
The global pork industry is increasingly dominated by a narrow set of high-yielding breeds (e.g., Large White, Landrace, Duroc, Pietrain, and Hampshire and their crosses), which reflects a troubling drift toward genetic uniformity (Sponenberg et al., 2019). Many local and heritage breeds have seen drastic population declines, with some now classified as threatened or extinct (fao.org/dad-is/risk-status-of-animal-genetic-resources/en/). Loss of such breeds not only represents a decline in biodiversity but also the disappearance of unique characteristics that may be critical for future challenges (e.g., climate change, new diseases). Conservation and valorisation of local pig breeds—through breeding programmes, niche markets, and policy support—are essential to reverse the erosion of genetic diversity and maintain a robust, adaptable pig farming sector (Čandek-Potokar et al., 2019).
This review focuses on native breeds from Central and South-Western Europe, which are still used in traditional pig production systems in this region due to their excellent adaptation and meat quality. Conventional breeds may be less suitable in regions where free-range environments predominate, as they are not conducive to large litters and pose a risk of piglet mortality. Climatic conditions in these regions may also be unfavourable for conventional breeds, and native breeds cope well with food shortages and make good use of feed (Leenhouwers & Merks, 2013; Wilkinson, 2011). Moreover, in the era of industrial food production, preserving biodiversity among pigs is essential globally. In 1992, the International Convention on Biological Diversity was established, which contributed to the creation of a global system for the protection of genetic resources of farm animals (Babicz et al., 2020). More than 70% of the factors responsible for biodiversity loss are related to food production and agriculture (Kalinowska et al., 2016). To reduce the adverse effects resulting from intensive production, it is crucial to gradually implement the principles of sustainable agriculture which are associated with the extensification of animal husbandry or organic production, and thus with the promotion and maintenance of native breeds (Eisler et al., 2014), especially in those areas where traditional pig production systems using native breeds still exist. These breeds represent a reservoir of genetic diversity within the pig species. They are associated with traditional, old-fashioned production methods and pork products, sometimes related to specific agro-ecosystems, such as the Iberian pig in the Dehesa (Rodriguez-Estévez et al., 2012). They have been neglected mainly due to their lack of competitiveness in industrial pig farming, with their lack of popularity being the main threat for their extinction. Moreover, African swine fever (ASF) can also be mentioned as a danger for native pig breeds in Europe. The disease spreads among wild boar populations, and despite the implemented biosecurity measures, outbreaks in domestic pigs were also noted (Szymańska and Dziwulaki, 2022; Škorput, et al., 2025).
Although the popularity of European local pig breeds has grown in recent years, as evidenced by a growing number of scientific publications on the subject, comprehensive syntheses remain limited. This explains the need for the present review, which aims to consolidate current knowledge and provide a clearer understanding of these breeds’ value in terms of meat quality.
Local autochthonous breeds of pigs are generally more robust in response to changing environmental factors (e.g., heat waves) and more resilient to diseases and parasites than crossbred and hybrid animals (Wojtysiak & Połtowicz, 2014). This means that local breeds’ productivity, fertility and meat quality are less affected by changes in environmental factors, including feed. In addition, these breeds can be fed diets based on natural resources, forage, and by-products without negative effects on health and welfare (Proorocu, 2021); thus, because these breeds are adapted to local and natural resources, they can efficiently utilise low-quality feed that does not compete with human food resources such as grains or legumes, and their breeding supports more sustainable food systems. Hence, they are a key pillar for smallholder farming and promote food security. There is, however, a drawback considering the profitability of rearing local breeds, as they are characterised by an unfavourable feed conversion ratio, increased carcass fatness, and lower carcass meatiness than commercial breeds (Kapelański et al., 1999). Varying carcass and meat quality is often the result of interactions between breed, feed management, and natural environmental factors (Rosenvold & Andersen, 2003). The breeding programmes for local breeds usually focus on preserving genetic diversity and limiting inbreeding levels rather than improving animal meat production, or robustness and foraging efficiency to make the best use of natural resources and by-products. Nevertheless, aiming to enhance the quality of pork may increasingly consider local breeds for commercial crossbreeding (Milczarek et al., 2012; Martins et al., 2020).
This review examines the carcass characteristics and meat quality of selected native pig breeds from Central and Southwestern Europe, with a focus on understanding the value of native breeds in terms of their unique traits, such as higher intramuscular fat content enhancing tenderness and juiciness, a favourable fatty acid profile rich in monounsaturated fats like oleic acid (notably in Iberian pigs), and superior sensory properties that reflect traditional, artisanal meat quality.
The European breeds studied here were selected from the FAO database (https://www.fao.org/dad-is/browse-by-country-and-species/en/) in May 2024, based on the criterion of having a census of at least 1,000 individuals (Table 1). even though these censuses may not have been exact. Subsequently, a literature search was immediately conducted using Google Scholar. The keywords used were names of the selected breeds and different slaughter and physicochemical parameters of pork meat (e.g., “pork”, “native breeds”, “genetic diversity”, “extensive farming”, “autochthonous breeds”, “organic farming”, “pH value”, “colour”, “cooking loss”, “shear force”, “drip loss”, “fatty acid profile”, “protein”, and “fat”) (Figure 1). For the final description in this review, only breeds with at least one research paper written in English on carcass or pork parameters were included. The most important information about the slaughter procedures and meat quality is included in Tables 1 – 4. Additional information has been added as supplementary tables.
Methodology for literature search
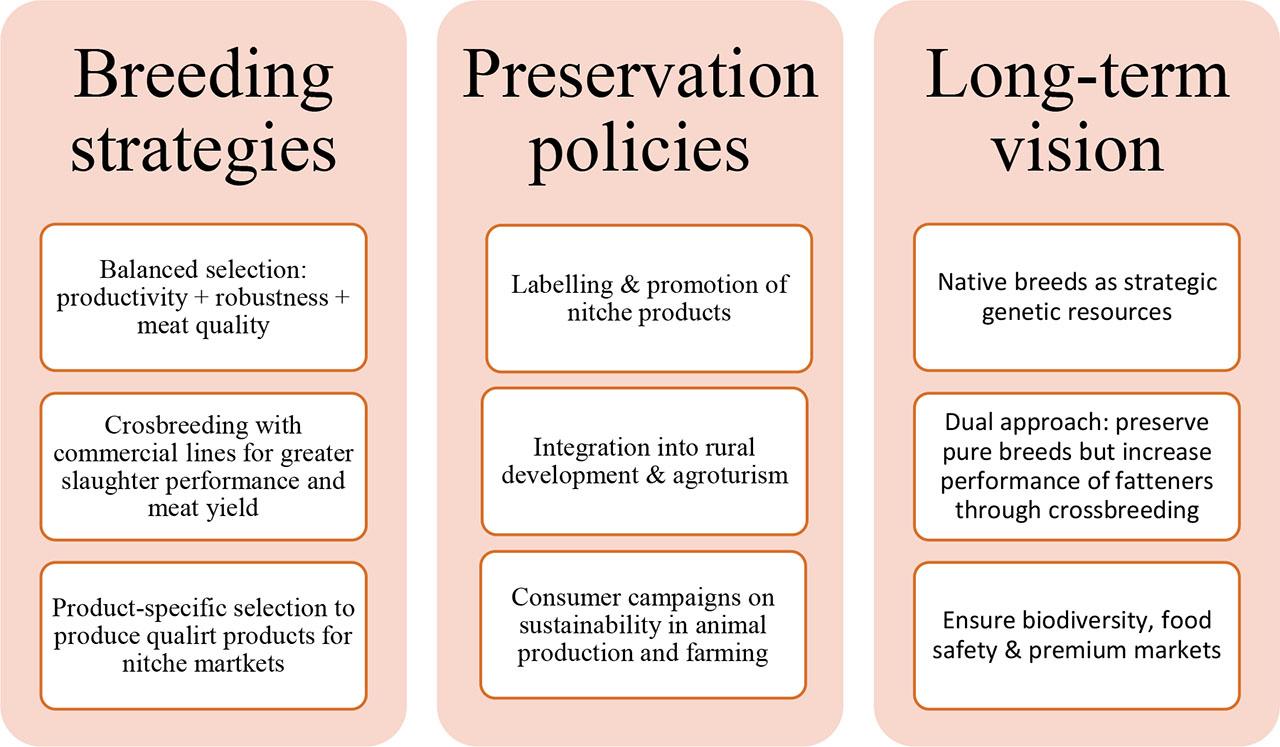
Recommendations scheme for European native pig breeds
Native pig breeds in selected European countries: population data and number of breeding females (FAO, 2024)
| Country | Breed | Total number of animals | Number of breeding females |
|---|---|---|---|
| SOUTH-WEST EUROPE | |||
| Italy | Cinta Senese | 1 992 (update 2023) | 692 (update 2023) |
| Apulo-Calabrese | 4 230 (update 2023) | 704 (update 2023) | |
| Mora Romagnola | 1 233 (update 2023) | 259 (update 2023) | |
| Nero Siciliano | 6 281 (update 2023) | 698 (update 2023) | |
| Spain | Iberian | 3 613 123 (2024) | 564 749 (2024) |
| Negra Canaria | 2 276 (2024) | 614 (2024) | |
| Porc Negre | 1 482 (2024) | 1 372 (2024) | |
| Mollorquí Porco Celta | 4 173 (2024) | 433 (2024) | |
| France | Gasconne | 1 760 (2024) | 1 591 (2024) |
| Portugal | Alentejana | 27 953 (2024) | 3 639 (2024) |
| Bisaro | 38 772 (2024) | 5 028 (2024) | |
| Malhado de Alcobaça | 3 301 (2024) | 429 (2024) | |
| CENTRAL EUROPE | |||
| Slovenia | Krskopolje | 2 006 (2024) | 296 (2024) |
| Poland | Pulawska | 3 404 (2024) | 2 488 (2024) |
| Zlotnicka Spotted | 1 056 (2024) | 936 (2024) | |
| Zlotnicka White | 1 214 (2024) | 1 058 (2024) | |
| Czech Republic | Prestice Black Pied | 900 (2022) | 394 (2022) |
| Belarus | Belorussion Large White | 2 958 300 (2006) | No data |
| Croatia | Black Slavonian pig | 3 200 (2023) | 2 143 (2023) |
| Hungary | Blond Mangalitsa | 3 385 (2023) | 3 332 (2023) |
| Red Mangalitsa | 1 542 (2023) | 1 503 (2023) | |
| Serbia | Mangalitsa | 4000 (2023) | 1954 (2023) |
| Ukraine | Mirgorod | 2 757 (2019) | 150 (2019) |
| Poltavian Meat | 1 392 (2023) | 130 (2023) | |
| Ukrainian Steppe White | 1 513 (2023) | 174 (2023) | |
Fattening and slaughter parameters of European native pig breeds
| Country | Breed | SW, kg | ADG, g | BF, mm last rib | BF, mm gluteus medius | DY, % | M, % | LMY, % | References |
|---|---|---|---|---|---|---|---|---|---|
| Italy | Cinta Senese | 127–147 | 416 | - | 20–49.3 | 82 | - | 72–76 | Pugliese et al. (2005), Sirtori et al. (2014) |
| Apulo-Calabrese | 144–157 | 359–413 | - | - | 83 | - | - | Micari et al. (2009), Scerra et al. (2024) | |
| Mora Romagnola | 193 | 376 | 57,1 | 62,1 | 80,4 | - | 39 | Fortina et al. (2005) | |
| Nero Siciliano | 62–121 | 162–600 | 39,1 | 39–49 | 79.1–82.5 | - | 40 | Pugliese et al. (2004), Liotta et al. (2005), Bozzi et al. (2019) | |
| Spain | Iberian | 152–190 | 563 | 7.63–80 | 65 | 77.9–81.2 | - | - | Tejeda et al. (2020), Serrano et al. (2009), Ayuso et al. (2018) |
| Porc Negre Mollorquí | 158 | 410 | 72 | 66 | 80,4 | - | - | Tibau et al. (2019) | |
| Porco Celta | 167.3 | - | 41.6–43.6 | 48,7 | 80,5 | - | - | Franco et al. (2014) | |
| France | Gasconne | ~175.3 | 439–472 | 44.7–47 | 46.1–49 | 78.7–80 | - | - | Mercat et al. (2019), Lebret et al. (2021) |
| Portugal | Alentejana | 100 | 333–583 | 38.1–55.6 | 39,4 | 73.3–80.9 | - | - | Freitas et al. (2007), Charneca et al. (2019), Martins et al. (2021) |
| Bisaro | 79 | 396 | 17–21,1 | - | 75.2–76.8 | - | - | Álvarez-Rodríguez Teixeira (2019), Charneca et al. (2019) | |
| Slovenia | Krskopolje | nd | - | 29 | 35.4–36.4 | 79.6–81.1 | - | 43–46 | Čandek-Potokar et al. (2003), Tomažin et al. (2018), Lukač et al.(2019) |
| Poland | Pulawska | ~100 | 638–810 | 23 | 12.73–28.86 | 75.33–80 | 49.4–56.11 | - | Wojtysiak & Połtowicz (2014), Debrecéni et al. (2018), Babicz et al. (2020), Milczarek (2021) |
| Zlotnicka Spotted | 107 | 590 | 41,3 | 28.9–34.96 | 76.51–80.5 | 41.83–46.28 | - | Kapelański et al. (2006), Grześkowiak et al. (2009), Bocian et al. (2012), Szulc et al. (2012) | |
| Zlotnicka White | >70 | - | 27,7 | 34,7 | 46,3 | - | Grześkowiak et al. (2009) | ||
| Czech Republic | Prestice Black Pied Pig | 94.2–115.2 | ~480 | 36.07–43.21 | 48.76–48.94 | Nevrkla, Václavková & Rozkot (2021) | |||
| Croatia | Black Slavonian pig | ~135 | - | 55 | 82 | - | 15–43 | Butko et al. (2007), Senčić et al. (2013), Kušec et al. (2019) | |
| Hungary | Blond Mangalitsa | 140–150 | 369 | - | - | - | - | 27 | Tomović et al. (2016), Nchama et al. (2023) |
| Mangalica in general term | 75–120 | 268–299 | - | 50.5–57 | 73–81.5 | - | - | Parunović et al. (2012), Hoha et al. (2016), Radović et al. (2017), Debrecéni et al. (2018), Charlton et al. (2022) | |
| Ukraine | Mirgorod | 100 | 632 | 30,9 | - | 72,5 | 50,4 | - | Tsereniuk et al. (2023) |
SW- slaughter weight; CW – carcass weight; CCW – cold carcass weight; HCW – hot carcass weight; ADG– Average daily gain; FBW – final body weight; BF – backfat; DY – dressing yield; M – meatiness; LMY – lean meat yield; nd – no data.
measured at 5 points,
dressing yield in the loin,
measured on the back,
from the age of 90 days to slaughter.
Physicochemical characteristics of meat from European native pig breeds
| Country | Breed | SW, kg | pH45 min | pH24h | L* | a* | b* | Cooking loss, % | WBSF, N, kg, kg/cm2 | Drip loss, % | References |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Italy | Cinta Senese | 127–147 | 6.2–6.4 | 5.82–5.78 | 46–50.1 | 11–14.9 | 2.9–5.4 | 17.9–30.3 | 94.14–99.87 N, 10.17 kg | 0.7–2.14 | Pugliese et al. (2005), Sirtori et al. (2014), Aquilani et al. (2019) |
| Apulo-Calabrese | 144.2 | - | - | 46.2–49.3 | 3.9–7.1 | 5.8–6.1 | - | - | - | Caparra et al. (2023), Scerra et al. (2024) | |
| Nero Siciliano | 62–121 | 6.18–6.29 | - | 46.7–50.1 | 14.7–15.3 | 4.9–5.8 | 23.5 | 9.99–10.11 kg | - | Pugliese et al. (2004) | |
| Mora Romagnola | 193 | 6,57 | 6,15 | 42,3 | 8,7 | 2,2 | - | - | - | Fortina et al. (2005) | |
| Spanish | Iberian | 152–190 | - | 5,62 | 39.8–41.7 | 8.7–11.3 | 6.4–13.6 | 24,1 | 4.33 kg/cm2 | - | Serrano et al. (2008), Fernández-Barroso et al. (2020), Tejeda et al. (2020) |
| Porco Celta | 167.3 | - | 5–46–5.55 | 47.3–50.7 | 10.4–13.1 | 10.7–11.7 | 19.7–21.9 | 3.64–3.8 kg/cm2 | 1.8–2.7 | Franco et al. (2014;2016) | |
| France | Gasconne | ~173.3 | 6,34 | 5.57–5.6 | 44–44.7 | 9.7–10.04 | 3.5–3.8 | 14,5 | 24.3 N | 1,7 | Mercat et al. (2019), Lebret et al. (2021) |
| Portugal | Alentejano | ~100 | - | 5.57–5.7 | 49.4–50.8 | 10.3–15.2 | 3.6–8.7 | 17.7–26.9 | 26.6 N | 1,3 | Martins et al. (2020;2021) |
| Bisaro | 79 | - | 5,44 | 55.1–55.7 | 3.4–7.8 | 4–10.9 | 28,9 | 47.4 N | 3,8 | Martins et al. (2020), Teixeira et al. (2021) | |
| Slovenia | Krskopolje | nd | 6,33 | 5.6–5.71 | 50,4 | 6,6 | 0,9 | 26,9 | 53.2 N | 4,2 | Čandek-Potokar et al. (2003), Lukač et al. (2019), Tomažin et al. (2018) |
| Poland | Pulawska | ~100 | 6.10–6.78 | 5.58–5.63 | 52.1–53.17 | 3.1–15.9 | 0.8–16.1 | 20.41–35.9 | 3.7–5.62 kg/cm2 | 1.7–24.89 | Wojtysiak & Połtowicz (2014), Bogucka & Kapelański (2016), Debrecéni et al. (2018), Milczarek et al. (2019), Milczarek (2021) |
| Zlotnicka Spotted | 107 | 6.38–6.57 | 5.56–5.67 | 47.9–49.3 | 4.77–17.2 | 2.5–5.04 | 27.46–29.36 | 43.61 N/cm2 | 2.36–3.4 | Grześkowiak et al. (2009), Bogucka & Kapelański (2016), Bocian et al. (2012), Szulc et al. (2012) | |
| Zlotnicka White | >70 | 6,38 | 5.53–5.72 | 43.9–49.54 | 6.3–14.23 | 3.34–4.9 | 22.76–27.62 | - | 3,4 | Grześkowiak et al. (2009), Migdał et al. (2017) | |
| Czech Republic | Prestice Black Pied Pig | 94.2–115.2 | 6.29 | 5.65 | 50.54 | 1.28 | 10.63 | 2.95 | Nevrkla, Václavková & Rozkot (2021) | ||
| Croatia | Black Slavonian pig | ~153.8 | 5,82 | 49,9 | 20 | 4,7 | - | - | - | Karolyi et al. (2004) | |
| Hungary | Blond Mangalitsa | 110–140 | 6.25–6.37 | 5.72–5.74 | 40.3–54 | 4–11.76 | 5.3–11.4 | 18,49 | 43.1 N | 5,9 | Tomović et al. (2016), Imrich et al. (2019) |
| Mangalica in general | 75–120 | 6,1 | 5.47–5.64 | 39.1–50.1 | 2.7–38.2 | 10.4–14.6 | 2.7–26.9 | 42.6 N, 5.05 kg | 4.8–8.8 | Stanišić et al. (2015), Radović et al. (2017), Debrecéni et al. (2018), Charlton et al. (2022), Nchama et al. (2023) | |
SW - slaughter weight; L* (lightness), a* (redness), b* (yellowness); WBSF - Warner–Blatzer shear force (values given in different units depending on the publication.
Protein, intramuscular fat content, total MUFA and PUFA fatty acids, and the n-6/n-3 ratio in LTL muscle in European native pig breeds
| Country | Breed | SW, kg | Rearing conditions | Protein, % | IMF, % | n-6/n-3 PUFA | Total MUFA (% of total FA) | Total PUFA (% of total FA) | References |
|---|---|---|---|---|---|---|---|---|---|
| Italy | Cinta Senese | 84 | outdoor | 23.09 | 6.66 | 12.5 | - | - | Aquilani et al. (2019) |
| 130 | indoor | 23.03 | 2.32 | 24.4 | 49.9 | 11.2 | Pugliese et al. (2013) | ||
| Apulo-Calabrese | 84.8 | outdoor | 20.66 | 1.86 | 8.2 | - | - | Caparra et al. (2023) | |
| 144–157 | indoor | 22 | 1.91–2.09 | 15.5*–2 | 48.8 | 12.4 | Aboagye et al. (2020), Scerra et al. (2024) | ||
| Nero Siciliano | 62–121 | outdoor | 22.24 | 5.77 | 10.7 | 53.3 | 10.9 | Pugliese et al. (2004) | |
| indoor | 23.42 | 6.25 | 18.3 | 42.2 | 14.4 | ||||
| Mora Romagnola | 193 | indoor | 23.3 | 6.1 | 47.6 | 11 | Fortina et al. (2005) | ||
| Spanish | Iberian | 152 | free range traditional system | 22.44 | 6.6 | - | 57.1 | 6.8 | Tejeda et al. (2020) |
| 145 | indoor | 21.5–21.6 | 6.61–8.6 | 17.7 | 52 | 6.5 | Pardo et al. (2021), Serrano et al. (2008) | ||
| Porco Celta | 167.3 | outdoor | 22.65 | 5.22 | 17.9 | 49.6 | 7.6 | Franco et al. (2014) | |
| France | Gasconne | ~175.3 | outdoor | 23.07 | 2.7 | 6.1–6.26 | 53.7 | 6.6 | Mercat et al. (2019) |
| Portugal | Alentejano | ~150 | outdoor | 23.6 | 6.9 | - | - | Martins et al. (2020) | |
| Bisaro | ~150 | outdoor | 23 | 6 | - | - | |||
| Slovenia | Krskopolje | nd | indoor | 23.2 | 2.81 | 15.7 | 47.5 | 11.6 | Lukač et al. (2019), Tomažin et al. (2018) |
| Poland | Pulawska | ~100 | indoor | 22.72–23.3 | 1.9–5.5 | 8.89–29.4 | 56.5 | 3.4 | Debreceni et al. (2018), Milczarek et al. (2021) |
| Zlotnicka Spotted | 107 | indoor | 23.27–25.2 | 2.04–2.6 | 10.78–12.94 | 49.4 | 11.1 | Grześkowiak et al. (2009), Debreceni et al. (2018) | |
| Zlotnicka White | >70 | indoor | 24.5 | 1.87 | 10.73 | 47.4 | 12.9 | Grześkowiak et al. (2009) | |
| Czech Republic | Prestice Blac Pied | 94.2–115.2 | indoor | 20.12 | 2.29 | 6.7 | 44.07 | 13.37 | Nevrkla, Václavková & Rozkot (2021); Belkova et al. 2023 |
| Croatia | Black Slavonian pig | ~135 | outdoor | 21.25 | 5.9 | Butko et al. (2007) | |||
| indoor | 21.3 | 4.95 | |||||||
| Hungary | Blond Mangalitsa | 110–140 | indoor | 21.83–24.64 | 1.34–5.86 | Tomović et al. (2016), Imrich et al. (2020) | |||
| Mangalica in general | 75–120 | outdoor | 21.45–21.7 | 7.57 | 9.2–12.97 | 44.8–55.5 | 6.5–15.6 | Parunović et al. (2012), Debrecéni et al. (2018) | |
| 75–120 | indoor | 19–24.1 | 1.9 | 17.92–37.3 | 57.2 | 5.9 | Parunović et al. (2012), Nchama et al. (2023) | ||
Content of total PUFA in % of IMF.
SW – slaughter weight; nd- no data; FA – fatty acids, MUFA – monounsaturated fatty acids, PUFA – polyunsaturated fatty acids, IMF – intramuscular fat, LTL – m. longissimus thoracis et lumborum.
Some general considerations regarding the performance of native pig breeds involve their use in extensive farming systems, which may include varying degrees of grazing. These systems are characterised by increased physical activity, dietary variability, lack of uniformity, longer production cycles, and the need to adapt fattening periods to the availability of natural resources or by-products. Additionally, management practices may aim to capitalize on compensatory growth during the fattening phase that follows a period of slower growth. Native breeds are also typically slaughtered at older ages and have higher weights than commercially improved breeds; Ludwiczak et al. (2023) have studied some of these factors in a recent review. The variety of breeds of each country are discussed in the following section.
Among the native Italian breeds, literature provides complete data on Cinta Senese, Apulo-Calabrese, Nero Siciliano, and Mora Romagnola. The fattening performance and slaughter procedures are presented in Table 2. Nero Siciliano is a breed from the island of Sicily in southern Italy that is traditionally reared under extensive conditions (Pugliese et al., 2004). It is characterised by the highest average daily gain (ADG) among Italian breeds, reaching up to 600g per day, which suggests better feed efficiency or adaptation for faster finishing (Bozzi et al., 2019). The lowest ADG was found in Cinta Senese (~ 400 g) and Apulo-Calabrese (ADG of ~300–400 g) (Caparra et al., 2023; Sirtori et al., 2014). Considering the slaughter procedures, Mora Romangola has the thickest backfat layer. Mora Romagnola comes from the Emilia–Romagna region in northern Italy. It is a slow-growing, heavy-weight type of pig (Bozzi et al., 2019), with the fat thickness ranging from 57 mm (at the last rib) to 62 mm (over the gluteus medius) (Fortina et al., 2005). Apulo-Calabrese and Mora Romagnola have the highest yields (~80–83%), making them productive in terms of meat output (Caparra et al., 2023; Fortina et al., 2005).
A similar balance between growth potential and product quality is also evident in Spain. The Iberian pig, the country’s most iconic native breed from the southwestern Iberian Peninsula (Tejeda et al., 2020), thrives in the extensive dehesa system, which enhances carcass quality while preserving the sensory characteristics of Iberian meat (Robina et al., 2013; Rodríguez-Estévez et al., 2009). To adapt to modern market needs, crossbreeding with Duroc has been introduced, improving economic efficiency while maintaining the high quality of dry-cured products such as Jamón Ibérico and Lomo Ibérico (Carrapiso et al., 2015). Quality depends on a combination of the genetics, the management system, and the final fattening diet (Ortiz et al., 2020). These factors are formalised in the Iberian pig quality standard (Real Decreto 4/2014, 2014), which defines fattening periods exceeding 12 months and slaughter weights up to 200 kg (Ayuso et al., 2018). In contrast, the Porc Negre Mollorquí reaches lower slaughter weights (~120 kilograms) and has a lower ADG (410 g vs. Iberian 563 g) (Ayuso et al., 2018; Serrano et al., 2009; Tibau et al., 2019). Another Spanish breed, the Porco Celta, differs in carcass characteristics, displaying the thinnest backfat (43–48 mm) (Franco et al., 2014; 2016).
In France, fewer breeds are documented, with the Gasconne pig being the main example (Table 1). It is a native breed of pig that survives in the foothills of the Pyrenees in southwestern France. This breed is characterised by a slow growth rate and a variable daily weight gain, with the adult body weight reaching 300 kg (Mercat et al., 2019). The slaughter yield for these pigs is 80%, and the backfat thickness is 44–50 mm (Lebret et al., 2021).
Portugal also contributes valuable genetic diversity with two traditional breeds: the Alentejano and the Bisaro. The Bisaro breed is maintained in northern Portugal, with the highest concentration of farms and animals observed in the Trás-os-Montes and Minho regions (Santos Silva et al., 2019). The meat from these animals is often smoked, which is typical for this region. The breed is characterised by slow growth (ADG ~390 g) and kept to a body weight of up to 180 kg (Santos Silva et al., 2019). Compared to the Bisaro pig, the Alentejano can reach greater ADG, from 330 to 580 g.
Most Central European countries have only a few native pig breeds, often adapted to traditional fat-based production (Table 1). In Slovenia, the Krškopolje pig – the only autochthonous breed – originates from Lower Carniola (Kastelic & Čandek-Potokar, 2013). Similarly, in neighbouring Croatia, one of the three native breeds, the Black Slavonian pig, is known for producing lard, bacon, and traditional high-quality meat products (Jelić et al., 2023). Both breeds share a common trait of developing a thick backfat layer, underlining their role in regional fat-rich culinary traditions. A comparable focus on fat production is also evident in the Mangalica, a Hungarian breed that traces its roots to the Serbian Šumadija. The Sumadija was brought to Hungary in the 19th century and crossed with local breeds such as the Alfoldi, Bakony, and Szalonta (Hoha et al., 2016; Addo & Jung, 2022). The Mangalica fatteners are slaughtered at an average body weight of 130 kg and are characterised by 65–70% fat in the carcass (Egerszegi et al., 2003). Poland, by contrast, has preserved three native breeds with a more balanced profile: Puławska, Złotnicka Spotted, and Złotnicka White. The Puławska, the country’s oldest native pig, combines heritage with productivity, reaching ADG values over 800 g/day (Babicz et al., 2020), making it one of the fastest-growing native breeds in Europe. Currently, this breed is a transitional form between the fat-meat type and the meat type. Among all Polish native breeds, Złotnicka Spotted is characterised by the thickest backfat layer of 41.3 mm, and at the same time, the lowest meat yield, of 41.83 % (Bocian et al., 2012; Szulc et al., 2012). As for Eastern Europe, only the Ukrainian Mirgorod pig is described in the literature. It was created in the Poltava region in Ukraine due to crossing the Ukrainian domestic pigs of variegated colours with boars of large white, Berkshire, and Tamworth breeds (Transpov, 2016). Mirgorod pigs are characterised by an average daily gain of 630 g, which places them among the fastest-growing native pigs in Europe (Tsereniuk et al., 2023). The Preštice Black-Pied Pig is the only indigenous Czech pig breed, developed in the 19th century through crossing local animals with English and German breeds. It is classified as a fat-meat type, traditionally used in the production of regional pork products due to its higher backfat thickness and moderate intramuscular fat (2.3–2.8%). Carcasses of pigs slaughtered at 94–115 kg show lean meat yields of around 48.8% and fat depths exceeding 36 mm. This breed is valued for its robustness, adaptability, and contribution to preserving Czech culinary heritage (Nevrkla et al. 2021). To summarise, most of the reviewed native breeds have a similar dressing yield between 70% and over 80%, and the lean meat content in the carcass is on average 40%. The highest daily gains were reported for Mirgorod (Ukraine), Puławska (Poland), Nero Siciliano (Italy), while the thickest backfat layer was in Mora Romagnola (Italy), and Mangalica (Hungary).
The most crucial physicochemical parameters of pork from selected native pig breeds are presented in Table 3. The parameters described in this chapter (i.e., pH, colour, water loss, and Warner–Bratzler shear force) were measured on the longissimus thoracis et lumborum (referred to as the LTL).
The pH value is most commonly recorded in three time periods postmortem: 45 minutes (pH45 min), 24 hours (pH24 h), and 48 hours (pH48 h). The pH value indicates the extent of changes occurring in the meat and indicates its quality and culinary suitability. Meat obtained directly after slaughter is characterised by a pH of 6.8–7.3. In this state, such pork has the highest capacity to bind water but has an unfavourable palatability and tenderness. During the progressive post-slaughter changes resulting from glycolytic processes, the pH value of the meat decreases until it reaches the ultimate level. This depends on the species, preslaughter stress, degree of bleeding, cooling, or content of fat and connective tissue in the meat. All the breeds had very similar pH values measured between 45 minutes and 24 hours postmortem. The pH24h ranged from 5.44 in the Bisaro breed (Martins et al., 2020) to 6.15 units in the Roma Romagnola (Fortina et al., 2005). Their highest values of pH ultimate (6.57 units) were reported in pork from the Mora Romagnola pigs (Fortina et al., 2005). For Prestice Black-Pied pigs, Nevrkla et al. (2018) report pH45min is 6.29 and pH24h is 5.65, values which are somewhat higher than in many commercial hybrids and indicate good meat stability. The lowest pH24 h values (< 5.5) were found in the meat of Porto Celta (Spain|), Bisaro (Portugal), and Mangalista (Hungary).
Another parameter that has been examined in most publications is meat colour (CIE - Commission Internationale de l’Éclairage), which includes three main coordinates: L* (lightness), a* (redness), and b* (yellowness), and allows for quality classification. It is also no less important in the context of consumer preference studies. The primary muscle’s pigment, myoglobin, converts into different chemical forms, depending on the storage conditions, including oxygen availability, temperature, and the final pH value in meat (Suman & Joseph, 2013). Consumers usually prefer meat with an intense, bright red colour. However, various factors such as breed, housing system, and nutrition, affect meat colour, thus the values reported in studies often show a wide range. Studies show that an L* value in the range of 49–60 for the LTL is perceived as desirable by consumers (Warriss & Brown, 1995). The L* (lightness) was the lowest (~40.0 units) in pork of the Iberian pigs (Spain), Mangalitsa (Hungary), and Mora Romagnola (Italy), meaning that those pigs had the darkest meat. Another essential colour coordinate is redness, marked with a*. The reddest meat (a*> 10.0 units) was found in Nero Siciliano (Italy), Cinta Senese (Italy), Porto Celta (Spain), and Alentejano (Portugal). The highest value was recorded for the Black Slavonian pig (a*= 20.02) (Karolyi et al., 2004). These findings suggest a clear interaction between pH decline and colour, reinforcing their joint role in determining consumer perception.
The ability of meat to retain water—both during storage (drip loss) and heat treatment (cooking loss)—is also influenced by pH and protein structure, creating another layer of association with the parameters above. Meat has the property of retaining water under the influence of force, and it is the water capacity (WHC) (Kilic et al., 2023; Torres Filho et al., 2017). On average, 90% of the total water is found in muscle proteins, and the remaining 10% is retained by connective tissue. A critical parameter of processed meat during heat treatment is cooking loss. During this process, the moisture content in the heat-treated product is significantly reduced (Vujadinović et al., 2014). The LTL of native pigs is characterised by a cooking loss of 14.52–35.92%. The lowest value for this parameter was recorded for the Gasconne pig - 14.5% (Lebret et al., 2021). In contrast, just over 17% was obtained for the Alentejano breed (17.7%) and the Cinta Senese pig breed (17.94%) (Martins et al., 2020; Sirtori et al., 2014). The lowest drip loss value was recorded for the Cinta Senese breed, at 0.7% (Pugliese et al., 2005), whereas the highest value, 8.8%, was obtained for the Mangalica (Debrecéni et al., 2018). The methodology may differ slightly between publications, which could constitute an additional source of variation. The type of heat treatment influences water loss, and it has been shown that meat cooked at temperatures between 60°C and 80°C exhibits lower water loss, resulting in more balanced and beneficial cooking loss compared to heat treatment by baking (Vujadinović et al., 2014). It has also been shown that the pH during meat storage affects water loss, with higher pH levels resulting in lower losses (Drummond & Sun, 2006). Cooking loss has a wide range among the native pig breeds (14.52–35.92%, Table 3), with the Gasconne pig showing the lowest value at 14.5%, indicating well-developed WHC. The Alentejano and Cinta Senese also have relatively low cooking losses (~17.7% and 17.94% respectively). The Cinta Senese again shows good WHC with the lowest drip loss (0.7%) (Pugliese et al., 2005; Sirtori et al., 2014; Aquilani et al., 2019), while the Puławska breed shows a much higher value, reaching up to 24.89 %, suggesting poorer ability to hold moisture (Wojtysiak & Połtowicz, 2014; Bogucka & Kapelański, 2016; Debrecéni et al., 2018; Milczarek et al., 2019; Milczarek, 2021).
Finally, texture, commonly measured as Warner–Bratzler shear force (WBSF), integrates the above parameters into the eating quality perceived by consumers. Tenderness depends not only on muscle structure but also on WHC and pH, which affect juiciness and fibre resistance. The measured parameters are typically shear force (WBSF), shear energy, and Young’s modulus. The shear force was chosen because it was the most prominent type of analysis in the cited literature. Table 3 presents the values for this parameter obtained using meat samples tested under various conditions. Notably, some analyses were performed on raw meat, while others were performed on thermally processed samples, resulting in a wide range of values. The values for WBSF were given in several units, as reported in the source publications. Pugliese et al. (2004) and Sirtori et al. (2014) examined the WBSF of fresh meat, which was an average of 10 kg (Nero Siciliano and Cinta Senese). Some of the publications in Table 3 reported values for this parameter in kg/cm2. In this case, the highest value was recorded for the Iberian breed (4.33 kg/cm2), and the lowest value was recorded for the Porco Celta breed (3.6 kg/cm2) (Franco et al., 2014; Fernández-Barroso et al., 2020). In turn, the highest values reported by the authors were shown for the Cinta Senese breed (94–99 N), and the lowest values were reported for the Gasconne breed (24.3 N) (Pugliese et al., 2005; Lebret et al., 2021).
The comparison in Table 4 shows that the protein content in the LTL is relatively uniform, generally ranging from 20% to 25%. The lowest protein content was observed in Mangalica pigs (19–21%), as reported in the studies of Parunović et al. (2012) and Nchama et al. (2023). While protein provides structural and nutritional value, the balance between protein and intramuscular fat (IMF) ultimately shapes the sensory and technological properties in pork. The IMF content, which determines marbling, has been repeatedly identified as a key driver of meat palatability and juiciness (Teixeira et al., 2021). It is generally recommended that its level in the LTL muscle averages around 2.5% (Sládek & Dračková, 2020). In the cited publications in Table 4, the IMF does not exceed 9%, but values as low as 1.3% have been reported, for instance, in the Mangalica breed (Imrich et al., 2020). Such variation may stem from differences in breed genetics and methodological inconsistencies across studies. Beyond genetics, environmental and management factors—such as slaughter age, body weight, and feeding regime—play a central role in IMF deposition (Lebret, 2008; Wood et al., 2008). Energy-rich diets with high starch-to-protein ratios, lipid supplementation, and extended finishing periods typically increase IMF levels, while restricted feeding and high-fibre diets can reduce them. Thus, IMF represents a complex trait shaped by the interaction between breed and management, directly influencing not only marbling but also the nutritional quality of pork (De Tonnac et al., 2023).
This link between the IMF and nutritional value becomes even more evident when considering the fatty acid profile of native breeds. The composition of fatty acids—particularly the balance of saturated, monounsaturated (MUFA), and polyunsaturated fatty acids (PUFA)—is increasingly recognised as a determinant of the contribution of pork to human health (Li et al., 2015). The contents of n-3 and n-6 polyunsaturated fatty acids (PUFAs) and their proportions are associated with human diseases, including obesity and cardiovascular diseases, and a high content of n-3 polyunsaturated fatty acids and a low ratio of n-6/n-3 polyunsaturated fatty acids are more beneficial (Simopoulos 2016). The recommended n-6/n-3 PUFA ratio ranges from 1:1 to 4:1 (Simopoulos, 2002, 2006). Still, nutritional standards state that the proper ratio of n-6 to n-3 PUFA in a daily diet should be in the range of 6:1 to 4:1 (Grześkowiak et al., 2010). The n-6/n-3 ratio in pork can be influenced by the animal’s diet, breed, and husbandry system. In Table 4, the ratio of n-6 to n-3 PUFAs varies between breeds and significantly deviates from the recommended value. The values closest to the ratio of 6:1 were found for three native breeds: Apulo-Calabrian (n-6/n-3= 1.99) (Scerra et al., 2024), Gascon (n-6/n-3= 6.1–6.2) (Mercat et al., 2019) and Prestice black pied breed, which had 6.7:1 (Nevrkla et al. 2021). A less favorable n-6/n-3 PUFA ratio was noted in Cinta Senese kept indoors (24.4), and Pulawska (up to 29.4). The highest MUFA content was found in the meat of the Nero Siciliano breed: 53% (Pugliese et al., 2004) and in pork from native breeds from Spain and France, for which the value of this parameter ranged from 49.5 (Porco Celta) to 52–57.1% (Porco Celta and Gasconne) (Tejeda et al., 2020; Lebret et al., 2021; Pardo et al., 2021). Lower MUFA levels were observed in some Cinta Senese (49.9%) and Porco Celta (49.6%) groups, though still within a favorable range. The last parameter included in Table 4 is the total PUFA content. The highest value, 15.7%, was recorded for the Apulo-Calabrese breed, whereas the lowest value, 3.39%, was recorded for the meat of the Pulawska pigs (Milczarek, 2021). PUFA content was low in Iberian (6.5–6.8%) and Mangalica (5.9–15.6%) – indicating more stable fat with less susceptibility to oxidation. These findings confirm that IMF quantity and fatty acid quality are inseparably linked characteristics—both influenced by breed genetics, diet, and management—and together they determine not only consumer satisfaction but also the nutritional value of pork. MUFA levels are consistently high in Mediterranean and Central European fatty-type breeds (Iberian, Mangalica, Pulawska), reflecting their traditional selection for meat quality and flavour. Native Northern and Eastern European breeds tend to have higher PUFA than Iberian or Mangalica, which may impact fat firmness and shelf life negatively but contribute to a healthier lipid profile. Breeds raised in outdoor/free-range systems often show a better (lower) n-6/n-3 ratio, highlighting both genetic and environmental influences.
This comprehensive review of native pig breeds across Southern, Central, and Eastern Europe highlights the diversity of genetic traits, regional production systems, and resulting meat qualities that define traditional pig farming. Native breeds such as the Mirgorod (Ukraine), Puławska (Poland), and Nero Siciliano (Italy) show the highest average daily gains (ADG), indicating high performance under appropriate husbandry conditions. On the other hand, breeds like the Mora Romagnola (Italy) and Mangalica (Hungary) stand out for their exceptionally thick backfat, contributing to their suitability for specific traditional meat products. Breed strongly affects the fatty acid composition of pork: Mediterranean fatty-type breeds (Iberian, Alentejano, Cinta Senese, Mangalica) are characterized by high IMF and MUFA, with relatively low PUFA – ideal for traditional dry-cured products. Northern/Eastern European breeds (Pulawska, Zlotnicka, Krskopolje) generally have lower IMF and higher PUFA, resulting in leaner but less oxidative-stable fat. N-6/n-3 ratios are influenced by both genetics and rearing system, but outdoor-raised pigs typically show more favorable ratios.
Regarding carcass characteristics, most breeds demonstrate comparable slaughter yields, yet differ in lean meat content and backfat thickness, reflecting adaptations to specific production goals. Meat quality traits, including pH, colour, water-holding capacity, and texture, further differentiate these breeds. Breeds like Cinta Senese, Gasconne, and Alentejano demonstrate low cooking and drip losses, indicative of strong water-holding capacity, which is a desirable trait in processed meat production. Meanwhile, breeds such as Black Slavonian and Nero Siciliano deliver intensely red meat, appealing to consumer preferences for richer meat colour. Texture and tenderness vary widely, influenced by genetic factors and meat processing methods. Notably, Gasconne pigs show the lowest shear force, suggesting superior tenderness, while Cinta Senese exhibits the highest, possibly linked to muscle structure and intramuscular fat content. Lastly, the chemical composition, particularly intramuscular fat, confirms the marbling potential of these breeds. While IMF varies across studies due to methodological differences, breeds such as Mangalica and Alentejano offer high marbling, contributing to superior flavour and palatability.
Based on this literature review, two main paths in pig production can be brought to light. 1. The path of industrial production breeding and rearing a few commercial breeds and their crosses. This maximises efficiency but reduces resilience, robustness, and uniqueness of meat attributes. 2. The path of sustainable/niche/extensive production, where native breeds are maintained for biodiversity, and premium-quality products. This maximises the genetic diversity and promotes local products.
The future of native breeds is mainly related to their role as genetic reservoirs for characteristics that may become vital in adapting to future challenges. However, there is one other perspective that may contribute to the increased popularity of these breeds. A modern consumer's attitude toward healthy nutrition shifts, leading them to search for quality products derived from sustainable animal production. EU consumers realise that while industrial systems prioritise quantity, native breeds emphasise unique meat quality from animals produced under high welfare standards. Above that, the role and future of native pig breeds extends beyond cultural preservation to sustainability, biodiversity, and food security. To summarise this review’s findings, a recommendations scheme for European native pig breeds has been created (Figure 2). In the case of breeding strategies, future breeding programmes should focus on all three groups of characteristics: productivity, disease resistance and robustness, and meat quality. Native breeds could be crossed with commercial lines to introduce robustness and quality traits without losing growth and feed efficiency performance. Breeding should focus on characteristics aligned with market niches (e.g., high oleic acid for health-conscious consumers, backfat for cured products). Among breed preservation policies, the inclusion of native pig farming in rural development strategies and agrotourism should be encouraged. Consumer awareness should be increased by promoting the cultural heritage, nutritional benefits, and sustainability of native breed pork to stimulate demand and sustain premium markets. From a long-term perspective, native pig breeds should not be viewed as relics of the past but as strategic genetic resources for future food security.
In summary, native pig breeds in Europe reflect a valuable balance between cultural heritage, genetic diversity, and meat quality. While they often grow more slowly than commercial hybrids, they provide unique sensory characteristics prized in traditional products. Genetic conservation and sustainable farming practices are essential to maintaining their role in regional economies and food traditions.