 (PerkinElmer Informatics, Inc., USA) (Fig. 2).
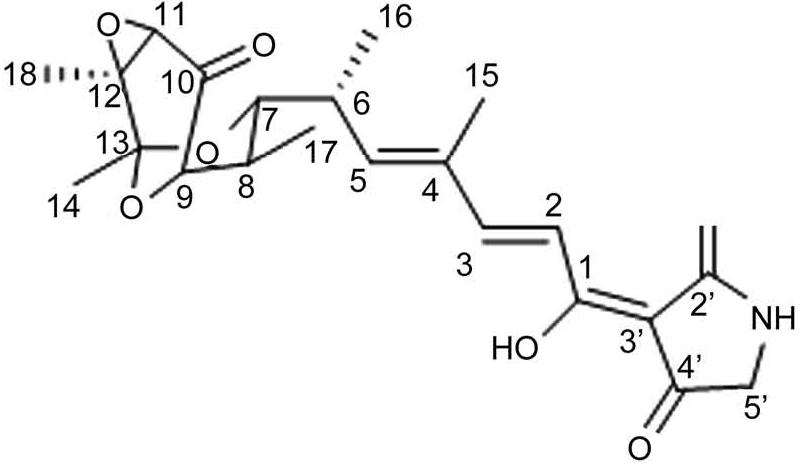
The structure of tirandamycin.
The activity assay was performed using a plate standoff assay using a concentration of 10 mg/ml, and three treatments were set up to measure the size of the ring of inhibition. As shown in Table III and Fig. S2, the purified compounds 44567-1 could inhibit P. aeruginosa (ATCC® 27853), compounds 44567-1 was shown to be more efficient against P. aeruginosa, with MICs of 8 μg/ml (Table IV).
Comparison of the antibacterial activities of 44567-1 against different targets.
| Number | Target organism | Bacteriostatic activity screening (diameter of inhibition circle) |
|---|---|---|
| 1 | Escherichia coli ATCC® 25922™ | - |
| 2 | Staphylococcus aureus ATCC® 25923™ | 3.0 ± 0.2 mm |
| 3 | Enterococcus faecalis ATCC® 29212™ | - |
| 4 | Klebsiella pneumoniae ATCC® 10031™ | - |
| 5 | Acinetobacter baumannii ATCC® 19606™ | - |
| 6 | Pseudomonas aeruginosa ATCC® 27853™ | 32.0 ± 0.3 mm |
| 7 | Salmonella choleraesu | - |
| 8 | Salmonella castellani | - |
| 9 | Candida albicans ATCC® 64550™ | - |
Our previous study based on the phylogenetic tree of 16S rRNA gene and whole genome sequences showed that strain TRM 44567 formed a unique cluster (Zhang et al. 2021). In recent years, due to the optimization of sequencing costs and the continuous improvement of sequencing methods, the microbial genomic information included in the databases has become richer and richer, and the genomic information plays a very important role in the mining of natural products (Crüsemann et al. 2021). Bioinformatics analyses show that each bacterial strain synthesizes more than 20 natural products, but about 90% of these natural products have not been studied yet, and therefore have great potential for development (Cragg et al. 2013). Genome mining is a natural product mining strategy based on genome sequences and biosynthetic pathways. At the same time, it can well correlate the chemical structures of secondary metabolites with their biosynthetic pathways, which is convenient for subsequent biosynthesis or combinatorial biosynthesis studies. This study obtained the complete genome sequence of strain TRM 44567 with an average GC content of 70.74%. The strain was found to possess 35 BGCs and the secondary metabolites identified in TRM 44567 and similar strains mainly belonged to the classes of PKS, NRPS, NISiderophore, and Lanthipeptide. The metabolic potential of these strains was found to be consistent. Strain TRM 44567 exhibited unique biosynthetic genes with the potential to produce new metabolites such as tirandamycin compared to other similar strains (Carlson et al. 2009). This suggests that strain TRM 44567 has a good metabolic capacity, and further exploration of its biosynthetic genes is important for discovering new metabolites or antibiotics.
Currently, it is widely accepted that a large proportion of microbial gene clusters are silenced under standard fermentation conditions. Silenced genes are the biosynthetic genes that are not directly related to growth or differentiation and are usually silenced or transcriptionally repressed to conserve the microbial energy sources (Louca et al. 2018). The “OSMAC” (one strain many compounds) strategy, proposed by prof. Zeeck and his colleagues, is a way to stimulate the expression of secondary metabolite genes by changing the culture conditions, which has been considered the simplest and most effective strategy to activate the silenced genes and has been widely used in secondary metabolite research (Schiewe and Zeeck 1999). For example, in this study, the production of the metabolite tirandamycin A was successfully determined by altering the medium composition of strain TRM 44567 combined with mass spectrometry release analysis.
Tirandamycin B and tirandamycin A were first isolated from S. tirandis and Streptomyces flaveolus in the 1970s, the antibiotics belonging to the tetramic acid (2,4-pyrrolidinedione) class, initially identified as inhibitors of bacterial RNA polymerase and HIV protease activity (Dhavan et al. 2016). Tuske et al. (2005) utilized X-ray to analyze the three-dimensional crystal structure of the structurally similar antibiotic rifamycin in complex with RNA polymerase. They elucidated the mode and mechanism of inhibition, which differs from rifampicin’s action on bacterial RNA polymerases. These findings suggest potential for tirandamycin in drug development, serving as a lead compound for molecular manipulation and combinatorial synthesis to create tetramic acid ring structures with enhanced or novel drug activity.
In the present study, tirandamycin was newly discovered from the desert-sourced Streptomyces sp. TRM 44567, which exhibits P. aeruginosa activity. Comparing the seven tirandamycin congeners reported in the literature by Rateb et al. (2014) we found that tirandamycin A had inhibitory activity against P. aeruginosa. Notably, we found that its homologues could serve as lead compounds for antifilarial drugs. Furthermore, Streptomycetes sp. 17944, which has been reported to produce tirandamycin, emerged as the most promising candidate for its production. Tirandamycin A was produced in the ISP2 medium, while the medium utilized in this study was the R5 medium. It is important to note that varying growth environments, potentially due to strain specificity, may influence different strains producing the same antibiotic.
In this study, we used genomic analysis to determine that strain TRM 44567 can produce multiple types of metabolites. The gene cluster analysis was used to determine its ability to produce tirandamycin. The best solid fermentation medium for this strain was obtained as R5 medium, and the compound, tirandamycin, was isolated by mass spectrometry coupled with preparative isolation. These findings suggest the potential for tirandamycin in drug development, serving as a lead compound for molecular manipulation and combinatorial synthesis to create tetramic acid ring structures with enhanced or novel drug activity. The present study enriches the small molecule compounds library and identifies the compounds’ novel activities.