
Campylobacter genus is comprised of Gram-negative, non-spore-forming rods and spiral and coccoid/spherical bacteria (Kreling et al. 2020). Campylobacter species such as C. hyointestinalis, C. jejuni, C. fetus, C. coli, C. upsaliensis, and C. lari are important human pathogens (Hlashwayo et al. 2021). They cause food and water-borne enteric infections worldwide, which occur after the consumption of non-chlorinated well water or contaminated surface water (Butzler 2004; Abulreesh et al. 2006; 2017), undercooked red meat or poultry products (Khan et al. 2018a; Ansarifar et al. 2023), and unpasteurized milk (Igwaran and Okoh 2019). Direct contact with infected individuals might also transmit campylobacteriosis. Similarly, nosocomial infection can occur, but reports of congenital transmission are rare. Children can acquire campylobacteriosis from immature diarrhetic animals (Kreling et al. 2020).
During the past three decades, Campylobacter species have emerged as important clinical pathogens of acute enteritis in Western countries. C. coli and C. jejuni are particularly important regarding gastrointestinal infections. Moreover, a link has been established between Guillain-Barre syndrome (GBS) and C. jejuni infection, further enhancing this species’ importance (Kreling et al. 2020). Campylobacteriosis occurrence is significantly higher in developing countries than in developed nations, and Campylobacter-associated infection affects a large number of children in developing countries. Community-based investigations in developing countries have revealed that 60,000/100,000 children (aged < 5 years) suffer from campylobacteriosis, which establishes it as a pediatric disease (Coker et al. 2002; Kreling et al. 2020). Human campylobacteriosis cases (> 90%) in developed countries mainly occur in the summer season with the increased consumption of undercooked meats at outdoor facilities. Campylobacteriosis could occur in individuals of all ages, but the infection rate is high in children (< 4 years) and young adults (15-44 years) (García and Heredia 2009). A relatively low infectious dose of C. jejuni (500-800 organisms) has been estimated in humans (Nachamkin et al. 2008). Standard Campylobacter spp. isolation from food and water is performed through selective enrichment and selective plating. Public Health England (PHE), US Food and Drug Administration (FDA), and International Organization for Standardization (ISO) have developed different Campylobacter spp. detection methods involving specific sample preparation steps, selective plating media, and enrichment broths.
This study overviews standard culture-based methods (ISO, PHE, and USFDA) for isolating Campylobacter spp. from food and water samples. It assessed the efficiency of culture media, pre-enrichment, selective enrichment, and selective plating media for detecting and monitoring Campylobacter spp. The review also elaborates on novel chromogenic culture media (advantages and constraints) to differentiate Campylobacter spp. in different samples (food and water). The limitations associated with culture-based detection of viable but non-culturable (VBNC) cells are discussed as well. Moreover, the review highlights alternative techniques and improvements for precise and efficient detection of Campylobacter spp. Overall, the study provides detailed insights regarding detecting Campylobacter spp. in food and water samples.
Standard Campylobacter spp. detection in food and water samples often requires pre-enrichment followed by selective enrichment (at certain intervals and temperatures) and selective plating. The complete isolation process (identification and confirmation) could take up to 7 days (Fig. 1). Campylobacter spp. detection procedures of PHE (2014), ISO (ISO 2017), and US-FDA Bacteriological Analytical Manual (FDA-BAM) (FDA 2001) share some similarities. However, FDA-BAM recommends the same Campylobacter spp. isolation procedure for all types of samples (shellfish, milk, cheese, and water). ISO recommends three different methods according to the Campylobacter spp. contamination levels and background bacteria in food and water samples. PHE also employs the same procedure for different surface water and environmental samples.
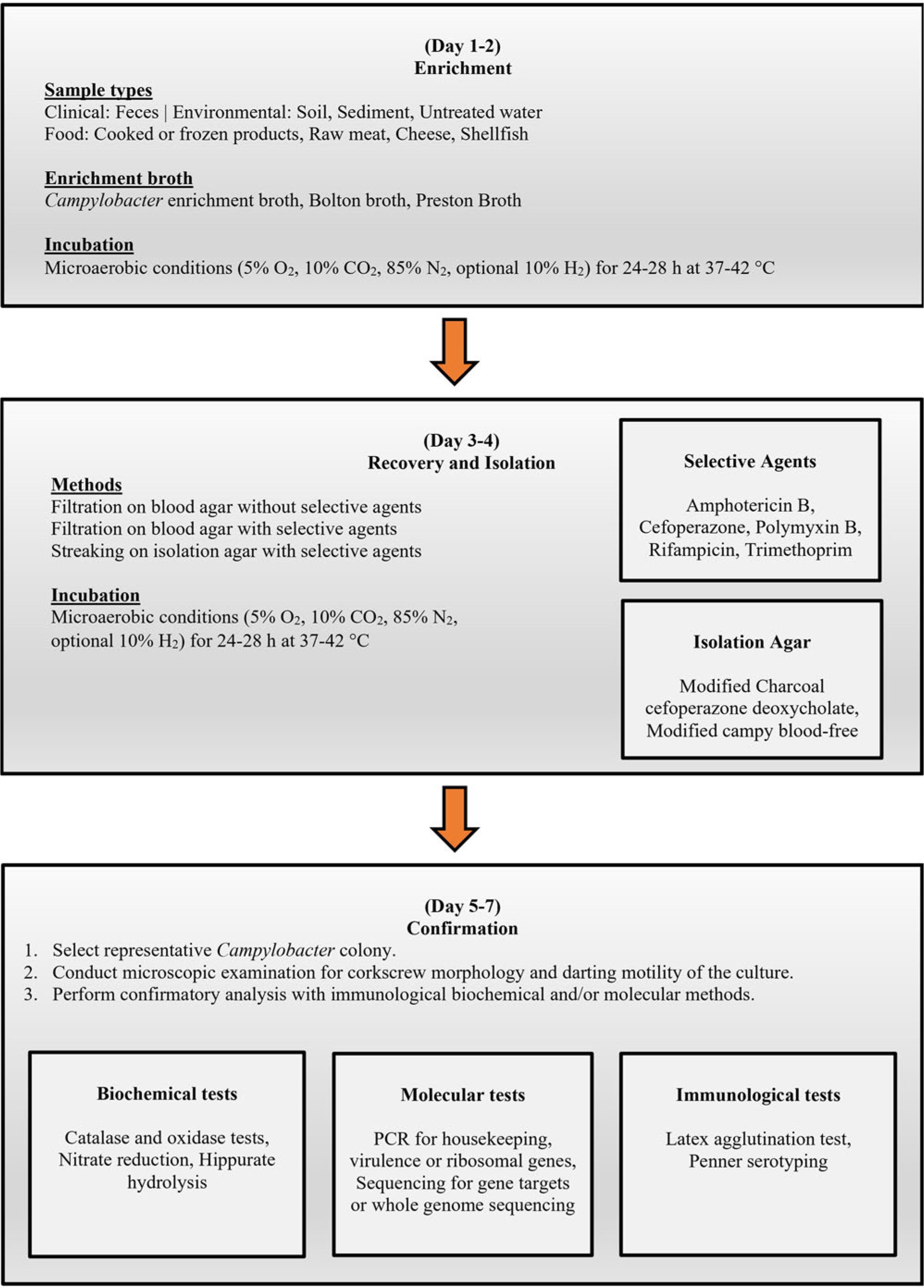
General flow chart showing the phases and the time required for the routine detection and identification/confirmation of Campylobacter spp. in food, water, and feces.
FDA has established five processing procedures before Campylobacter spp. detection from food and water samples (FDA 2001). Sample preparation differs, whereas the detection procedure remains similar for all sample types and food sample/homogenate quantiles. Campylobacter spp. isolation from most foods [vegetables, poultry (Fig. 2), water (Fig. 3), shellfish (Fig. 4), milk (Fig. 5), and cheese (Fig. 6)] require a Bolton broth-based pre-enrichment under microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%). Pre-enrichment temperature and incubation time could vary among various sample types (Fig. 2-6), which is followed by enrichment (20-44 hours, 42°C) under microaerobic conditions. The enrichment culture is streaked on an FDA-recommended selective plating media (Abeyta-Hunt-Bark agar (AHB), modified charcoal cefoperazone deoxycholate agar (mCCDA), or Abeyta-Hunt-Bark without antibiotics). Then, plates are incubated under microaerobic conditions (24-48 hours, 42°C) (Table I). FDA protocol for the identification and confirmation of presumptive Campylobacter spp. colonies often relies on biochemical features of Campylobacter spp. (FDA 2001). Initially, suspected colonies are examined for oxidase and catalase and oxidase followed by physiological and biochemical tests such as nitrate reduction, Hippurate hydrolysis, reaction on triple sugar iron agar (TSI), nalidixic acid resistance, growth at different temperatures (42°C, 35-37°C, and 25°C), and growth in glycerin. Table II enlists the biochemical and physiological features of different Campylobacter spp. to confirm their identification.
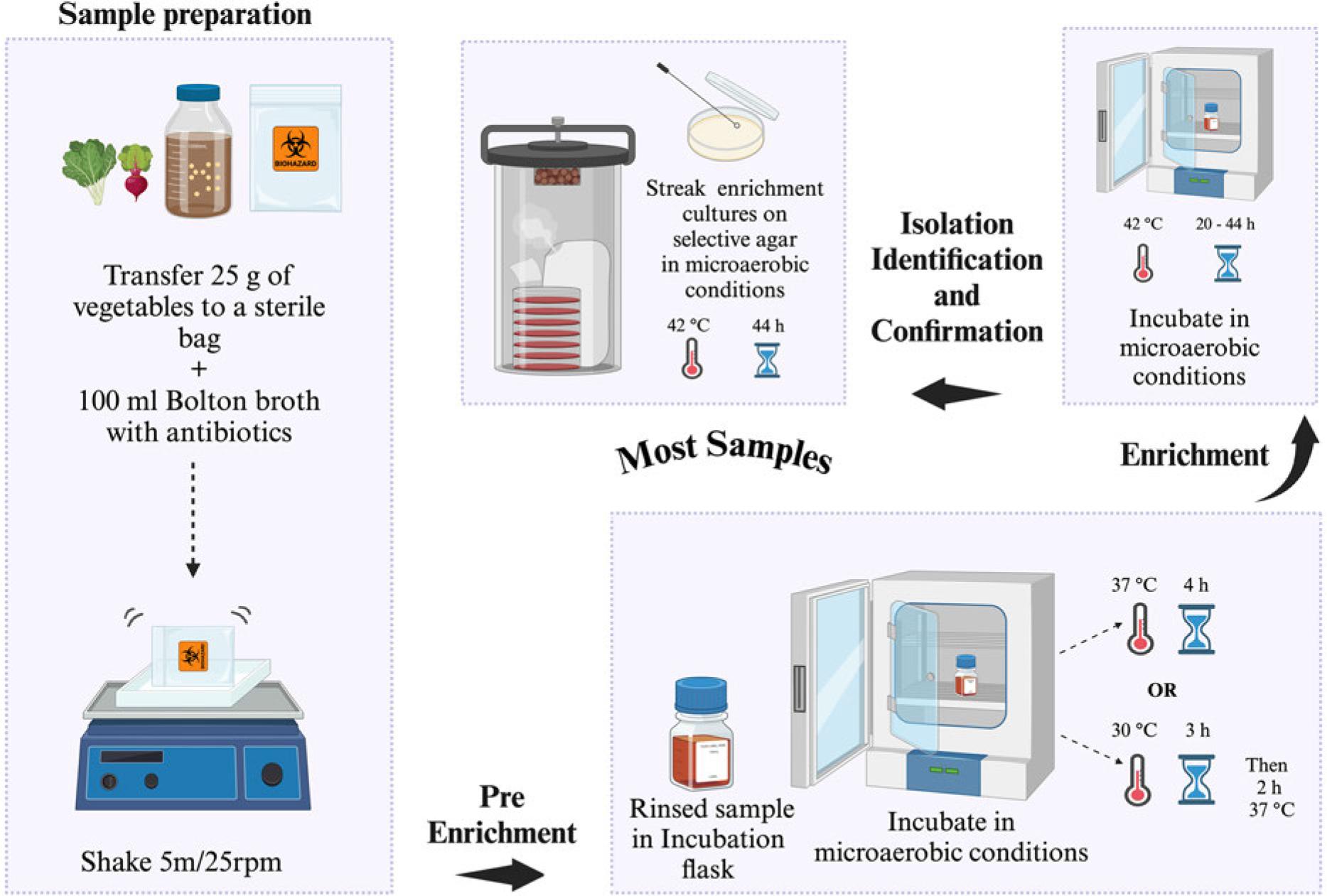
US FDA-BAM procedure for the isolation of Campylobacter spp. from most of the food samples (e.g., vegetables/raw meat). Selective agar recommended for isolation: modified charcoal cefoperazone deoxycholate agar (mCCDA) or Abeyta-Hunt-Bark agar, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
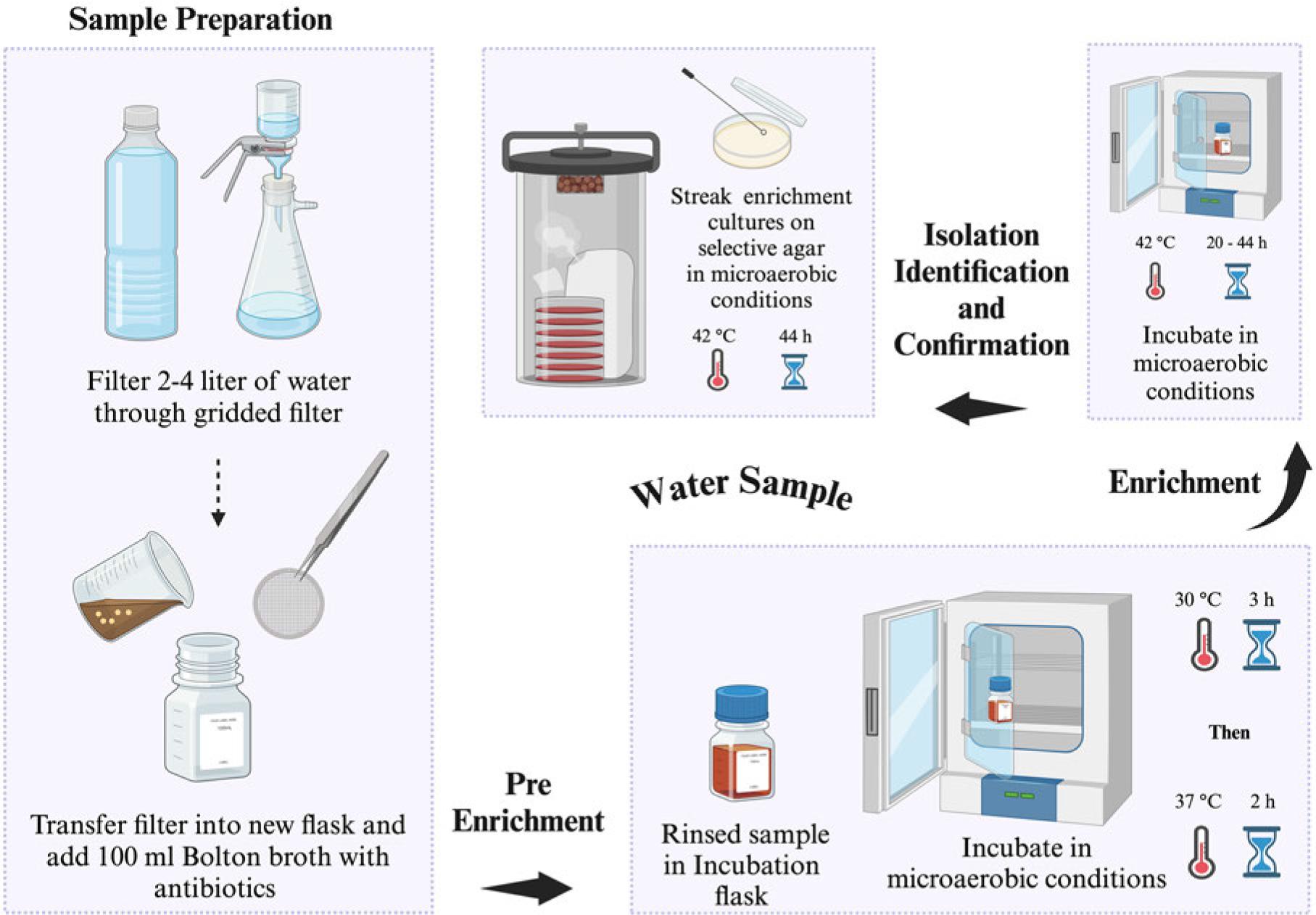
US FDA-BAM procedure for the isolation of Campylobacter spp. from drinking water. Selective agar recommended for isolation: modified charcoal cefoperazone deoxycholate agar (mCCDA) or Abeyta-Hunt-Bark agar, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
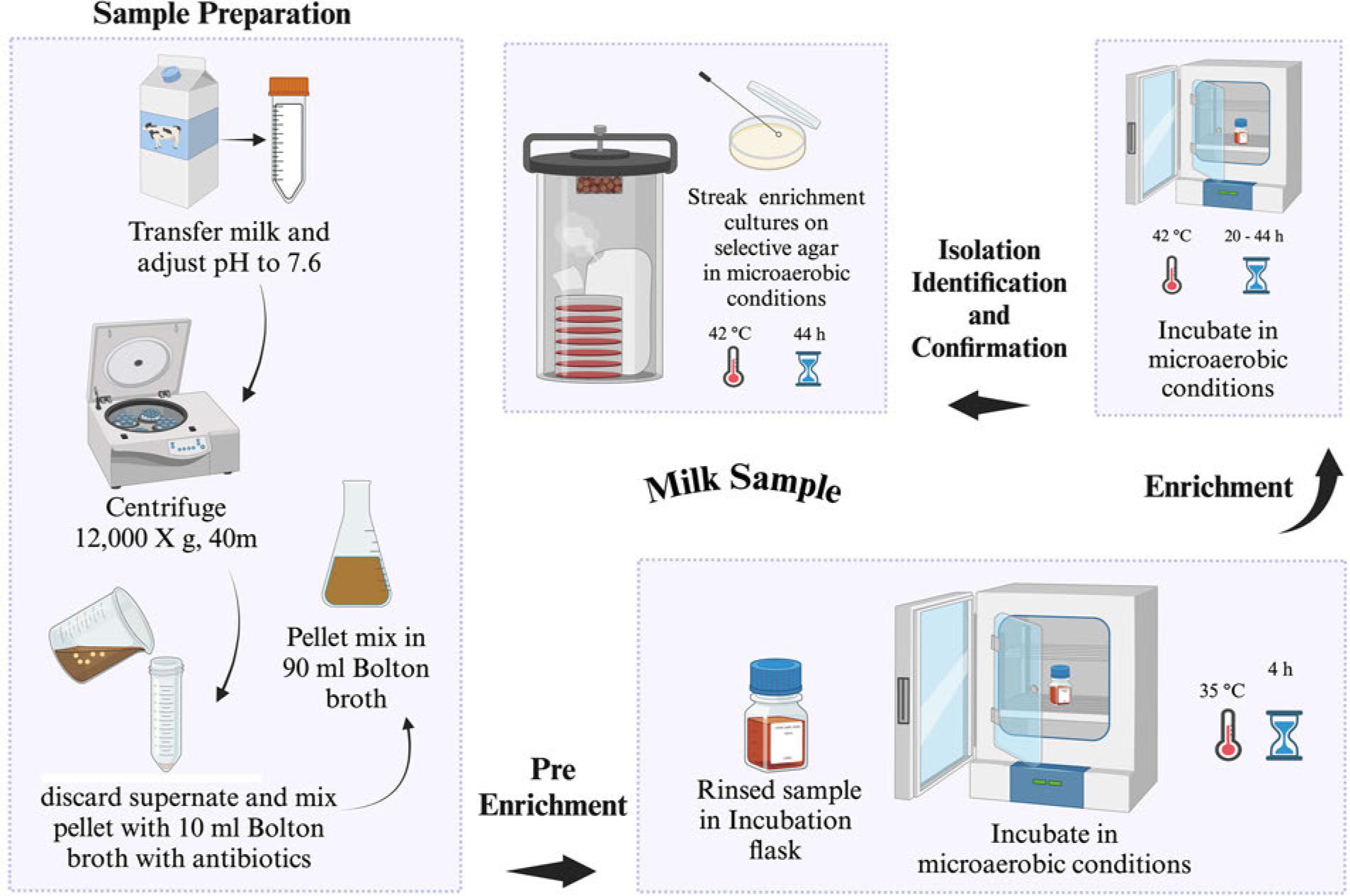
US FDA-BAM procedure for the isolation of Campylobacter spp. from milk. Selective agar recommended for isolation: modified charcoal cefoperazone deoxycholate agar (mCCDA) or Abeyta-Hunt-Bark agar, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
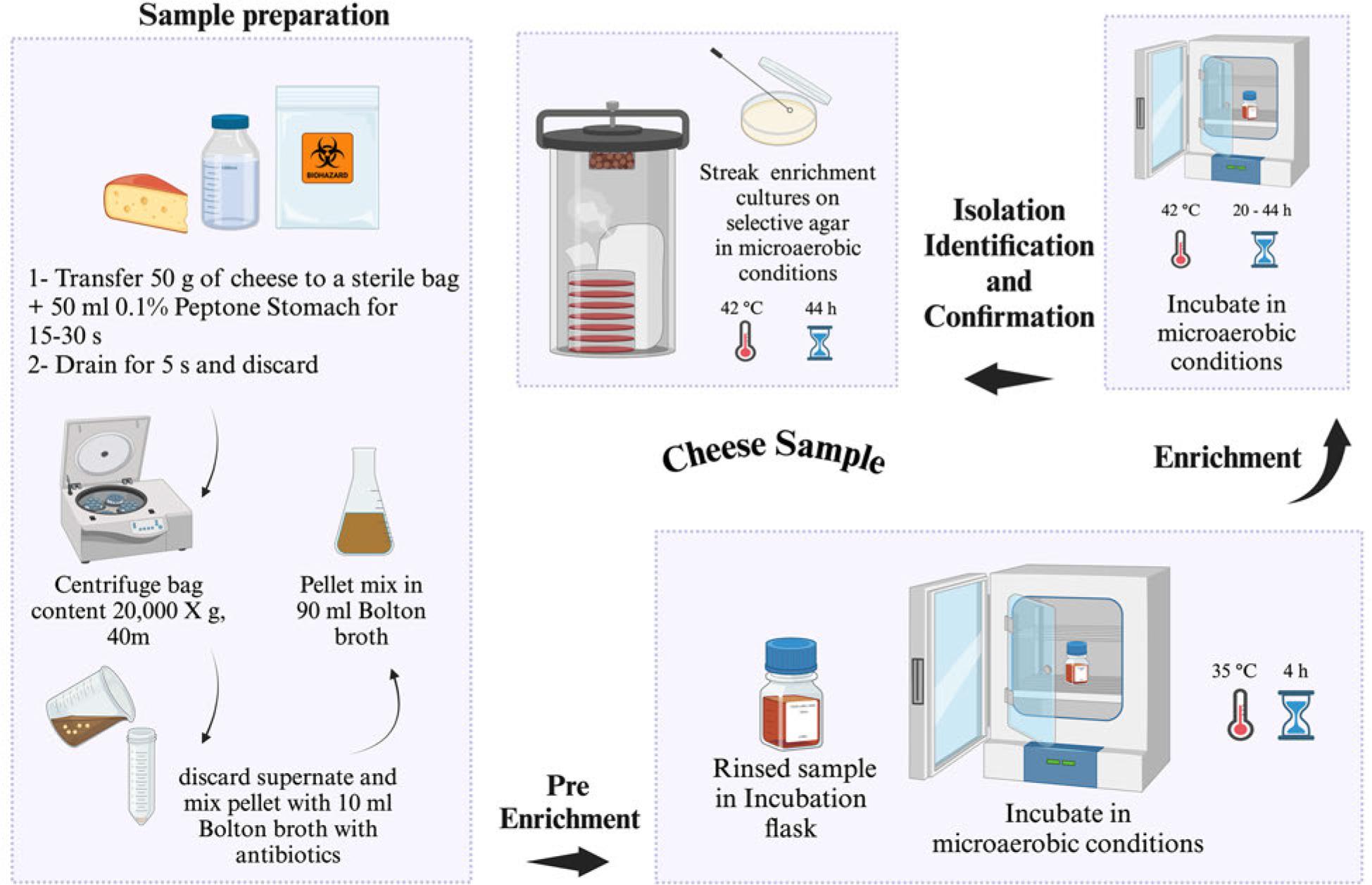
US FDA-BAM procedure for the isolation of Campylobacter spp. from cheese. Selective agar recommended for isolation: modified charcoal cefoperazone deoxycholate agar (mCCDA) or Abeyta-Hunt-Bark agar, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
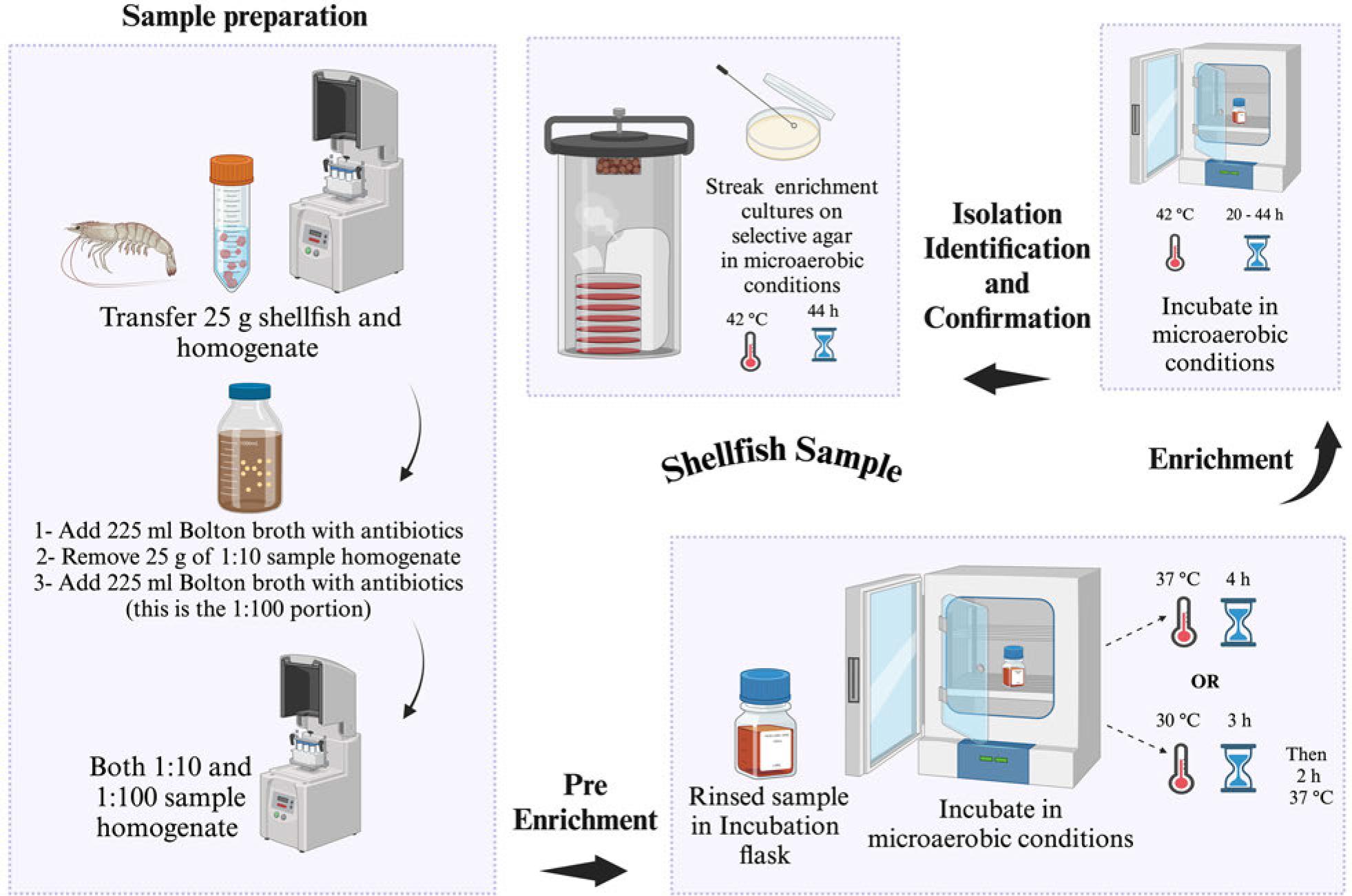
US FDA-BAM procedure for the isolation of Campylobacter spp. from shellfish. Selective agar recommended for isolation: modified charcoal cefoperazone deoxycholate agar (mCCDA) or Abeyta-Hunt-Bark agar, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
| Organization | Sample type | Preenrichment | Enrichment | Selective plating | Reference |
|---|---|---|---|---|---|
| FDA | All sample types | Bolton broth | Bolton | mCCDA and/or Abeyta-Hunt-Bark agar | FDA 2001 |
| ISO | Samples with low campylobacters versus low background bacteria | Bolton broth | Bolton broth | mCCDA + other media of choice | ISO 2017 |
| ISO | Samples with low campylobacters versus high background bacteria | Not required | Preston broth | mCCDA + other media of choice | ISO 2017 |
| ISO | Samples with high campylobacters | Not required | Not required | mCCDA | ISO 2017 |
| ISO | All samples (colony count) | Not required | Not required | mCCDA | ISO 2017 |
| PHE | All food samples | Bolton broth | Bolton broth | mCCDA | PHE 2014 |
mCCDA - modified charcoal cefoperazone deoxycholate agar
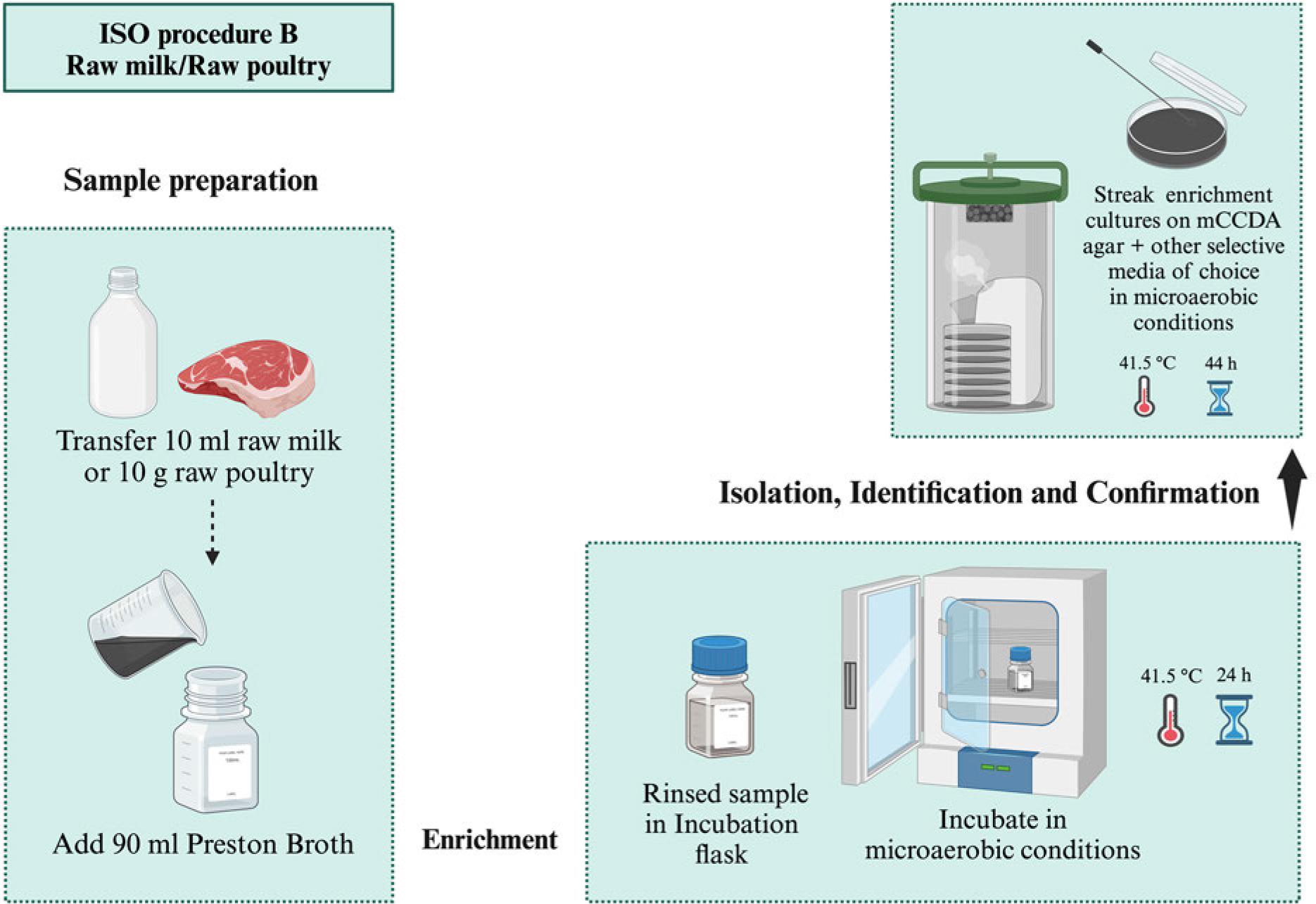
ISO proposed three procedures for Campylobacter spp. isolation according to their contamination levels and background bacteria (ISO 2017). Samples with lower numbers of Campylobacter spp. and background bacteria are subjected to Bolton broth-based pre-enrichment (4-6 hours, 37°C) followed by enrichment under a microaerobic atmosphere (44 hours, 41.5°C). Then, selective plating is carried out using mCCDA and another media of choice, followed by incubation (44 hours, 41.5°C) (Procedure A, Fig. 7) (Table I). Preston broth is used for the selective enrichment (24 hours, 41.5°C) of Campylobacter spp. in samples with their lower numbers and high background bacteria followed by mCCDA mediabased selective plating as mentioned in procedure A (Procedure B, Fig. 8) (Table I). The samples with high Campylobacter spp. levels are subjected to direct plating on selective agar (mCCDA) without enrichment steps (Procedure C, Fig. 9) (Table I). ISO recommends a colony count method for Campylobacter spp. enumeration in food and water samples (ISO 2017). It is carried out by spreading water and milk samples (1 ml) or food homogenate (1 ml) on a well-dried mCCDA plate surface. This approach can also be followed for samples’ serial dilutions. Then, the plates are incubated (40-44 hours, 41.5°C) under microaerobic conditions without pre-enrichment or enrichment steps (Fig. 10). ISO recommends the microscopic confirmation of suspected Campylobacter spp. colonies through motility and morphological appearance (ISO 2017). Moreover, aerobic growth (25°C) and oxidase activity should also be analyzed. Other biochemical tests can be performed as well to differentiate Campylobacter spp. colonies (Table II). ISO protocols also suggest PCR-based molecular identification and confirmation of presumptive Campylobacter spp. colonies (ISO 2017).
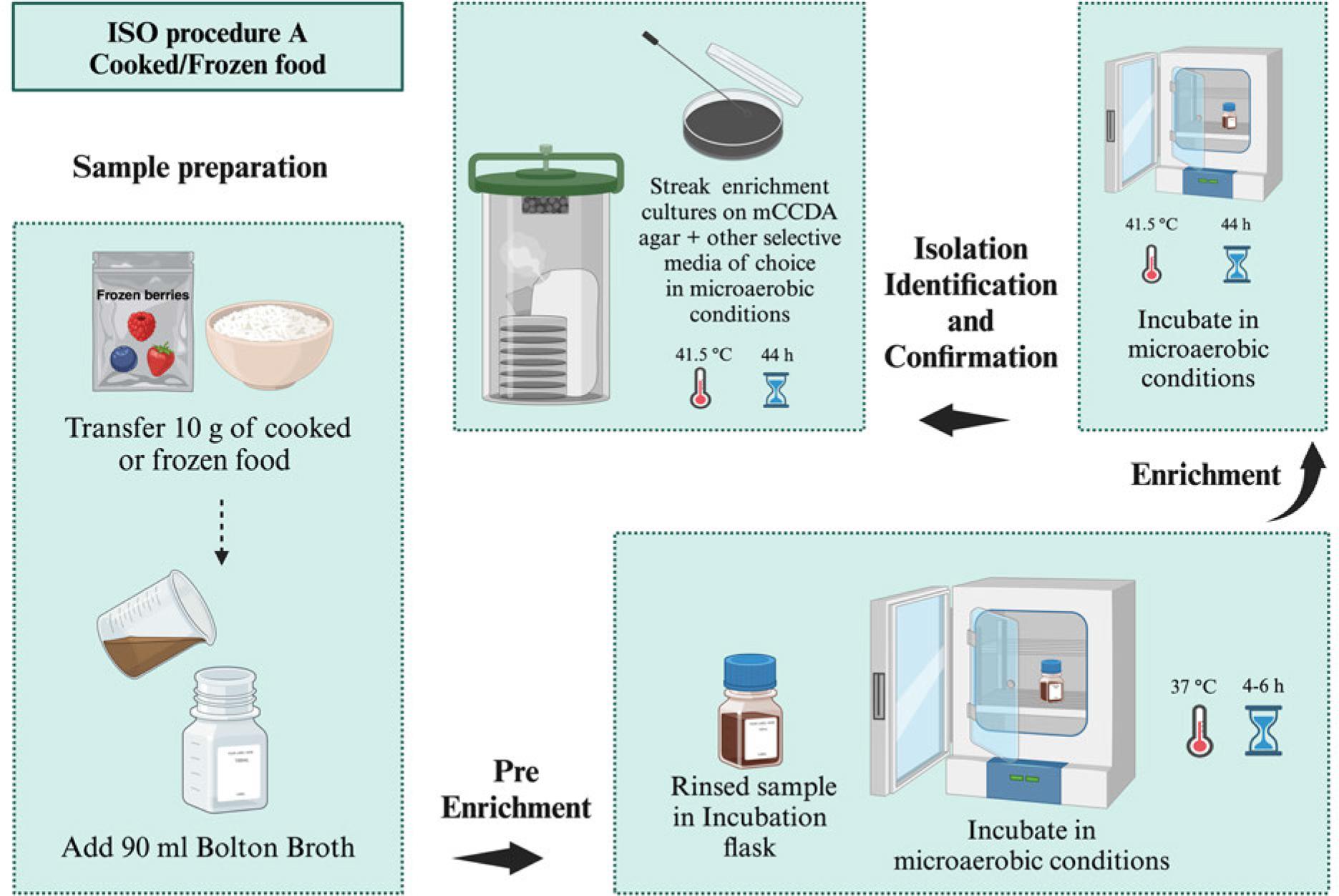
ISO procedure A for the isolation of Campylobacter spp. from samples with low number of Campylobacter and low number of background bacteria (e.g., cooked/frozen food), microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).ISO procedure B for the isolation of Campylobacter spp. from samples with a low number of Campylobacter versus a high number of background bacteria (e.g., milk/raw meat), microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%). ISO procedure B for the isolation of Campylobacter spp. from samples with a low number of Campylobacter versus a high number of background bacteria (e.g., milk/raw meat), microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
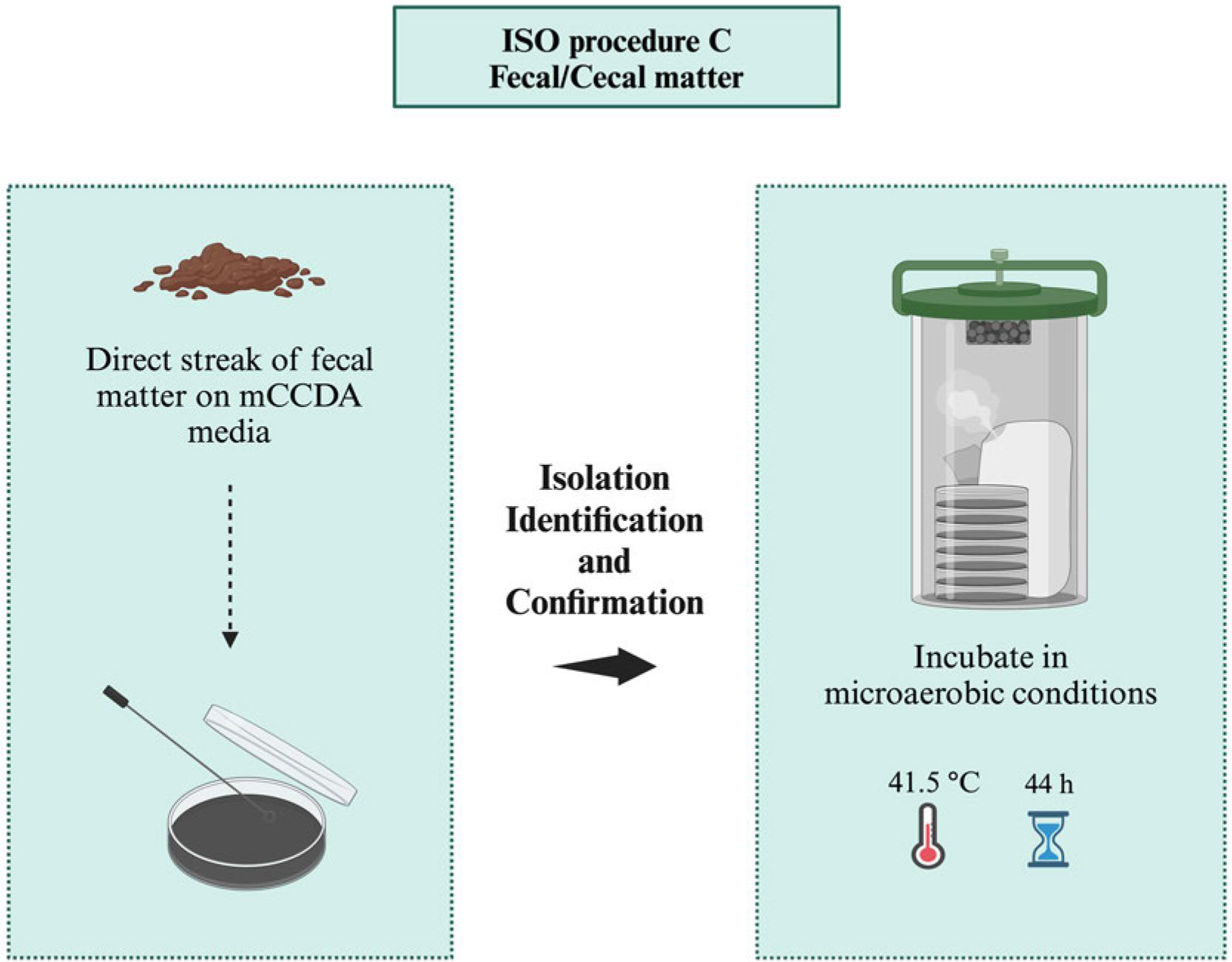
ISO procedure C for the isolation of Campylobacter spp. from samples with high number of Campylobacter (e.g., fecal/raw meat), microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
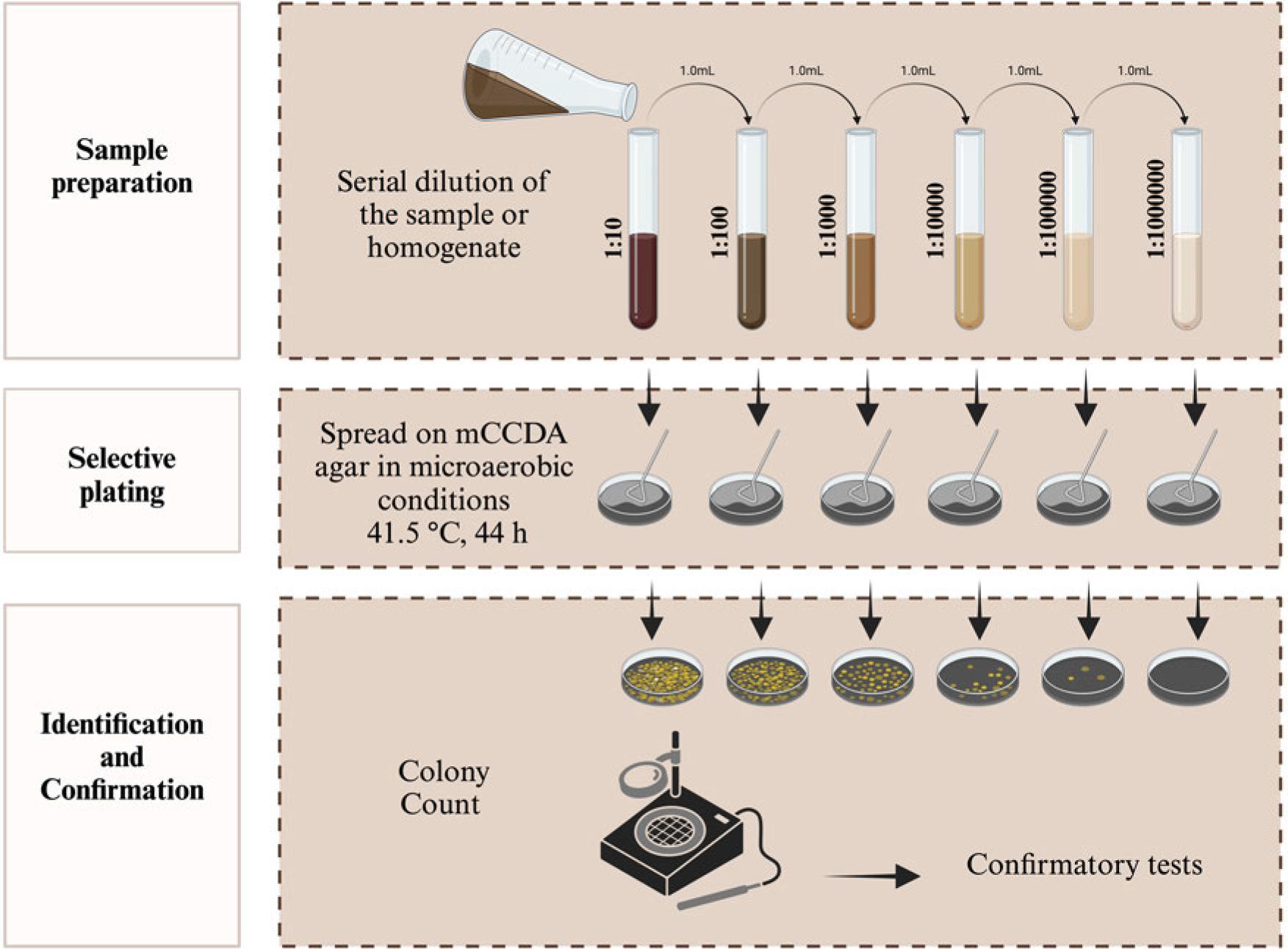
ISO procedure for the colony count of Campylobacter spp. in all food samples, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
| Species | Oxidase | Catalase | Growth at | Hippurate hydrolysis | Aerobic growth | Nitrate reduction | H2S | Sensitive to | TSI | Growth 1% glycine | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25°C | 37°C | 42°C | Cephalothin | NA | |||||||||
| Campylobacter jejuni | + | + | − | + | + | + | − | + | + | R | S | − | + |
| Campylobacter coli | + | + | − | + | + | − | − | + | + | R | S | − | + |
| Campylobacter fetus | + | + | + | + | (V) | − | − | + | V | S | R | − | + |
| Campylobacter upsaliensis | + | −/W | − | + | V | − | − | + | U | S | S | − | − |
| Campylobacter cinaedi | + | − | − | + | − | − | − | + | + | S | S | − | + |
| Campylobacter lari | + | + | − | + | + | − | − | + | + | R | R | − | + |
| Campylobacter fennellia | + | − | − | + | − | − | − | − | + | S | S | − | + |
| Campylobacter hyointenstinalis | + | + | V | + | V | − | − | + | + | S | S | + | − |
NA – Nalidixic acid, TSI – triple sugar iron agar R – resistant, S – sensitive, V – variable, W – weak, U – unknown, (V) – a few strains +
Source: modified from Corry et al. (1995); Corry and Atabay (2011)
PHE protocols of Campylobacter spp. isolation from food samples involve enrichment of homogenate (25 g) in Bolton broth (10−1 dilution) and incubations for 5 hours at 37°C and 44 hours at 41.5°C (PHE 2014). Then, enrichment cultures are streaked on mCCDA media plates and microaerobically incubated (44 hours, 41.5°C) (Fig. 11, Table I). The microaerobic growth (41.5°C) of presumptive Campylobacter spp. colonies is compared to the aerobic growth (25°C) on blood agar plates to confirm their identity (PHE 2014). The procedure involves the examination of five suspected colonies from each mCCDA plate. PHE protocol also recommends other confirmatory steps such as cell motility’s microscopic examination and Oxidase test. Furthermore, PHE also recommends optional confirmation through PCR assay and latex test kits [Campylobacter Latex Kit (LIOFILCHEM® S.r.l., Italy), Oxoid™ DrySpot™ Campylobacter Test Kit (Thermo Fisher Scientific, Inc., USA), and Campylobacter Confirm Latex kit (Bio-Rad Laboratories, Inc., USA)] (PHE 2014).
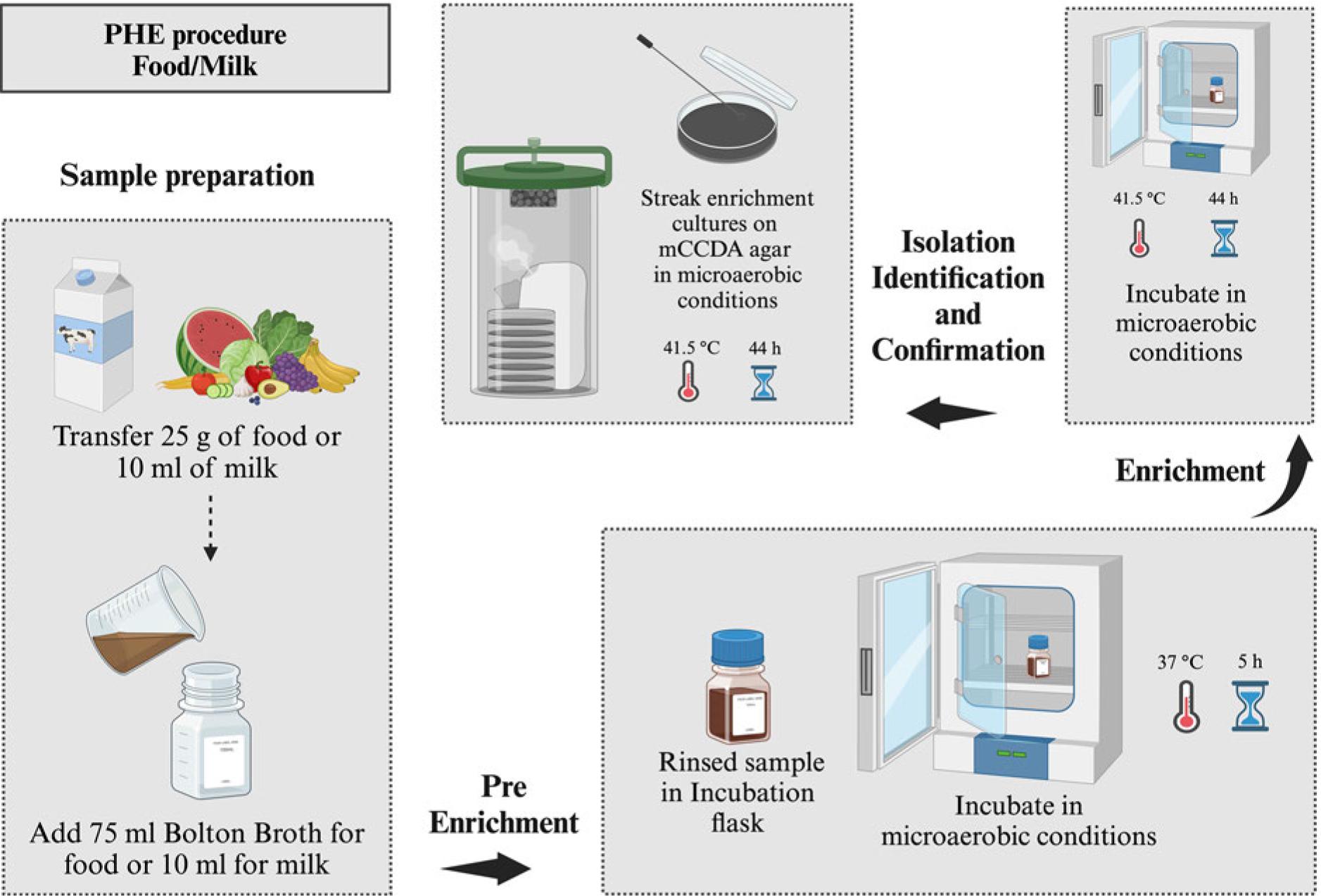
Public Health England procedure for the isolation of Campylobacter spp. from all types of food and milk, microaerobic conditions (N2: 85%, CO2: 10%, and O2: 5%).
| Media | Basal medium | Antioxygen system | Cephalosporin | Trimethoprime | Polymyxin B/colistin (C) | Rifampicin | Novobiocin | Vancomycin | 5-Fluorouracil or Na-deoxycholate (D) | Bactiracin | Antifungals |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Broth media | |||||||||||
| Bolton | A | 5% LH, 0.1 g HE, 0.5 g P, 0.5 g B | 20 czone | 20 | − | − | − | 20 | − | − | 50 CY |
| Preston | NB | 5% LH/FPB | − | 10 | 5000 | 10 | − | − | − | − | 100 CY |
| Exter | NB | 5% LH, 0.5 F, 0.2 B, 0.2 P | 15 czone | 10 | 4 g (C) | 10 | − | 10 | − | − | − |
| Rosef and Kapperud | NB | − | − | 5 | 2500 | − | − | 10 | − | − | − |
| Doyle and Roman | BB | 7% LH | − | 5 | 20000 | − | − | 15 | − | − | 50 CY |
| Agar media | |||||||||||
| Skirrow | BAB | 7% LH | − | 5 | 2500 | − | − | 10 | − | − | − |
| Campy-BAP | BA | 10% S | 15 cthin | 5 | 2500 | − | − | 10 | − | − | 2 AM |
| Butzler | TB | 10% S | 15 cthin | − | 10000 (C) | 5 | 25000 | 50 CY | |||
| CCD | NB | 4 g C, 0.25 F, 0.25 P | 10 czolin | − | − | − | − | − | 1000 (D) | − | − |
| mCCD | NB | 4 g C, 0.25 F, 0.25 P | 32 czone | − | − | − | − | − | 1000 (D) | − | 10 AM |
| Karmali | CA | 4 g C, 0.32 g HT, 0.1 P | 32 czone | − | − | − | − | 20 | − | − | − |
CCD – charcoal cefazolin deoxycholate, mCCD – modified charcoal cefoperazone deoxycholate, Campy–BAP – Campy Brucella agar, NB – nutrient broth, BB – Brucella broth base, BAB – blood agar base, BA – Brucella agar base, TB – thioglycolate broth base, CA – Columbia agar base, LH – lysed horse blood, HE – haemin, B – sodium metabisulfate, P – sodium pyruvate, F – ferrous sulfate, FPB – ferrous sulfate/sodium pyruvate/sodium metabisulfate, S – sheep blood, C – charcoal, HT – hematin, czone – cefoperazone, cthin – cephalothin, czolin – cefazolin, CY – cycloheximide – actidione, AM – amphotericin B, (C) – colistin, (D) – Na deoxycholate, A – 10 g meat peptone, 5 g lactoalbumin hydrolysates, 5 g yeast extract 5 g NaCl, 0.6 g NaCO3
Source: modified from Corry et al. (1995); Corry and Atabay (2011)
Foodborne Campylobacter spp. is conventionally recovered by adopting culturing and isolation methodologies (Corry et al. 1995; Silva et al. 2011; Fung et al. 2018). Selective enrichment is the initial step in conventional Campylobacter spp. recovery methods, which is followed by selective plating for isolation and confirmatory tests (immunological, molecular, and biochemical). The enrichment broths could revive stressed and inhibitor-exposed bacteria in the tested matrix and facilitate the recovery of isolated bacteria even at low concentrations (Corry et al. 1995; FDA 2001; Corry and Atabay 2011; ISO 2017; USDA FSIS 2020). Enrichment broths differ in nutrient composition, incubation time, oxygen-degradation, environment and temperature requirements, and antimicrobial substances are added to restrain the growth of other competing microorganisms (Baylis et al. 2000; Oyarzabal et al. 2007) (Table III). Numerous enrichment broth formulations have been formulated for Campylobacter spp. isolation including Exeter broth, Bolton broth (BB), Modified CCD broth, Preston broth (PB), Doyle and Roman broth, and Rosef and Kapperude Campylobacter enrichment broth (Doyle and Roman 1982; Rosef and Kapperud, 1983; Bolton et al. 1984a; Baylis et al. 2000; McBride et al. 2002). Bolton broth and Preston formula are crucial for primary selective enrichment and are recommended due to satisfactory output, particularly in low bacterial count and stressed bacteria (Bolton et al. 1983; Repérant et al. 2016).
The enrichment stage enhances microfloral growth in the target samples. Therefore, selective substances should be used to optimize Campylobacter spp. growth conditions for better recovery. There is no specific standard method for recovering Campylobacter species, particularly non-thermotolerant species (Perry 2017). Antimicrobials-supplemented basal medium (nutrient broth or Brucella broth) is the main ingredient of enrichment broths (Bolton and Robertson 1982; Corry et al. 1995). Enrichment broths were initially supplemented with lysed sheep or horse blood to reduce oxidative toxins’ damage (Bonnet et al. 2019). However, comparatively high blood cost and inessential isolation of Campylobacter spp., from poultry meat reduced their applications (Karmali et al. 1986; Bolton et al. 2002; Line 2006; Liu et al. 2009). Blood-free formulas are more convenient and can also be integrated with molecular techniques for rapid pathogen detection and identification. Campylobacter spp. isolation does not require a rich basal medium. USDA Food Safety and Inspection Service also uses blood-free Bolton broth, the best enrichment alternative (Baylis et al. 2000; USDA FSIS 2011). The buffered peptone water is quite similar to Bolton broth’s basal component and is equally effective for Campylobacter spp. isolation from broiler meat (Oyarzabal et al. 2007).
Bolton broth is recommended for the enrichment of all sample types, particularly the US FDA recommends it for the recovery of Campylobacters spp. from various types of samples (environmental, food, and clinical). ISO also recommends Bolton broth for samples’ enrichment with lower Campylobacter spp. and background bacterial count (FDA 2001; ISO 2017). Bolton broth contains different nutrients, including yeast and peptone extract, sodium pyruvate, alpha-ketoglutaric acid, hemin, and sodium metabisulphite. Hemin helps in overcoming trimethoprim antagonism of yeast extracts (Table III). The addition of sodium metabisulphite and sodium pyruvate allows aerobic incubation, whereas sodium carbonate provides carbon dioxide for bacterial growth (Post 1995). The medium contains antibiotics (cycloheximide, cefoperazone, trimethoprim, and vancomycin) and lysed horse blood. Antibiotics restrict the growth of non-specific contaminating microorganisms (Post 1995; Line et al. 2008; Yoo et al. 2014). Specific substrates in Bolton broth limit trimethoprim antagonism, whereas hemin, ferrous sulfate/sodium metabisulfite/sodium pyruvate (FBP) mixture, and blood enhance oxygen quenching (Table III). Vancomycin in Bolton broth suppresses Gram-positive cocci but has lower efficacy against Campylobacters spp. as compared to rifampicin in Exeter and Preston broths (Donnison 2003). However, Bolton broth is preferred for Campylobacter spp. isolation from poultry samples (Baylis et al. 2000). Bolton broth remains unable to detect certain Campylobacter species (C. coli and C. jejuni) in vegetables and chicken (Jasson et al. 2009; Habib et al. 2011). Antibiotics in Bolton broth enhance its selectivity (Moran et al. 2011). However, cefoperazone might reduce the selectivity in mCCDA (modified charcoal cefoperazone deoxycholate) agar and Bolton broth, which could be due to the absence of rifampicin and polymixin (Jasson et al. 2009) (Table III). However, it is still helpful for samples with lower numbers of sublethally damaged or stressed Campylobacter spp. and samples containing lower numbers of non-target organisms (ISO 2017). Several modifications of incubation temperatures and selective agents have been suggested for accurate and improved Campylobacter spp. detection (Baylis et al. 2000).
Preston broth is another commonly used enrichment broth for the isolation of Campylobacter spp. from various complex samples, including environmental (FDA 2001; ISO 2017), turbid surface water, and food specimens (Humphrey et al. 1995; Abulreesh et al. 2005). Preston medium is nutrient broth comprised of lysed horse blood and antibiotics (cycloheximide, rifampicin, polymyxin B, and trimethoprim). In contrast, it does not contain yeast extract (trimethoprim antagonist) (ISO 1995) (Table III). Rifampicin is highly effective against Gram-positive bacteria. The culture media is incubated at 42°C under a microaerobic atmosphere (Post 1995). The presence of cycloheximide/amphotericin B, polymyxin B, trimethoprim, and rifampicin significantly enhances the selectivity of Preston broth (Table III). Polymyxin B inhibits the growth of extended-spectrum beta-lactamase (ESBL) bacteria as it possesses high activity against Gram-negative bacteria (Bolton and Robertson 1982). Therefore, samples with high background flora (ESBL bacteria) are preferably grown in Preston broth (ISO 2017). Preston broth has demonstrated high selectivity against non-target flora during Campylobacter spp. enrichment (Bolton and Robertson 1982; Uyttendaele and Debevere 1996; Jasson et al. 2009; Habib et al. 2011; Ugarte-Ruiz et al. 2012). ISO also recommends Preston broth for Campylobacter spp. isolation from samples (poultry and milk) with high background bacteria (ISO 2017). Ugarte-Ruiz et al. (2012) have compared different enrichment methods and noted better efficacy of Preston broth than Bolton broth, which allowed the growth of some Escherichia coli strains that could hinder Campylobacter spp. growth to produce false-negative outcomes. Contrarily, some studies have depicted inhibited growth of Campylobacter strains (C. coli) in Preston broth, leading to false negative results (Goossens et al. 1986; Baylis et al. 2000; Paulsen et al. 2005).
Exeter broth is routinely used in various laboratories to analyze water and food samples. It is also a primary enrichment medium for freshwater microbiological investigations (McBride et al. 2002). Exeter selective broth’s formulation is based on a lysed horse blood (5%) supplemented nutrient broth (Humphrey et al. 1995; de Boer and Humphrey 1991) (Table III). Later on, the formula was modified (Humphrey 1986), and oxygen-quenching agents ferrous sulfate/sodium metabisulfite/sodium pyruvate (FBP) mixture of Boltonv et al. (1984a, b) were added, which allowed aerobic incubation of Exeter broth. Exeter broth also contained different antibiotics, including cefoperazone (against Pseudomonas spp., and Enterobacterales), rifampicin, polymyxin B, amphotericin (against yeasts and molds), and trimethoprim (Humphrey et al. 1995). Modified charcoal cefoperazone deoxycholate broth (mCCD) is another blood-free selective enrichment broth that was modified from the original charcoal cefazolin deoxycholate (CCD) medium (Bolton et al. 1984b). The mCCD broth was mainly comprised of nutrient broth, cefoperazone, casein hydrolysates, bacteriological charcoal, and FBP supplements (sodium pyruvate, sodium deoxycholate, and ferrous sulfate) (Table III). The mCCD broth contains different Campylobacter spp. growth-promoting components and helps in the direct isolation of Campylobacter spp. from animal and human feces (Hutchinson and Bolton 1984; Bolton et al. 1984b). Charcoal, deoxycholate, and cefoperazone combination inhibits bacterial growth (commensal flora and common contaminants) in food and clinical samples.
Rosef and Kapperud Campylobacter enrichment broth (Rosef and Kapperud 1983) contains sodium chloride, peptone, and antimicrobials (polymyxin B, vancomycin, and trimethoprim). Cysteine hydrochloride and sodium succinate-supplemented Brucella broth served as the basal medium in Doyle and Roman enrichment broth (DREB) (Doyle and Roman 1982) (Table III). Antibiotics (polymyxin B, vancomycin, cycloheximide, and trimethoprim) and lysed horse blood were also added for better enrichment efficiency. Doyle and Roman (1982) used Brucella broth as a basal medium and altered its selectivity through significantly increased concentrations of cycloheximide and polymyxin (Table III), facilitating the selective recovery of lower Campylobacter spp. numbers in food samples. Cysteine hydrochloride and Succinate were also added, whereas lysed horse blood (7%) acted as an oxygen-quenching system. The medium was able to analyze raw milk and hamburger (0.1 to 4.0 cells/gram) but remained ineffective for poultry samples, which might be due to the diverse types and amounts of flora in these samples. Therefore, the DREB medium was further modified to rapidly enrich C. jejuni from raw chicken carcass samples (Rothenberg et al. 1984). Doyle and Roman enrichment broth was established as the most suitable for detecting lower C. jejuni levels in chicken meat samples after 12 months of storage at -18°C. Rothenberg et al. (1984) performed a comparison study of Park and Stankiewiez enrichment broth, Doyle and Roman enrichment broth, and a newly developed enrichment broth for C. jejuni isolation from raw chicken and revealed the highest selectivity potential of Doyle and Roman enrichment broth (Khan et al. 2018b).
All the Campylobacter spp. isolating plating media used for food samples are either direct compositions or modified forms of clinical media that were developed for Campylobacter spp. isolation from fecal and clinical samples. Different types of plating media are available for Campylobacter spp. isolation with varying selectivity. These media are divided into two groups such as bloodcontaining solid media known as Campylobacter blood agar plates [Skirrow agar, Campy Brucella agar (Campy-BAP), Butzler agar, and Preston agar], and charcoalbased solid plating media [Karmali agar, and mCCD agar] (Corry et al. 1995; Corry and Atabay 2011). Despite poor productivity and sensitivity in food samples, Karmali agar and mCCDA are the best media for Campylobacter spp. isolation as colonies are easily recognizable in both media (Jacobs-Reitsma and De Boer 2001; Chon et al. 2011; El Baaboua et al. 2022).
Charcoal compounds and blood can reduce the toxic oxygen derivatives to generate a microaerobic environment for Campylobacters growth. Agar plates were also developed without charcoal or blood, demonstrating considerably lower efficacy than charcoal or blood-added broths (Bolton and Robertson 1982; Hutchinson and Bolton 1984) (Table III). The resistance of thermophilic Campylobacter spp. to various antibiotic combinations in media determines its efficacy. Antibiotics such as polymyxin, vancomycin, rifampicin, trimethoprim, cefoperazone, nystatin, cephalothin, cycloheximide, and colistin inhibit background microbiota growth in samples and allow the isolation of slow-growing Campylobacter spp. (Martin et al. 2002; Oyarzabal et al. 2005; Vandenberg et al. 2005; Williams and Oyarzabal 2012; Zhang and Sahin 2013) (Table III). The capability of contaminating-flora inhibition differentiates between various media. All the selective agents facilitate the growth of C. coli and C. jejuni. To date, no medium can inhibit C. coli while allowing the growth of C. jejuni or vice versa. Other Campylobacter species (C. hyointestinalis, C. lari, C. fetus, C. upsaliensis, and C. helveticus) also grow on most media to some extent, particularly at a less selective temperature of 37°C.
Skirrow’s selective agar medium was the first widely recommended for C. coli and C. jejuni isolation from human feces (Skirrow 1977). It replaced the complicated method of selective filtration through 0.65 μm pore-size membranes. Skirrow’s Campylobacter selective agar contains peptone, lysed horse blood (7%), and antibiotics (trimethoprim, vancomycin, and polymyxin B) (Skirrow 1977; Corry et al. 1995) (Table III). The addition of vancomycin (inhibits Gram-positives), trimethoprim (broad-spectrum antibiotic), and polymyxin B (antifungal) mixture enhances its selectivity. The addition of lysed horse blood neutralizes trimethoprim antagonists of basal medium, leading to promoted growth of polymyxins-resistant Gram-negative Proteus spp. The incubation temperature of 42°C also contributes to the medium’s selectivity. Thus, only thermophilic Campylobacter spp. can grow in Skirrow’s medium, whereas the growth of non-thermophilic strains (C. fetus subsp. fetus) is restricted at 42°C. Skirrow’s medium is sometimes used for Campylobacter spp. detection in food samples but remains ineffective for many other types of samples (Post 1995; Corry et al. 1995; Corry and Atabay 2011).
Campylobacter-selective agars contain different antibiotic combinations. Blood-containing Campy Brucella agar plate, also known as Campy-BAP, has been widely used (Blaser et al. 1979). Campy-BAP is a Brucella base agar that contains five antimicrobial agents (cephalothin, vancomycin, amphotericin B, polymyxin B, and trimethoprim), and is supplemented with sheep blood (10%) (Line 2001) (Table III). Antibiotics such as polymyxin B, cephalothin, and colistin might inhibit the growth of C. coli, C. jejuni, and C. fetus subsp. fetus (Ng et al. 1985; Goossens et al. 1986; Ng et al. 1988). Beuchat (1985) compared various enrichment techniques and direct isolation media to enumerate five C. jejuni strains in stored/refrigerated chicken meat. Campy-BAP agar and blood-free Campylobacter medium exhibited higher C. jejuni strains detection capability than Doyle and Roman enrichment broth and modified Butzler agar (Khan et al. 2018b). Endtz et al. (1991) investigated five types of selective media, including Campy-BAP and charcoal cefazolin deoxycholate agar (CCDA). They noted a better recovery rate with the CCDA medium (83%) as compared to the Campy-BAP medium (75%). CCDA medium also effectively suppressed normal enteric flora contamination. Campylobacter spp. colonies, particularly C. coli, appeared atypical on the Campy-BAP medium. The strains mainly produced homogeneous, discrete, and grey colonies, which were difficult to differentiate from coliform colonies in several cases. Campylobacter spp. colonies exhibited transparent and moist growth on other media. The morphology of colonies on Campy-BAP medium complicated the Campylobacter spp. identification process (Bolton et al. 1983). Preston Campylobacter selective agar was specifically developed for Campylobacter spp. isolation from diverse specimens (environmental, human, and animal) (Post 1995). Bolton and Robertson (1982) prepared Preston medium by dissolving nutrient broth in New Zealand agar and adding horse blood (saponin-lysed) and antibiotics (trimethoprim, polymyxin, actidione, and rifampicin). Preston medium demonstrated high Campylobacter spp. isolation rate from all tested samples and remained the most selective medium compared to other media (Bolton and Robertson 1982).
Campylobacter agar (Butzler’s) is used to isolate Campylobacter species selectively from different specimens, including clinical samples. Lauwers et al. (1978) reported the first selective formulation containing sheep blood agar and five antimicrobials (cephalothin, novobiocin, colistin, actidione, and bacitracin) where bacitracin and cephazolin inhibited the Gram-positive bacteria, and colistin and novobiocin inhibited the Gram-negative enteric flora. Further addition of cycloheximide inhibited the growth of common clinical mycotic contaminants. This medium was developed as an alternative to filtration and culturing on an elective blood-thioglycollate agar medium employed to examine human blood and fecal samples for vibrios (Dekeyser et al. 1972). Cephalothin, in addition to the original formula (bacitracin, cycloheximide, novobiocin, and colistin), significantly enhanced its selectivity with the filtration method. Sheep blood agar serves as the basal medium in Butzler’s agar (Table III). Initial incubation is carried out at 42°C, and the temperature gradually decrease to grow C. jejuni but hinders the growth of C. etus subspecies intestinalis (Goossens et al. 1983).
Modified charcoal cefoperazone deoxycholate agar (mCCDA) is enlisted in international standard protocols and is widely used worldwide, where it is recommended as the plating media of choice for the detection and enumeration of Campylobacter spp. (FDA 2001; PHLS 2002; PHE 2014; ISO 2017). It generates satisfactory results and is recommended for selective plating. mCCDA medium is based on the Bolton et al. (1984b) formula and is comprised of New Zealand agar, nutrient broth, bacteriological charcoal, sodium pyruvate, casein hydrolysates, ferrous sulfate, and sodium deoxycholate (Table III). The selectivity of this media was further enhanced by replacing cephazolin with cefoperazone (Hutchinson and Bolton 1984). Initially, its development was aimed at thermotolerant Campylobacter spp. isolation from human fecal samples but then emerged as a specified standard medium for Campylobacter spp. isolation from food samples. The blood was replaced with sodium pyruvate, charcoal, and ferrous sulfate in the mCCDA medium, increasing the aerotolerance and growth of Campylobacter spp. Casein hydrolysate in this medium promotes the growth of C. lari environmental strains, whereas sodium deoxycholate and cefoperazone provide the required selectivity (Bolton et al. 1988).
Campylobacter selective mCCDA agar is a widely used blood-free plating medium (Bolton and Robertson 1982; Hutchinson and Bolton 1984; Piersimoni et al. 1995). Thus, it helps to avoid the disadvantages of blood, such as easy contamination, short life, and expensive nature (Piersimoni et al. 1995). The stickiness of Campylobacter spp. colonies to the plate surface in some cases is the only limitation that complicates harvesting (Stern et al. 1992). mCCDA and Skirrow media containing different antimicrobials have been used for culturing Campylobacter spp. where cefoperazone in mCCDA media proved a more effective selective agent and efficiently suppressed the enteric flora (Goossens et al. 1986; Gun-Munro et al. 1987) (Table III). The higher efficacy of broad-spectrum cefoperazone (cephalosporin) has been established against Enterobacteriaceae family members and pseudomonads (Gun-Munro et al. 1987).
Karmali is a charcoal-based, blood-free selective medium. It is comprised of Columbia agar base, hematin, activated charcoal, sodium pyruvate, cycloheximide, cefoperazone, and vancomycin (Karmali et al. 1986). Karmali medium was developed to overcome mCCDA selective agar-associated limitations (Bolton et al. 1984b). Karmali et al. (1986) demonstrated significantly higher selectivity of Karmali agar and better Campylobacter spp. isolation rate from fecal samples as compared to Skirrow’s medium. Similar to blood, charcoal also acts as a quenching agent for enhanced aerotolerance against oxygen derivatives’ toxicity (Bolton et al. 1984a; Bolton and Coates 1983). Thus, charcoal-based agar is a better alternative for blood-containing agar in developing countries, which face erratic availability of sterile blood.
Karmali agar medium contains sodium pyruvate in the selective supplement, whereas it is found in the basal medium of other blood-free Campylobacter spp. isolation media (mCCDA). Ferrous sulphate in mCCDA media is replaced with hemin in the Karmali medium. Vancomycin in the Karmali medium replaces the deoxycholate of mCCDA media (Table III) and strongly inhibits the growth of Gram-positive microorganisms. Vancomycin is particularly effective against enterococci and thus eliminates bile salts’ inherent variability. Cefoperazone in this medium efficiently suppresses the growth of Pseudomonas spp. whereas cycloheximide more effectively inhibits yeasts than amphotericin B (Post 1995; Corry et al. 1995). The three antibioticselective agents (cycloheximide, cefoperazone, and vancomycin) in the Karmali medium (Table III) efficiently restrict the growth of Gram-negative and Grampositive bacteria, and yeasts. During the development of this media, the efficacy of these antibiotic-selective agents was individually assessed (Post 1995; Corry et al. 1995). Karmali medium has been proven more selective than Skirrow medium. Some C. coli strains are cephalosporins-susceptible and the Skirrow medium performs better for isolating these strains than the Karmali medium. During a study, combining Skirrow and Karmali mediums produced near-optimal results for thermotolerant Campylobacter spp. isolation from fecal samples (Post 1995). Gun-Munro et al. (1987) revealed that charcoal and cefoperazone-containing Campylobacter spp. isolation media (mCCDA) generated better outcomes than earlier formulations.
Adding chromogenic agar media in isolation protocols enhanced Campylobacter species identification capability through distinctive colony color. Synthetic chromogenic enzyme substrates in chromogenic media aid in their utility as both differential and selective media to identify the target isolate through their enzyme activity (Manafi 2000; Perry 2017). A few commercial chromogenic agar plates are available in Latin America, the USA, and Europe. These plates are utilized to isolate Campylobacter spp. from meat, carcass rinse, environmental samples, and poultry meat. Campylobacter spp. isolating (food samples) sensitivity of chromogenic agars is similar to traditional plates (Ahmed et al. 2012; Seliwiorstow et al. 2014).
CHROMagar™ Campylobacter (CHROMagar™, France), CampyFood® agar (bioMérieux, France), R&F® Campylobacter media (R&F Products, USA), and Brilliance™ CampyCount Agar (Thermo Scientific™, Thermo Fisher Scientific, Inc., USA) are Campylobacter chromogenic media, facilitating visual recognition of Campylobacter spp. colonies without requiring subsequent culturing and confirmation tests. Thus, these media decrease the analysis’s cost and time (Al-Wasify 2013; Gharst et al. 2013a; 2013b; Josefsen et al. 2015). Campy-Food® agar was the first commercial chromogenic-like agar that matched CCDA capabilities to isolate and enumerate Campylobacter spp. (C. coli and C. jejuni) from poultry samples. CampyFood® agar plating medium was recommended for its high selectivity and better performance than the specificity (68%) and sensitivity (100%) of mCCDA (Habib et al. 2011). Moreover, it eliminates or minimizes the contamination of swarming and spreading colonies in tested samples (Ahmed et al. 2012). CampyFood® agar plates are easy to handle and produce results comparable to those of other media (Habib et al. 2011; Ugarte-Ruiz et al. 2012). However, some other bacterial species might also grow on the plates, leading to Campylobacter spp. colonies’ overestimation. Therefore, the CampyFood® medium is not entirely different for Campylobacter spp. isolation (Habib et al. 2008; 2011; Ahmed et al. 2012). An investigation in Chile reported a higher CampyFood® medium-based Campylobacter spp. isolation rate from chicken meat (83%) than mCCDA (67%) (Fernández-Riquelme 2011).
A selective chromogenic medium, Brilliance™ Campy Count agar, was developed explicitly for Campylobacter spp. (C. coli and C. jejuni) enumeration from poultry samples. Brilliance™ CampyCount agar is comprised of an amino acid and salt mix that allows accurate, clear, and specific C. coli and C. jejuni enumeration on poultry carcass samples (Atlas and Snyder 2013). Brilliance™ CampyCount medium was carefully developed to achieve C. coli and C. jejuni growth while inhibiting the growth of non-target microorganisms. This medium indicates the colonies through a color change to dark red. Thus, all C. jejuni/coli colonies become readily identifiable within 48 hours on a transparent BCC medium (Habib et al. 2011). Habib et al. (2011) indicated that Brilliance™ CampyCount and CampyFood® media efficiencies were comparable to mCCDA for Campylobacters spp. enumeration in naturally contaminated chicken meat. Brilliance™ CampyCount agar can be a potential alternative to mCCDA, but further investigation is required to enhance its selectivity for improved accuracy of Campylobacter spp. enumeration and minimum background microflora (Ahmed et al. 2012).
CHROMagar™ Campylobacter is a selective chromogenic medium that is widely used for presumptive identification, direct qualitative detection, and differentiation of main thermo-tolerant Campylobacter spp. (C. lari, C. jejuni, and C. coli) from environmental and food samples by following the ISO 10272-1:2017 (2017) method. CHROMagar™ Campylobacter comprises of a chromogenic substrate, agar, yeast extract, peptones, Sodium chloride, and a selective mix (Sylte et al. 2018). CHROMagar™ Campylobacter is also a blood-free transparent agar-like CLA-S medium that helps visualize and enumerate Campylobacter spp. colony forming unit (CFU) by producing purple colonies (Sylte et al. 2018). R&F® Campylobacter chromogenic agar plating medium targets the C-2 esterase enzyme of C. coli and C. jejuni. C. coli and C. jejuni are C-2 esterase positive, whereas other microorganisms remain negative to this enzyme. R&F® Campylobacter chromogenic medium has enhanced sensitivity, and visual identification can easily distinguish the colonies. All current Campylobacter spp. isolation broth, media, and plates are modifications of media, which were developed almost three decades ago when achieving microaerobic conditions in the laboratories was challenging (Oyarzabal and Fernández 2016).
Culture-based techniques are a standard cultivation method for bacterial detection and enumeration of food and water (Talaro and Talaro 2002). However, some foodborne and waterborne enteric bacteria, including Campylobacter spp. could enter a viable but non-culturable (VBNC) state that can somehow lose their growing capability on culture media (Oliver 2000; Santos et al. 2023). Despite non-culturability, VBNC cells are not considered dead due to different dissimilarities. A damaged membrane is the main feature of dead cells that cannot retain plasmids and chromosomal DNA, whereas the membrane of VBNC cells remains intact with undamaged DNA and plasmids (Cook and Bolster 2007; Oliver 2010). Dead cells become inactive metabolically, but VBNC cells remain metabolically active and perform respiration (Lleò et al. 2000; Besnard et al. 2002). Gene expressions stop in dead cells and transcription continues in VBNC cells followed by the production of mRNA (Lleò et al. 2000). VBNC cells, in contrast to dead cells, continue to uptake and incorporate amino acids into proteins (Lleó et al. 1998). VBNC bacterial cells retain their virulence and cause infection upon entry into hosts. Thus, they are a serious concern for public health, particularly about water and foodborne pathogens (Fakruddin et al. 2013; Li et al. 2014; Ramamurthy et al. 2014). Several studies have reported VBNC C. jejuni colonization in the rat guts, suckling mice, fertilized chicken eggs, and chicks (1-week-old) (Jones et al. 1991; Saha et al. 1991; Stern et al. 1994; Talibart et al. 2000). Baffone et al. (2006) successfully used artificial seawater to resuscitate the C. jejuni VBNC cells after 142 days by passing through the mouse intestine. Thus, VBNC C. jejuni could retain the virulence and infectivity. However, the infective capability of environmental VBNC cells without resuscitation remains unclear (McDougald et al. 1998). An in vitro study has also reported the invasion of Caco-2 human intestinal epithelial cells by VBNC C. jejuni (Chaisowwong et al. 2012).
Disease diagnosis and etiological agents’ identification in clinical, water, and food samples are still highly dependent on culture-based techniques. The inability to culture microorganisms could be a major limitation in disease diagnosis and treatments. This situation complicates pathogen detection in environmental, water, and food samples. Thus, potentially hazardous contaminations could remain undetected, and water and foodborne VBNC bacteria could seriously threaten public health (Li et al. 2014; Pan and Ren, 2023). VBNC state in food and water could be generally attributed to low-grade or aseptic infections and could be mistakenly linked to viruses if no bacteria are detected (Fakruddin et al. 2013).
Generally, the enrichment step resuscitates VBNC and damaged cells. Therefore, enrichment culture of water and food bacteria in selective/basal broth notably enhances the retrieval of experimentally injured Campylobacter spp. (Humphrey 1989). An enrichment regime involving incubation (4 hours, 37°C) in broth [lysed horse blood (5%), sodium metabisulphite (0.02%), sodium pyruvate (0.02%), and ferrous sulfate (0.05%)] followed by another incubation (44 hours, 43°C) significantly improved damaged Campylobacter spp. cells’ recovery from river water samples (Humphrey and Muscat 1989). It might have facilitated the repair of injured cells before exposure to high temperatures (Humphrey 1986; Humphrey and Muscat 1989).
Propidium monoazide (PMA)-viability-qPCR approach could successfully detect the natural occurrence of VBNC Campylobacter spp. cells in environmental samples (chicken manure and barn). The study further demonstrated the Campylobacter spp. viability in water and soil up to 63 and 28 days, respectively (Reichelt et al. 2023). PMA-qPCR technique also efficiently detected laboratory-induced C. jejuni VBNC cells in UHT and pasteurized milk (Zhang and Lu 2023). It can also quantify VBNC Campylobacter spp. to provide insights into the unculturable Campylobacter spp. prevalence in agri-food productions and environment (Lv et al. 2019). This method offers an effective solution to overcome the limitations of traditional culture-based methods. However, it requires costly apparatus and highly trained personnel during the pre-treatment of samples for a successful VBNC Campylobacter spp. DNA isolation.
PCR-based direct Campylobacter spp. cell detection in environmental and food samples is a time-effective approach as compared to culture-based identification and confirmation. Different PCR protocols with diverse primers have been developed for Campylobacter spp. detection in wastewater and water samples (Birkenhead et al. 1993; Kirk and Rowe 1994; Waage et al. 1999; Savill et al. 2001; Alexandrino et al. 2004). Most PCR-based studies analyzed the Campylobacter spp. absence or presence in samples whereas some studies obtained quantitative results by employing real-time PCR (Yang et al. 2003). These protocols facilitated the recovery from Campylobacter spp.-seeded cultures. Direct PCR assay could efficiently detect the natural occurrence of Campylobacter spp. in polluted drinking water without an enrichment step (Jackson et al. 1996; Moore et al. 2001). Yang et al. (2003) also employed a direct PCR approach to successfully detect C. jejuni in naturally contaminated water without prior enrichment of samples. PCR-based direct Campylobacter spp. detection in clean drinking water might be feasible. However, it could generate false negative results in samples containing high levels of background bacteria (Abulreesh et al. 2014).
In contrast to contaminated drinking water samples, Campylobacter spp. presence in milk, poultry, and environmental murky water samples remains comparatively low with high levels of background microbiota and PCR inhibitors. Therefore, the enrichment step becomes mandatory before PCR detection (Abulreesh et al. 2014; Ricke et al. 2019). Similarly, multiple studies have performed enrichment steps before PCR detection of Campylobacter spp. in various types of samples such as river and spiked estuarine water samples (Hernandez et al. 1995; Waage et al. 1999), spiked and naturally polluted sewage samples (Koenraad et al. 1995; Waage et al. 1999), spiked and natural food contamination samples (Giesendorf et al. 1992; Denis et al. 2001; Sails et al. 2003), samples of natural poultry and human fecal contaminations (Rasmussen et al. 1996; Denis et al. 1999; Vanniasinkam et al. 1999), spiked chicken rinse water (Ng et al. 1997; Josefsen et al. 2004), and murky pond water (Abulreesh et al. 2014). Generally, enrichment incubation increases the target cell population for better PCR detection.
Direct PCR, multiplex PCR, and qPCR can also amplify the DNA of dead cells and naked DNA fragments in water and food samples without the enrichment step (Mandrell and Wachtel 1999; Ricke et al. 2019). The presence of dead Campylobacter spp. cells in water samples depict contamination but are no longer harmful to public health (Theron and Cloete 2004). Therefore, the induction of an enrichment step before PCR assay enhances the detection of viable cells. Selective enrichment followed by PCR assay has emerged as a standard method for Campylobacter spp. detection in environmental samples (Josefsen et al. 2004; Abulreesh et al. 2014). The FISH (fluorescence in situ hybridization) method differentiates DNA fragments and whole cells. A fluorescent Campylobacter spp.-specific oligonucleotide probe is used to label the whole cells, followed by epifluorescence microscopy. Lehtola et al. (2005) detected C. coli after membrane filtration of spiked tap water and noted hybridized cells with different fluorescence brightness, which helped separate senescent and actively growing C. coli cells. Immunebased assay could be another alternative to traditional culture-based methods. However, immune-assay kits are yet to be validated for Campylobacter spp. detection in food (poultry) samples, possibly due to matrix-induced sensitivity loss (Ricke et al. 2019).
Despite extensive developments and research of alternatives for precise and rapid Campylobacter spp. detection, identification, and quantification in samples (environmental, food, and clinical), and culture-based techniques are still the gold standard. Standard organizations recommend immune-based and molecular approaches for the identification and precise confirmation of presumptive Campylobacter spp. colonies along with conventional confirmatory tests (FDA 2001; PHE 2014; ISO 2017).
Molecular and culture methods of Campylobacter spp. detection present particular advantages and disadvantages over each other, which are discussed below:
Traditional culturing grows only viable Campylobacter spp. cells via selective media and thus achieves high specificity by restricting the interference of other microorganisms. This approach detects only live pathogens to indicate a current risk of the infection. Molecular techniques (PCR and qPCR) can efficiently detect Campylobacter-specific DNA in complex samples. However, these methods can detect dead and live cells, including nonviable cells’ residual DNA, and potentially yield false positives related to infection risk.
The sample’s initial bacterial load and competing flora could alleviate the sensitivity of culture-based methods. The pathogenic VBNC cells of Campylobacter spp. could remain undetected in selective media, which results in the underreporting of infections. On the other hand, Campylobacter spp. DNA can be detected by more sensitive molecular approaches even in background flora-contaminated samples. The sensitivity of PMA-qPCR is even better as it could exclude the dead cells in the sample.
The enrichment, selective plating, and incubation steps lengthen the culture-based methods and take several days to produce results. This limits routine pathogen monitoring, where quick outputs are preferred. Conversely, PCR produces results within hours to help in real-time pathogen monitoring and timely response during outbreaks.
Microbial culturing can be performed without costly equipment, so these methods are preferred in less developed laboratories. However, laborious culture-based approaches are time-consuming and require more manual handling. Conversely, expensive reagents and equipment (qPCR and PCR thermocyclers) are required for rapid molecular techniques. Moreover, highly trained personnel are required to perform these procedures, which limits their utility in settings with fewer resources.
The cultured colonies facilitate further analysis through genotyping, biochemical tests, and antibiotic susceptibility testing, a prerequisite for outbreak tracking and epidemiological studies. Molecular approaches (qPCR, DNA sequencing, and multiplex PCR) facilitate precise and rapid identification of pathogens, genetic analysis, and epidemiological tracking. However, further analyses are restricted in this case, as these methods cannot provide live bacterial culture.
Culture-based techniques can be applied to diverse types of water and food samples. However, complex food matrices can affect the detection process. Similarly, environmental and food samples can also alleviate the detection efficiency of molecular approaches. Therefore, an increase in bacterial cell numbers via pre-enrichment is often required before analysis.
ISO and FDA have standardized culture-based detection protocols as the gold standard for water and food safety testing. Conversely, rapidly emerging molecular techniques have yet to be universally standardized for Campylobacter spp. detection in water and food samples. Their consistency across laboratories and validation remains a challenge for regulatory acceptance.
Briefly, the cost-effective culture-based techniques are highly reliable and essential for live pathogen detection and regulatory compliance. On the other hand, molecular approaches rapidly generate sensitive results and thus are particularly useful for prompt outbreak response. However, high cost and necessary technical assistance restrict their large-scale applicability. Integrating both approaches could provide comprehensive, timely detection of pathogens by balancing the culturebased specificity and speed of molecular methods.
Campylobacter spp. isolation and detection from water and food sources is necessary for public health as these pathogens are associated with widespread enteric infections. The specificity and regulatory acceptance of ISO, FDA, and PHE-recommended culture methods make them the gold standard for Campylobacter spp. detection. The lengthy procedures and background flora-associated lower sensitivity are major limitations. These can be overcome by adding selective enrichment and plating media steps for the reliable identification of Campylobacter spp.
The use of chromogenic media is a cost-effective and rapid approach that yields varying colors in different Campylobacter spp. colonies. However, they should be standardized for consistent results at varying microbial contaminations in foods. The undetected VBNC cells of Campylobacter spp. are a major threat to food safety. The sensitivity of advanced immunological and molecular methods (PCR and PMA-qPCR) to Campylobacter spp. is higher than conventional procedures. However, the need for highly trained personnel and expensive apparatus limits their applications.
Integrating conventional culturing and recent molecular techniques is the way forward that could improve the specificity, speed, and sensitivity for robust surveillance of foodborne pathogens. Simultaneously, sustained adaptations and innovations are mandatory for Campylobacter spp. detection to ensure public health safety, particularly in low-resource regions. Standardization and validation of novel methods are necessary for improving Campylobacter spp. monitoring to curb their global infections.