
Fish from aquaculture have been more widely available to consumers, which has been attributed to a 12-fold increase in production over the last thirty years (Ritchie and Roser, 2024). Today, aquaculture accounts for one in three fish consumed by humans. As a result of the rapid growth of production, there is currently a demand for aquafeeds (Awad et al., 2024; Mohammady et al., 2023, 2025; Refaey et al., 2023). It is essential to note that the viability of aquaculture is dependent on a variety of factors, one of which is the availability of inexpensive feed (Tacon et al., 2011). The development of more affordable aquafeed would greatly benefit the rapid expansion of the aquaculture sector since feed expenses constitute around 60% of total operational costs (D’Abramo, 2021; FAO, 2022). Since there is a lack of interest in traditional aquafeed ingredients like fish meal, readily available alternative protein sources are required. The proportions of these ingredients in marketed aqua-feeds have decreased recently as a result of many studies looking at the advantages of replacing fish meals with plant components.
However, employing plant-based products has some known disadvantages, most notably when comparing the amounts of cholesterol, fatty acids, and amino acids in marine and plant feed components, as well as the presence of anti-nutritional substances in plant feedstuffs (Hassaan et al., 2015, 2018; Sison, 2016). Significant adverse outcomes are linked to extensive substitution, and these composition differences may impact feed utilization efficiency. For instance, when young Atlantic salmon (Salmo salar) were given a diet where 80% of the fish meal was replaced with plant-based ingredients, their developmental results were reduced (Kousoulaki et al., 2022). When juvenile rainbow trout (Oncorhynchus mykiss) were provided a diet that included marine materials, they showed more development compared to the fish fed a completely plant-based diet (Lazzarotto et al., 2018). The researchers of these trials postulated that the observed decrease in growth and the effects on cholesterol metabolism were primarily attributed to the utilization of plant-based meals rather than fish meals.
While certain research studies have indicated that providing animals with plant protein diets may adversely affect their performance, previous reports have demonstrated that by employing the following nutritional strategies, it is possible to sustainably feed aquatic animals plant protein diets without experiencing such adverse effects: The inclusion of deficient amino acids (Goda et al., 2007; Liti et al., 2006) and utilization of external enzymes (Hassaan et al., 2019, 2021; Jiang et al., 2014; Soltan et al., 2023; Wiszniewski et al., 2022) is a situation that has been seen across multiple plant sources. The study conducted by Nandeesha et al. (2002) examined the implementation of a fish meal-based diet followed by a day of plant-based feed. In addition to other contemporary dietary techniques, the inclusion of specific additives has been explored as a viable alternative (Aksnes et al., 2006; Johnson et al., 2015; Lee et al., 2015; Sarker et al., 2007).
It is anticipated that fish meal will cease to serve as a substantial protein source in the fish diet, leading to a potential increase in the use of fish meal-free diets in the near future. Consequently, the future expansion and viability of aquaculture may heavily rely on the identification of innovative, suitable, and cost-effective plant protein constituents that may replace fish meal without compromising animal performance (Gatlin III et al., 2007). A number of scientists initiated efforts to substitute fish meals with more cost-effective plant protein sources in response to the positive outcomes shown in the above-mentioned previous investigations. To investigate the impact of plant proteins on fish growth and development in the context of substituting fish meals in feeds, this review thoroughly evaluates several pieces of research conducted on the replacement of fish meals with plant protein components in fish. These studies have examined the effects of substituting fish meal with plant protein on various aspects of fish performance, including growth performance, and cholesterol metabolism gene expression. The content provided in this study is expected to serve as a catalyst for researchers to incorporate and explore more plant protein constituents in the development of aquaculture diets. Furthermore, it will necessitate careful formulation and supplementation strategies to ensure the nutritional adequacy of plant-based diets in aquaculture systems. Lastly, it would encourage the reduction of fish meal use in diets to ensure the long-term viability of fish farmers who depend on fish meal for their feed needs.
Replacing fish meal with plant protein in aquafeeds exerts multifaceted effects on growth performance, and outcomes depend strongly on species, degree of replacement, processing of plant ingredients, and use of functional supplements and amino acid fortification (Egerton et al., 2020; Mugwanya et al., 2023; Qian et al., 2024). Across carnivorous and many omnivorous fishes, high fish meal substitution with untreated plant meals frequently depresses feed intake, growth rate, and protein retention, but a growing body of work also demonstrates that well-balanced blends of processed plant proteins, supplemented with limiting amino acids, cholesterol, or protein hydrolysates, can sustain growth and feed efficiency comparable to fish meal-based diets at moderate-to-high replacement levels (Daniel, 2018; Mugwanya et al., 2023).
Many studies report that diets strongly depleted in fish meal and dominated by conventional plant meals reduce voluntary feed intake and specific growth rate in several cultured species, particularly carnivores, largely because plant protein sources differ from fish meal in amino acid balance, palatability, digestibility, and antinutrient load. Meta-analyses focused on carnivorous fish indicate that excessive fish meal replacement with generic plant proteins often increases feed conversion ratio and compromises protein efficiency ratio, especially in marine carnivores, even when survival is not affected (Qian et al., 2024). Mechanistically, reduced apparent digestibility coefficients of some legume and oilseed meals, damage to intestinal epithelium, altered gut microbiota, and exacerbated inflammatory responses collectively impair nutrient assimilation and thereby constrain growth despite adequate dietary crude protein (Qian et al., 2024).
Plant-rich formulations may also induce hypocholesterolemia, distort lipid metabolism, and depress growth in fish that evolved on animal-based prey, because most plant ingredients are naturally low in cholesterol relative to fish meal. Studies using high soybean meal diets show that, in the absence of adequate exogenous cholesterol, fish exhibit reduced weight gain, impaired lipid deposition patterns, and altered plasma cholesterol fractions, even when protein and energy levels are formulated to be isonitrogenous and isoenergetic with fish meal controls (Deng et al., 2009; Twibell and Wilson, 2004). In addition, some works associate high plant protein inclusion with elevated muscle protein degradation, modifications in muscle fiber morphology, and up-regulation of lysosomal proteolysis pathways, all of which diminish net protein accretion and thus growth performance under high fish meal replacement (Alami-Durante et al., 2010; Snyder et al., 2012).
A central constraint on plant protein utilization is their typically lower apparent digestibility and the presence of heat-labile and heat-stable antinutritional factors, including protease inhibitors, lectins, phytic acid, saponins, and non-starch polysaccharides, which can reduce nutrient availability and damage gut integrity (Francis et al., 2001; Lifsey et al., 2020). These antinutrients can precipitate intestinal enteritis, villus atrophy, and changes in mucosal barrier function, thereby decreasing absorptive surface area and contributing to reduced feed efficiency and growth (Hussain et al., 2024). In addition, many plant proteins are deficient or marginal in one or more indispensable amino acids, particularly lysine, methionine, and in some cases threonine and tryptophan, and uncorrected imbalances not only limit protein synthesis but may also result in increased catabolism of amino acids for energy, exacerbating poor protein retention (Egerton et al., 2020).
Processing technologies such as fermentation, enzymatic treatment, or protein concentration can mitigate several of these issues by degrading antinutrients, improving protein solubility and digestibility, and modifying flavor and palatability. A recent systematic review and meta-analysis on fermented plant proteins in aqua-feeds concluded that substituting fish meal with fermented plant protein can enhance growth performance, feed utilization, and antioxidant and digestive enzyme activity across multiple species, if replacement levels and species-specific tolerances are respected (Mugwanya et al., 2023). Similarly, a meta-analysis focusing on carnivorous fishes showed that deep-processed ingredients produce far fewer negative outcomes than raw meals when less than half of fish meal protein is replaced, underscoring the importance of ingredient quality rather than plant origin alone (Qian et al., 2024).
Despite the documented risks at high inclusion, numerous studies demonstrate that well-formulated plant protein blends can partially or even largely replace fish meal without compromising growth, especially when replacement is moderate and diets are carefully balanced. For example, experimental diets using mixed plant proteins such as cottonseed and rapeseed meals in yellow catfish showed that replacing a substantial proportion of fish meal with a 2:3 cottonseed-to-rapeseed protein blend maintained growth performance and liver protein content comparable to fish meal controls at lower replacement levels, though excessive substitution triggered oxidative and inflammatory responses. In marine species like ballan wrasse, plant-based protein ingredients, notably soy protein concentrate and pea protein, successfully substituted fish meal without adverse effects on growth, survival, or feed efficiency when the marine-to-plant protein ratio was kept within a moderate range and diets remained isonitrogenous (Cavrois-Rogacki et al., 2022; Han et al., 2022; Mugwanya et al., 2023).
Species-specific differences in tolerance to plant protein are evident in Atlantic salmon (Egerton et al., 2020), largemouth bass (Liu et al., 2021) rockfish (Kim and Cho, 2024) and Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂ (Ye et al., 2020). Meta-analytical results from Mugwanya et al. (2023) and Qian et al. (2024) showed African catfish, hybrid striped bass, and black sea-bass generally utilize plant protein more efficiently than groupers and Japanese seabass, the latter showing more pronounced growth suppression with rapeseed-rich formulations. Moreover, for Atlantic salmon parr, one study found that reducing fish meal from 35% to 15% of the diet and replacing it with plant proteins led to reduced growth, whereas supplementing high-plant protein diets with fish protein hydrolysates restored growth to levels equivalent to fish meal-rich control feeds, highlighting the value of functional animal protein fractions within predominantly plant-based formulations (Egerton et al., 2020). Collectively, these findings support the view that fish meal can often be reduced by roughly 40–60% of total dietary protein in many species when plant protein is used as protein concentrates or isolates, combined in complementary blends, and fortified with limiting amino acids.
Cholesterol is abundant in fish meals but lacking in most plant-based substances (Cheng and Hardy, 2004; Zhang et al., 2019). In addition, feeding 1.0% cholesterol significantly increased the weight growth rate and feed intake in juvenile channel catfish fed a diet based on soybean meal (Twibell and Wilson, 2004) and Japanese flounder fed a diet based on soybean meal (Deng et al., 2010). These studies suggest that combining more plant protein sources or adding supplements like cholesterol can help in the growth of fish. Table 1 shows some studies supporting the idea that fish meals fully and partially replaced by a mixture of plant proteins or the addition of supplements in fish diets did not affect the animals’ performances.
Studies of fish meal fully and partially replaced by plant protein in fish diets
| Species | Plant protein ingredients | Fish meal (FM) replacement level | Main effects on growth and feed utilization | References |
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 |
| Larval cobia (Rachycentron canadum) | Mixed plant proteins and oils in larval diets | Plant proteins and oils as major protein and lipid sources | Larval cobia showed good growth on diets containing a wide range of plant proteins and oils, supporting the feasibility of early plant-based feeding. | Craig et al. (2006) |
| Juvenile Siberian sturgeon (Acipenser baerii) | Extracted soybean meal (partially replacing animal-origin proteins: fish meal and meat-and-bone meal) plus crystalline amino acids (L-lysine, DL-methionine) in some treatments | Partial replacement of animal protein by extracted soybean meal; tested with and without AA supplementation | Partial replacement of fish meal and meatandbone meal with extracted soybean meal reduced growth; however, supplementing L-lysine and DL-methionine improved growth and FCR, highlighting the importance of amino acid balancing in high soybean diets. | Mazurkiewicz et al. (2009) |
| Juvenile Japanese flounder (Paralichthys olivaceus) | Soybean meal, corn gluten meal, and soybean phospholipid | Diets with graded soybean meal; 24% dietary SBM identified as safe level | SBM inclusion above 24% adversely affected growth and protein and lipid metabolism, whereas 24% SBM supported normal growth and feed utilization. | Deng et al. (2006) |
| Cobia (Rachycentron canadum) | Rapeseed meal and other plant-based ingredients (in diets comparing FM replacement by rapeseed meal) | Graded inclusion of rapeseed meal | Growth and body composition were maintained up to moderate rapeseed meal levels; excessive inclusion reduced growth, indicating a species-specific tolerance threshold. | Luo et al. (2012) |
| Hybrid sturgeon (Acipenser baerii × A. schrenckii) | Corn gluten meal as the major plant protein | Up to 55% corn gluten meal replacing part of FM | Corn gluten meal up to 55% did not affect growth or FCR and reduced feed cost by about 30% compared with fish meal-based diets. | Sicuro et al. (2012) |
| Siberian sturgeon (Acipenser baerii) | Mixture of soybean meal and wheat gluten meal, fortified with crystalline EAAs and monocalcium phosphate | 100% of dietary protein from the soy–wheat gluten mixture (fish meal free) | Complete fish meal replacement with soybean meal and wheat gluten, when supplemented with crystalline amino acids and mineral phosphorus, produced no adverse effects on growth or protein utilization. | Yun et al. (2014) |
| Rainbow trout (Oncorhynchus mykiss) | Plant protein concentrates (mainly soy, pea and wheat fractions) plus multiple EAA supplementations, krill meal and krill soluble as attractants | 100% FM substitution by plant concentrates | Total fish meal replacement with adequately supplemented plant protein concentrates showed no reduction in feed intake or growth, demonstrating feasibility of completely plantbased protein in salmonids when attractants and EAAs are used. | Lee et al. (2015) |
| Juvenile Amur sturgeon (Acipenser schrenckii) | Soy protein isolate (SPI) as principal plant protein | Replacement series up to 100% FM with SPI | Inclusion of SPI at high levels depressed growth and feed utilization; brokenline analysis recommended a maximum of about 57.6% fish meal replacement by SPI to avoid significant growth and FCR deterioration. | Jiang et al. (2018) |
| Atlantic salmon (Salmo salar) | High-level mixed plant proteins (soy protein concentrate, wheat gluten, pea protein and other plant concentrates) plus partly hydrolyzed and soluble fish proteins | Diets with 80% of protein from plant sources; fish protein hydrolysate/soluble fractions at 10% | Reducing fish meal from 35% to 15% with high plant protein decreased growth but adding 10% partly hydrolyzed or soluble fish protein to the 80% plant-protein diet restored growth to the level of fish meal-rich controls and increased circulating branched-chain amino acids. | Egerton et al. (2020) |
| Multiple species (metaanalysis) | Fermented soybean meal, fermented rapeseed meal, and other fermented plant proteins | Wide range; many trials up to full FM replacement in experimental diets | Systematic review found that replacing FM with fermented plant proteins can maintain or even improve growth, FCR and health markers compared with conventional plant meals, thanks to higher digestibility and lower antinutrient content. | Mugwanya et al. (2023) |
| Yellow catfish (Pelteobagrus fulvidraco) | Mixed plant protein: cottonseed protein concentrates and rapeseed protein concentrates (2:3 ratio) | Up to 60% FM replacement by the cottonseed–rapeseed mixture | At 20–40% replacement, growth and liver protein content were maintained, but 60% replacement triggered oxidative stress, inflammatory responses and reduced growth, defining a practical upper limit for this mixture. | Han et al. (2022) |
| Juvenile ballan wrasse (Labrus bergylta) | Soy protein concentrate, pea protein concentrates and other plant proteins in commercialstyle diets | High but partial replacement (majority of dietary protein from plant ingredients) | Plantbased protein ingredients successfully replaced much of the fish meal without negative effects on growth, feed intake or survival when diets were balanced for amino acids and energy. | Cavrois-Rogacki et al. (2022) |
| Multiple carnivorous fish (metaanalysis) | Various plant proteins: soybean meal, soy protein concentrate, pea protein, rapeseed/canola meal, sunflower meal, corn gluten, wheat gluten, etc. | Typically, 30–60% FM replacement across trials | Systematic metaanalysis showed that, across carnivores, moderate fish meal replacement (up to about 50%) by processed plant proteins usually maintains growth and FCR, whereas very high replacement, especially with minimally processed SBM, tends to reduce growth. | Qian et al. (2024) |
Step-by-step summary of cholesterol metabolism enzyme activity and related gene expression
| No. | Step | Key genes/enzymes | Action | Outcome |
|---|---|---|---|---|
| 1 | Activation of sterol regulatory element-binding proteins (SREBPs) | SREBP-2 | Responds to low cholesterol, enhances transcription of biosynthetic genes | Initiates cholesterol production pathways |
| SREBP-1 | Enhances the transcription of genes involved in lipid synthesis. | Drives fatty acid metabolism in response to low intracellular levels. | ||
| 2 | Cholesterol biosynthesis | HMG-CoA reductase (HMGCR) | Converts HMG-CoA to mevalonate, the rate-limiting step in cholesterol synthesis. | Produces cholesterol for cellular membranes and steroid synthesis. |
| Squalene synthase (SQS) | Converts farnesyl pyrophosphate to squalene. | |||
| Lanosterol synthase (LSS) | Converts squalene into lanosterol, a precursor for cholesterol. | |||
| 3 | LDL cholesterol uptake | Low-density lipoprotein receptor (LDLR) | LDLR binds and internalizes LDL particles from the bloodstream, transporting cholesterol into cells. | Supplements intracellular cholesterol levels, especially in low dietary cholesterol scenarios. |
| 4 | Cholesterol regulation | Liver X receptor (LXR) | Activated by high intracellular cholesterol. LXR enhances the transcription of ABCA1 and ABCG5/ABCG8. | Promotes cholesterol efflux and prevents cellular cholesterol overload. |
| 5 | Cholesterol efflux | ATP-binding cassette (ABC) transporters: ABCA1, ABCG5, and ABCG8 | ABCA1: Effluxes cholesterol to form HDL (high-density lipoproteins). | Removes excess intracellular cholesterol and maintains lipid balance. |
| ABCG5/ABCG8: Excretes cholesterol into bile. | ||||
| 6 | Cholesterol storage | Acyl-CoA: cholesterol acyltransferase (ACAT) | Converts free cholesterol into cholesterol esters for storage in lipid droplets. | Prevents excess free cholesterol accumulation in cellular membranes. |
| 7 | Cholesterol catabolism | Cholesterol 7-alpha-hydroxylase (CYP7A1) | Converts cholesterol into bile acids, the first and rate-limiting step of bile acid synthesis. | Facilitates cholesterol excretion and digestion, reducing intracellular cholesterol levels. |
Research indicates that substituting fish meals with plant protein sources generally reduces total cholesterol levels in fish. Liu et al. (2021) conducted a study indicating that substituting fish meals completely with soybean meals in juvenile redlip mullets did not increase total cholesterol levels. A comparable trend was noted in Nile tilapia (Mahmoud et al., 2014), indicating that replacing fish meal with soybean meal did not negatively affect serum cholesterol levels. Yaghoubi et al. (2016) observed an increase in serum total cholesterol in silvery-black porgy (Sparidentex hasta) juveniles when fed diets with higher levels of soybean meal substituting fish meal, suggesting that plant-based diets may affect lipid metabolism differently compared to marine-based diets. Hansen et al. (2007) observed a significant reduction in T-CHO levels after replacing fish meal with plant proteins in Atlantic cod. The research conducted by Shen et al. (2020) also revealed that substituting a fraction of fish meal with cottonseed protein concentrate markedly decreased T-CHO levels in juvenile golden pompano. Research indicates that non-conventional plant proteins, such as sesame oil cake and rapeseed meal, lead to lower cholesterol levels compared to fish meal diets (Dossou et al., 2018; Jahanbakhshi and Hedayati, 2013). The findings indicate that plant proteins may decrease dietary lipid content, leading to reduced blood cholesterol levels. The source of plant protein employed may provide differing effects depending on the fish species and dietary composition. Rahmdel et al. (2018) observed that the partial substitution of fish meal with sunflower meal markedly increased overall cholesterol levels in common carp. Table 3 demonstrates the effects of replacing fish meal with different plant proteins on specific fish species.
Step-by-step summary of cholesterol anabolism
Effect of replacing fish meal with plant protein on serum total cholesterol (T-CHO)
| Species studied | Plant ingredients used | Remarks on total cholesterol | References |
|---|---|---|---|
| Atlantic cod | Plant protein blend | Reduced total cholesterol | Hansen et al. (2007) |
| Great sturgeon | Sesame oil cake | Decreased total cholesterol | Jahanbakhshi and Hedayati (2013) |
| Nile tilapia (Oreochromis niloticus) | Soy protein concentrate | No significant impact on total cholesterol | Mahmoud et al. (2014) |
| Silvery-black porgy (Sparidentex hasta) | Significantly increased total cholesterol, triglyceride and very low density lipoprotein | Yaghoubi et al. (2016) | |
| Tilapia (Oreochromis niloticus × O. aureus) | Rubber seed meal | Reduced total cholesterol with no adverse growth effects | Deng et al. (2017) |
| Turbot (Scophthalmus maximus) | High level of plant protein | T-CHO in plasma significantly reduced | Gu et al. (2017) |
| Largemouth bass (Micropterus salmoides) | Soybean protein concentrate | Significant reduction in T-CHO | Ren et al. (2018) |
| European sea bass | Soybean meal | Decreased total cholesterol | Bonvini et al. (2018) |
| Red sea bream (Pagrus major) | Rapeseed meal | Significant decrease in total cholesterol | Dossou et al. (2018) |
| Common carp | Sunflower meal | Significant increase in total cholesterol | Rahmdel et al. (2018) |
| Black sea bream | Plant protein with taurine supplement | T-CHO lower with all-plant protein diets | Tong et al. (2020) |
| Rainbow trout | Corn gluten meal | Reduced total cholesterol | Staessen et al. (2020) |
| Juvenile golden pompano (Trachinotus ovatus) | Cottonseed protein concentrate | Lowered total cholesterol | Shen et al. (2020) |
| Juvenile redlip mullet | Soybean meal | No adverse impact on serum cholesterol | Liu et al. (2021) |
| Rainbow trout (Oncorhynchus mykiss) | Dephenolized cottonseed protein | Moderately lowered total cholesterol | Zhao et al. (2021 b) |
| Grouper (Epinephelus coioides) | Extruded soybean meal | Reduced total cholesterol | Zhao et al. (2021 a) |
| Rockfish (Sebastes schlegeli) | Various plant protein sources | No significant effect on T-CHO | Kim et al. (2021) |
| Tilapia (Oreochromis niloticus) | Canola meal | Increased total cholesterol | Iqbal et al. (2022 b) |
| Tilapia (Oreochromis niloticus) | Sunflower meal | Reduced cholesterol | Iqbal et al. (2022 a) |
| Gibel carp (Carassius auratus gibelio) | Soybean meal | Decreased total cholesterol | Uyisenga et al. (2023) |
| Juvenile olive flounder (Paralichthys olivaceus) | Plant proteins and meat meal | T-CHO varied from 156.8 to 157.9 mg/dl | Sim et al. (2023) |
| Nile tilapia (Oreochromis niloticus) | Plant protein and animal protein meal | Significant effect on T-CHO | Li et al. (2023) |
| Nile tilapia (Oreochromis niloticus) | Cholesterol and bile acid supplements in plant-based diets | T-CHO lowest in the control group | Jiang et al. (2024 b) |
| Rockfish (Sebastes schlegeli) | Plant proteins with jack mackerel meal | No significant effects on T-CHO | Kim and Cho (2024) |
| Largemouth bass (Micropterus salmoides) | Plant protein sources and mixed animal | T-CHO increased with mixed protein | Chen et al. (2024) |
LDL-C and VLDL-C are critical biomarkers of cardiovascular health in fish, with elevated levels often indicating impaired health or dietary inadequacies. Studies demonstrate that replacing fish meal with plant proteins often leads to reduced levels of LDL-C and VLDL-C. The study by Carneiro et al. (2020) revealed that substituting fish meal with Chlorella sp. meal resulted in reduced LDL-C and VLDL-C levels in zebrafish, suggesting a potential lipid-lowering effect of plant-derived proteins. Liu et al. (2021) revealed a comparable result when soybean meal substituted fish meal in juvenile redlip mullet, resulting in reduced LDL-C and VLDL-C levels. Zhang et al. (2019 a) demonstrated that juvenile rice field eels consuming diets containing soy protein concentrate instead of fish meal exhibited reduced LDL-C levels. Comparable results were observed in sea bass and rainbow trout in the studies conducted by Bonvini et al. (2018) and Lazzarotto et al. (2018), respectively. Studies demonstrate that soybean meal and various plant protein concentrates reduce LDL-C levels in tilapia and carp (Iqbal et al., 2022 b; Ren et al., 2017). The observed reductions are likely attributable to the lower fat and cholesterol content found in plant-based diets in contrast to fish meals, which are inherently high in cholesterol. Most studies indicate that plant-based protein diets can reduce LDL-C levels; however, the extent of reduction may differ depending on the kind and quantity of plant protein consumed. Deng et al. (2017) demonstrated that soy protein significantly decreased LDL-C levels in tilapia, whereas proteins such as rubber seed meal exhibited a less marked impact. Research on very low-density lipoprotein (VLDL-C) is limited compared to other lipid metrics; however, various studies indicate that substituting fish meal with plant proteins has a negligible effect on VLDL-C levels. Peng et al. (2020) demonstrated that replacing soy protein in carp diets resulted in considerable reductions in VLDL-C levels. López et al. (2015) reported related findings in tilapia, indicating that VLDL-C levels remained constant but total cholesterol and LDL-C markedly diminished with the incorporation of soy protein. Lazzarotto et al. (2018) and Palomba et al. (2022) revealed that the partial substitution of fish meal with plant proteins, such as corn gluten meal, led to negligible and insignificant reductions in VLDL-C levels in rainbow trout. Rahmdel et al. (2018) indicated that plant proteins, such as sunflower meal, may lead to a slight spike in VLDLC levels in fish, although the underlying mechanism is not well understood. These researches indicate that while VLDL-C may demonstrate a decreased response to dietary changes relative to other lipid characteristics, it might still experience moderate decreases after the incorporation of fish meal. Further investigation is required to assess the impact of different plant proteins on VLDL-C and its broader consequences for fish health and lipid transport. Table 4 summarizes some research on the effects of replacing fish meal with different plant proteins on LDL-C and VLDL-C levels in specific fish species.
Effect of replacing fish meal with plant protein on serum LDL and VLDL
| Species studied | Plant ingredients used | Effects on LDL | Effects on VLDL | Remarks | References |
|---|---|---|---|---|---|
| Atlantic cod (Gadus morhua L.) | Plant protein mix | Lower LDL levels | – | Stable plasma lipid concentrations | Hansen et al. (2007) |
| Juvenile sturgeon (Huso huso) | Sesame oil cake, corn gluten | Lowered LDL levels | Significant reduction in VLDL | Improved growth performance | Jahanbakhshi and Hedayati (2013) |
| Nile tilapia (Oreochromis niloticus) | Soybean meal | Significant reduction in LDL | – | No significant change in HDL | Mahmoud et al. (2014) |
| Totoaba juveniles (Totoaba macdonaldi) Nile tilapia (Oreochromis niloticus) | Soybean meal | – | VLDL slightly decreased but not statistically significant | – | López et al. (2015) |
| Juvenile gibel carp (Carassius auratus gibelio) | Corn gluten meal | Decreased LDL | – | Reduced plasma cholesterol | Ren et al. (2017) |
| Rainbow trout (Oncorhynchus mykiss) | Plant protein | Reduced LDL levels | Reduced VLDL levels | Reduced serum cholesterol | Lazzarotto et al. (2018) |
| Common carp | Sunflower meal | A moderate reduction in LDL levels | VLDL slightly decreased | Higher replacement levels increased total cholesterol | Rahmdel et al. (2018) |
| Juvenile European sea bass | Soybean meal | Significant decrease in LDL | No significant changes in VLDL | Improved immune response | Bonvini et al. (2018) |
| Red sea bream (Pagrus major) | Rapeseed meal | Moderate reduction in LDL | Significant reduction in VLDL levels | Slight increase in HDL levels | Dossou et al. (2018) |
| Rice field eel (Monopterus albus) | Soy protein concentrate | Reduced LDL levels | Slight decrease in VLDL | Enhanced lipid metabolism | Zhang et al. (2019 a) |
| Hybrid grouper (Epinephelus lanceolatus♂× Epinephelus fuscoguttatus♀) | Plant protein sources | Reduced LDL levels | – | No impact on HDL or growth | Ye et al. (2019) |
| Juvenile golden pompano (Trachinotus ovatus) | Cottonseed protein concentrate | Decreased LDL | – | No negative impact on liver function | Shen et al. (2020) |
| Zebrafish | Chlorella sp. meal | Lower LDL levels | Decreased VLDL | Decreased total cholesterol and triglycerides | Carneiro et al. (2020) |
| Black sea bream (Acanthopagrus schlegelii) | Soybean meal, corn gluten meal | LDL moderately decreased | VLDL moderately reduced | Decreased total cholesterol | Tong et al. (2020) |
| Grass carp (Ctenopharyngodon idellus) | Soy protein concentrate, corn gluten | – | VLDL levels decreased moderately | – | Peng et al. (2020) |
| Juvenile redlip mullet | Soybean meal | Significant decrease in LDL | VLDL levels unchanged | No adverse effects on growth or health | Liu et al. (2021) |
| Rainbow trout | Dephenolization cottonseed protein concentrate | Decreased LDL | Moderate reduction in VLDL | Positive effects on blood lipid profile | Zhao et al. (2021 b) |
| Tilapia (Oreochromis niloticus) | Canola meal | Significant increase in LDL | Moderate increase VLDL levels | Improved feed efficiency | Iqbal et al. (2022 b) |
| Tilapia (Oreochromis niloticus) | Sunflower meal | Moderate decrease in LDL | VLDL levels remained unchanged | Iqbal et al. (2022 a) |
High-density lipoprotein (HDL-C), frequently referred to as “beneficial” cholesterol, plays a crucial role in lipid transport and overall metabolic health. Many researches yield ambiguous results concerning HDL-C levels after replacing fish meal with plant proteins. HDL cholesterol, known for its beneficial attributes, demonstrates varied reactions to the substitution of plant protein in multiple investigations. Zhao et al. (2021 b) demonstrated that incorporating concentrated dephenolization cottonseed protein into the diet of Oncorhynchus mykiss (rainbow trout) did not significantly impact HDL-C levels, although it did reduce LDL and total cholesterol levels. Lazzarotto et al. (2018) and Palomba et al. (2022) conducted research that revealed no significant alterations in HDL-C levels when plant proteins substituted fish meal in the diets of rainbow trout.
Conversely, Shen et al. (2020) reported a notable decrease in HDL-C levels in juvenile golden pompano following the partial substitution of fish meal with cottonseed protein concentrate. Zhao et al. (2021 a) reported a modest reduction in HDL-C levels in hybrid groupers when fish meal was substituted with rendered extruded soybean meal, suggesting that HDL-C responses may vary based on the specific type of plant protein utilized. In certain instances, HDL-C levels were elevated following the substitution of plant protein. Zhang et al. (2019 a) investigated the rice field eel and found that soy protein concentrates elevated HDL-C levels, which is beneficial for the welfare of fish. The differences may be due to species-specific responses to the inclusion of plant proteins or the specific types of plant proteins utilized, as certain proteins contain antinutritional factors that could hinder lipid metabolism. Further research is necessary to elucidate the effects of plant protein-based diets on lipid metabolism and overall fish health. Table 5 presents a summary of research regarding the impact of substituting fish meal with various plant proteins on the HDLC levels in select aquatic species.
Effect of replacing fish meal with plant protein on serum HDL and TG
| Species studied | Plant ingredients used | Effects on HDL | Effects on triglycerides (TG) | References |
|---|---|---|---|---|
| Great sturgeon | Sesame oil cake | Significant increase in HDL | Significant reduction in TG | Jahanbakhshi and Hedayati (2013) |
| Nile tilapia (Oreochromis niloticus) | Soybean meal | HDL levels increased slightly | Triglyceride levels decreased | López et al. (2015) |
| Rainbow trout (Oncorhynchus mykiss) | Corn gluten, soybean meal, wheat gluten and soy protein concentrate | HDL levels increased | TG levels reduced | Lazzarotto et al. (2018) |
| Red sea bream | Rapeseed meal | Significant increase in HDL | Significant reduction in TG | Dossou et al. (2018) |
| Common carp | Sunflower meal | HDL levels unchanged | TG levels slightly reduced | Rahmdel et al. (2018) |
| European sea bass | Soybean meal | Slight improvement in HDL levels | No significant changes in TG | Bonvini et al. (2018) |
| Hybrid grouper | Plant protein sources | Moderate increase in HDL | Ye et al. (2019) | |
| Rice field eel | Soy protein concentrate | HDL levels stable | TG levels moderately decreased | Zhang et al. (2019 a) |
| Hybrid tilapia | Soy protein concentrate, corn gluten | Significant improvement in HDL | TG levels decreased moderately | Peng et al. (2020) |
| Juvenile golden pompano (Trachinotus ovatus) | Cottonseed protein concentrate | HDL levels increased | TG levels decreased | Shen et al. (2020) |
| Black sea bream (Acanthopagrus schlegelii) | Supplementing taurine in plants | HDL levels moderately increased | TG levels decreased moderately | Tong et al. (2020) |
| Juvenile redlip mullet | Soybean meal | No significant change in HDL levels | No significant changes in TG | Liu et al. (2021) |
| Rainbow trout (Oncorhynchus mykiss) | Dephenolized cottonseed protein | Slight increase in HDL | TG levels moderately reduced | Zhao et al. (2021 b) |
| Tilapia (Oreochromis niloticus) | Canola meal | Increased HDL | Increased TG | Iqbal et al. (2022 b) |
| Tilapia (Oreochromis niloticus) | Sunflower meal | Reduced HDL | Reduced TG | Iqbal et al. (2022 a) |
| Tilapia (Oreochromis niloticus) | Corn gluten meal, soybean meal, cottonseed meal and rapeseed meal | Increased TG | Jiang et al. (2024 b) |
Triglycerides (TG) represent a significant lipid parameter that is affected by dietary intake. Multiple studies indicate that substituting fish meal with plant protein correlates with reduced triglyceride levels. Research suggests that plant-based diets often lead to altered lipid profiles in fish, including changes in serum triglyceride (TG) levels. In many cases, when fish meal is replaced with plant protein sources such as soybean meal, corn gluten meal, or rapeseed meal, the TG levels may decrease due to the absence of marine-derived omega-3 fatty acids and cholesterol. Studies by Gatlin III et al. (2007) and Zhou et al. (2017) indicate that these changes may be due to the reduced bioavailability of these essential lipids in plant proteins, which can result in compromised lipid metabolism. Deng et al. (2017) found that substituting fish meal with rubber seed meal in tilapia diets led to a notable decrease in serum triglycerides, suggesting a lipid-lowering impact of plant-based components. This change might be linked to the absence of marine lipids, which are abundant in fish meal, and the possible lipid-modulating properties of certain plant-derived ingredients (Cai et al., 2022; Sarker et al., 2018). López et al. (2015) found that substituting fish meal with soy protein concentrate and taurine in totoaba juveniles did not significantly affect triglyceride levels, indicating that the choice of plant protein and the addition of essential amino acids like taurine can influence blood lipid profiles. Comparable findings were observed in studies involving juvenile blunt snout bream (Ahmed et al., 2019) and juvenile turbot (Nagel et al., 2012). This suggests that the inclusion of essential amino acids such as taurine can mitigate some of the lipid profile changes associated with plant-based feeds. Table 5 presents a summary of research regarding the impact of substituting fish meal with various plant proteins on the triglyceride levels of certain fish species.
Not all plant proteins produce identical impacts on blood cholesterol levels. The substitution of fish meal with plant proteins in fish feed has demonstrated inconsistent impacts on fish blood cholesterol levels. Although numerous research suggests that plant protein diets decrease total cholesterol, LDL-C, and triglycerides, the impact on HDL-C and VLDL-C remains inconsistent. The results are affected by the kind of plant protein employed, the degree of fish meal substitution, and the particular kind of fish examined. Bonvini et al. (2018) discovered that soybean meal generally elevated total cholesterol and LDL in sea bass, whereas Shen et al. (2020) noted decreased cholesterol levels using cottonseed protein concentrate in golden pompano. Zhang et al. (2019 a) observed a beneficial elevation in HDL-C levels when soy protein concentrates substituted fish meal in rice field eel, indicating that plant proteins vary in their capacity to influence cholesterol and lipid metabolism in fish. Furthermore, Hansen et al. (2007) and Mahmoud et al. (2014) emphasized the necessity of equilibrating plant-based protein sources with vital elements like methionine and taurine, which are generally prevalent in fish meal but are possibly lacking in plant proteins. These amino acids are essential for sustaining satisfactory lipid metabolism and averting adverse changes in cholesterol levels.
Muscle cholesterol in fish nutrition corresponds to the cholesterol found in the muscle tissues of fish, which is essential for sustaining cellular functions, supporting membrane integrity, and facilitating muscle contraction and signaling pathways; this is particularly crucial for aquatic animals that depend on robust muscle function for feeding and swimming (O’Keeffe and Gudbrandsen, 2023). Muscle cholesterol levels can be affected by the fish’s diet, specifically the ratio of fish meal to plant-based protein sources (Kim and Cho, 2024). Cholesterol is an essential element of cell membranes, enhancing their stability, fluidity, and permeability. In myocytes, cholesterol preserves the dimensional stability of the cell membrane, enabling efficient cellular activity across varying environmental and metabolic conditions (Meng et al., 2023). Optimal cholesterol levels in muscle tissue enable cells to effectively manage the physical requirements of movement and activity, particularly in swimming fish (Barrientos et al., 2015, 2017).
The cholesterol concentrations in fish muscle tissues are directly correlated with dietary cholesterol consumption (Jiang et al., 2024 a; Li et al., 2023; Meng et al., 2023). Plant-derived protein sources, including soy, rapeseed, and pea meals possess minimal or negligible cholesterol content. Replacing fish meal with these plant proteins reduces the dietary cholesterol available for muscle tissue (Cai et al., 2022; Macusi et al., 2023). Research on fish species, including Atlantic salmon (Salmo salar) (Ytrestøyl et al., 2015) and European sea bass (Dicentrarchus labrax) (Kaushik et al., 2004), has shown that the partial or total replacement of fish meal with plant proteins, such as soybean protein, results in a reduction of muscle cholesterol (Kaushik et al., 2004; Ytrestøyl et al., 2015). A study on gilthead seabream (Sparus aurata) showed that replacing fish meal with plant protein concentrates significantly decreased muscle lipid content, including cholesterol (Couto et al., 2016). Zhang et al. (2020) demonstrated that plant-based meals decreased muscle cholesterol levels in giant yellow croaker (Larimichthys crocea). Furthermore, cholesterol participates in the creation of lipid raft microdomains within the cell membrane that enhance numerous signaling pathways. These rafts are crucial for intercellular interaction among muscle cells and for reacting to external stimuli, including nutrient accessibility or stress (Bagam et al., 2017; Kulkarni et al., 2022; Sviridov et al., 2020). In fish, especially cultured fish, the feed directly impacts muscle composition, including cholesterol levels, which may influence the activity of lipid rafts in muscle cells (Meng et al., 2023; Sissener, 2018). Empirical evidence corroborating this concept is derived from investigations on model organisms, wherein cholesterol deprivation hindered lipid raft formation and compromised cellular signaling (Simons and Sampaio, 2011). Although direct investigations into lipid raft dynamics in fish musculature are scarce, studies on mammals indicate that diminished cholesterol levels result in decreased lipid raft stability and modified cellular communication (Lingwood and Simons, 2010). Considering the evolutionary conservation of cellular systems, it is probable that fish muscle cells undergo analogous perturbations when cholesterol levels are diminished as a result of plant protein diets.
In response, fish may need to augment endogenous cholesterol production in muscle tissues or improve cholesterol uptake from circulation through lipoproteins such as low-density lipoproteins. In fish fed plant-based diets, muscle cells may enhance the expression of enzymes such as HMG-CoA reductase (HMGCR), which plays a role in cholesterol biosynthesis, to internally synthesize more cholesterol (Li et al., 2023; O’Keeffe and Gudbrandsen, 2023). Plant-based diets frequently include phytosterols, which may hinder the absorption of cholesterol in the intestine. Phytosterols compete with cholesterol, diminishing the total cholesterol absorbed from the diet and conveyed to muscle tissue (Poli et al., 2021; Racette et al., 2010). This may require modifications in cholesterol metabolism inside the muscle to sustain optimal cellular function. Additionally, muscle cells have the capacity to accumulate cholesterol as cholesterol esters. The enzyme acyl-CoA (cholesterol acyltransferase) facilitates the conversion of free cholesterol into a storable form, hence regulating excess cholesterol. Cholesterol reserves can be utilized when necessary for membrane repair or other cellular processes. Moreover, plant-based diets for fish must adjust to the reduced availability of dietary cholesterol. They may augment cholesterol production or elevate the expression of cholesterol transport proteins, such as low-density lipoprotein receptor (LDLR), to guarantee adequate cholesterol delivery to muscle tissues. Bou et al. (2017) discovered that Atlantic salmon consuming soy protein concentrate had a compensatory enhancement in cholesterol production pathways within muscle tissue, attributable to the reduced dietary cholesterol levels. This signifies the muscle’s capacity to adjust to dietary modifications to sustain cholesterol levels essential for cellular activity. Geay et al. (2011) revealed that muscle tissue in European sea bass (Dicentrarchus labrax) consuming plant protein-based diets (having low-cholesterol) had elevated expression of genes associated with cholesterol production, including genes in relation to LC-PUFA and cholesterol biosynthetic pathways (HMGCR). This adaptive response enabled the fish to sustain cholesterol levels in muscle despite diminished food consumption. A study conducted by (Caballero-Solares et al., 2017) demonstrated that substituting fish meal with plant proteins in the diets of European sea bass resulted in elevated cholesterol accumulation in muscular tissues. This was demonstrated by increased LDLR gene expression, signifying improved uptake of circulating cholesterol to offset the reduced cholesterol levels in the plant-based diet.
The enzymatic activity and the gene expression of cholesterol metabolism in fish nutrition elucidates the biochemical processes related to the production, transport, usage, and regulation of cholesterol in fish (Castro et al., 2015; Martini and Pallottini, 2007). Table 2 shows the step-by-step summary of cholesterol metabolism enzyme activity and related gene expression. Cholesterol is an essential element in fish, serving critical functions in cell membrane architecture, hormone synthesis, and bile acid generation. The enzymes and genes implicated in cholesterol metabolism regulate cholesterol levels, adapting to food intake and physiological requirements. Substituting fish meal with plant protein sources markedly influences the expression of genes associated with cholesterol metabolism (Kim and Cho, 2024; Mugwanya et al., 2023).The expression of particular genes, including HMG-CoA reductase (HMGCR), low-density lipoprotein receptor (LDLR) (Martini and Pallottini, 2007), ATP-binding cassette transporters (ABC family), sterol regulatory element-binding proteins (SREBPs), and liver X receptor (LXR), is considerably influenced. The genes and enzymes associated with cholesterol production, transport, and control are expressed in reaction to dietary variables, such as the consumption of proteins, fats, oils, and other nutrients. In fish, dietary transitions from fish meal to plant protein sources markedly affect the expression of genes and enzymes activities associated with cholesterol metabolism. As these diets frequently lack adequate cholesterol and essential fatty acids, fish typically regulate the expression of these genes and the activities of these enzymes as a compensatory mechanism to enhance endogenous cholesterol homeostasis.
SREBPs are critical transcription factors that regulate the expression of genes involved in lipid and cholesterol metabolism. SREBPs exist in two main isoforms: SREBP-1, which primarily controls fatty acid and triglyceride synthesis, and SREBP-2, which regulates cholesterol biosynthesis and uptake. Replacing fish meal with plant protein sources typically leads to increased expression of SREBPs, particularly SREBP-2, as fish attempt to compensate for the lower dietary cholesterol provided by plant proteins. This upregulation activates genes involved in cholesterol biosynthesis, such as HMG-CoA reductase (HMGCR), to maintain cholesterol homeostasis. Bou et al. (2017) found that replacing fish meal with soy protein concentrate in Atlantic salmon (Salmo salar) diets led to the upregulation of SREBP-2 expression. This upregulation was linked to the lower dietary cholesterol in plant-based diets, which triggered the activation of cholesterol biosynthesis genes, including HMGCR.
In a study on Atlantic salmon (Salmo salar), Caballero-Solares et al. (2018) demonstrated that diets containing plant protein induced an increase in SREBP-2 activity. Zhu et al. (2020) reported that feeding plant protein to rainbow trout (Oncorhynchus mykiss) resulted in an increased expression of SREBP-2 and its target genes, thereby increasing SREBP-2 enzyme activity. Mellery et al. (2015) demonstrated that replacing fish meal with plant protein sources, including soy protein and rapeseed protein, in rainbow trout diets significantly increased SREBP-2 expression in the liver. Liland (2011) showed replacing fish meal with plant proteins and vegetables oils in Atlantic salmon (Salmo salar) diets led to increased expression of SREBP-2 and its downstream genes, including HMGCR. Much research with similar results is shown in Table 6. This was interpreted as a compensatory mechanism to cope with the lower dietary cholesterol provided by plant proteins (Caballero-Solares et al., 2018; Liland, 2011; Mellery et al., 2015; Zhu et al., 2020).
Effect of replacing fish meal with plant protein on squalene synthase and lanosterol synthase
| Species studied | Plant ingredients used | Effects on squalene synthase (SQS) | Effects on lanosterol synthase (LSS) | References |
|---|---|---|---|---|
| Rainbow trout (Oncorhynchus mykiss) | Soybean meal | Upregulation of SQS genes and increased enzyme activity | Increased LSS expression and enzyme activity | Turchini and Francis (2009) |
| Atlantic salmon (Salmo salar L.) | Vegetable proteins | Increased SQS gene expression and enzyme activity | Increased LSS gene expression and enzyme activity | Bendiksen et al. (2011) |
| European sea bass (Dicentrarchus labrax) | Rapeseed and sunflower meal | Increased SQS gene expression and activity | Upregulated LSS expression to compensate for low dietary cholesterol | Guerreiro et al. (2015) |
| Atlantic salmon (Salmo salar) | Soy protein concentrate | Increased SQS expression | Increased LSS expression and enzyme activity | Ytrestøyl et al. (2015) |
| Rainbow trout (Oncorhynchus mykiss) | Plant protein sources | Upregulated SQS activity in response to low cholesterol | Enhanced LSS expression due to reduced dietary cholesterol | Zhu et al. (2018) |
| Atlantic salmon (Salmo salar) | Pea, soy and wheat | Increased SQS gene expression and enzyme activity | increased enzyme activity | Dhanasiri et al. (2020) |
| Nile tilapia (Oreochromis niloticus) | Soybean meal and pea protein | Increased SQS expression | Increased LSS expression | El-Sayed et al. (2021) |
| Rainbow trout (Oncorhynchus mykiss) | Cottonseed protein concentrate | Increased SQS expression | Enhanced LSS expression | Liu et al. (2022) |
| Common carp (Cyprinus carpio) | Cottonseed protein concentrate | Increased SQS expression due to low dietary cholesterol | Enhanced LSS expression | Fan et al. (2024) |
While SREBP-2 primarily regulates cholesterol metabolism, SREBP-1 plays a crucial role in controlling fatty acid synthesis. Plant-based diets, which are often deficient in essential fatty acids found in fish meal, can lead to the upregulation of SREBP-1 as fish attempt to synthesize the necessary fatty acids endogenously. In a study by Morais et al. (2011), Atlantic salmon fed plant protein-based diets showed increased SREBP-1 expression, leading to higher rates of fatty acid synthesis. This was linked to the lower availability of omega-3 fatty acids in plant-based diets, which prompted the fish to up-regulate SREBP-1 to synthesize fatty acids. In Atlantic salmon, diets containing protein-based diets resulted in increased SREBP-1 expression, driving the synthesis of fatty acids to compensate for the lower lipid content of plant-based diets (Caballero-Solares et al., 2017).
SREBPs are regulated by a feedback mechanism based on intracellular cholesterol and fatty acid levels. When dietary cholesterol is low, as in plant-based diets, SREBP-2 is cleaved and translocated to the nucleus, where it activates genes involved in cholesterol biosynthesis, such as HMGCR (Kortner et al., 2013; Ranasinghe et al., 2022). Similarly, when fatty acids are deficient, SREBP-1 is activated to enhance fatty acid synthesis. Phytosterols, present in many plant proteins, also affect SREBP regulation. These sterols compete with cholesterol for absorption, further reducing cholesterol availability and enhancing the activation of SREBP-2. Summary of research regarding the impact of substituting fish meal with various plant proteins on SREBPs of certain fish species is shown in Table 8.
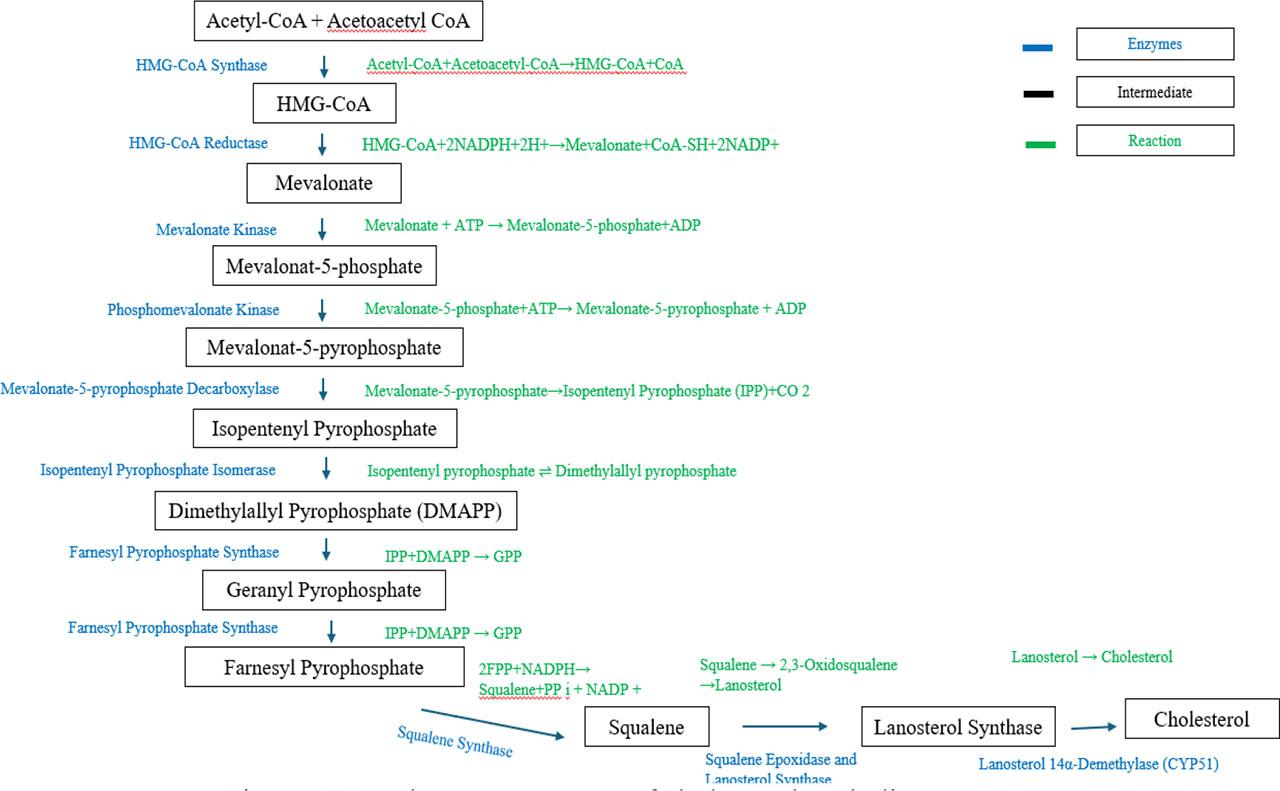
SQS, an essential enzyme in cholesterol biosynthesis, has been the focus of several studies examining how plant protein-based diets affect fish cholesterol metabolism and overall health. SQS catalyzes the conversion of farnesyl pyrophosphate into squalene, the first committed step in the sterol biosynthesis pathway. This leads to the production of squalene, a precursor to all sterols, including cholesterol. The pathway continues from squalene, eventually leading to the production of lanosterol, which is then converted into cholesterol through a series of further enzymatic reactions. Lanosterol synthase (LSS), also known as oxidosqualene cyclase (OSC), is responsible for the cyclization of 2,3-oxidosqualene (the oxygenated form of squalene) into lanosterol, a critical intermediate in the multi-step process that ultimately leads to the synthesis of cholesterol. This cyclization reaction is a complex transformation where the linear squalene molecule undergoes structural reorganization into the four-ring structure characteristic of sterols like cholesterol. The production of lanosterol marks the first step in the sterol branch of the mevalonate pathway, which is crucial for cholesterol production. Once lanosterol is synthesized, it undergoes multiple enzymatic modifications, including demethylation and saturation reactions, leading to the formation of cholesterol.
SQS and LSS function downstream of HMG-CoA reductase, which is responsible for producing farnesyl pyrophosphate, the substrate for SQS. Since SQS catalyzes a rate-limiting step in sterol synthesis, it is tightly regulated. When cellular cholesterol levels are high, the activity of SQS is downregulated, thereby reducing cholesterol synthesis. Conversely, under conditions of low cholesterol (for instance when fish are fed plant-based diets with low cholesterol content), SQS expression can increase, promoting endogenous cholesterol production to compensate for reduced dietary intake.
In a study by Turchini and Francis (2009) on rainbow trout (Oncorhynchus mykiss), the replacement of fish meal with soybean meal resulted in a significant reduction in dietary cholesterol intake. This led to an increase in the expression of genes involved in cholesterol synthesis, including SQS and LSS. Similarly, Amritha et al. (2020) observed an upregulation of SQS and LSS in Atlantic salmon (Salmo salar) fed a wheat gluten-based diet. Bendiksen et al. (2011) studied the effects of replacing fish meal with lupin protein concentrate in European sea bass (Dicentrarchus labrax) and reported similar findings. Kemski (2018) explored the expression of SQS and LSS in yellow perch (Perca flavescens), comparing fish meal-based diets with soybean meal diets but observed fish on plant-based diets had significantly elevated SQS and LSS gene expressions. Kemski (2018) and Turchini and Francis (2009) noted that while the fish exhibited upregulated SQS and LSS expressions, they also showed impaired growth and liver health, indicating that endogenous cholesterol biosynthesis might not fully compensate for the lack of dietary cholesterol. Amritha et al. (2020) and Bendiksen et al. (2011) also revealed that despite the increase in SQS and LSS expression, the fish demonstrated reduced cholesterol levels and hepatic fat accumulation. This suggested that while SQS and LSS activities were elevated in response to the low cholesterol content in the diet, the overall lipid metabolism was disrupted, leading to suboptimal health outcomes. This highlights a limitation in the ability of plant-based diets to support cholesterol balance through endogenous synthesis alone.
One of the unique challenges posed by plant-based diets is the presence of phytosterols, which are structurally similar to cholesterol and can compete with cholesterol for absorption in the intestine. Guerreiro et al. (2015) and Sissener et al. (2017) conducted a study on European sea bass and Atlantic salmon, respectively. They observed that when fish meal was replaced with plant protein sources rich in phytosterols, there was reduction in the absorption of cholesterol which led to a compensatory increase in SQS and LSS expressions. However, despite the upregulation of SQS and LSS, cholesterol levels in the fish remained lower than those fed fish meal-based diets, indicating the limitations of endogenous synthesis in maintaining cholesterol homeostasis in the presence of phytosterols. Sissener et al. (2017) provided further insights into the interaction where there were decreased plasma cholesterol levels. Several studies have explored strategies to mitigate the negative effects of plant-based diets on cholesterol metabolism. Yao et al. (2021) investigated the addition of cholesterol supplements and essential fatty acids to plant-based diets in common carp (Cyprinus carpio). They found that supplementing the diet with bile acids significantly reduced the compensatory upregulation of SQS and LSS, and improved overall lipid metabolism and growth performance. This suggests that while plant-based diets lead to increased SQS and LSS expression, appropriate supplementation can help restore balance and reduce the need for endogenous cholesterol synthesis. Summary of research regarding the impact of substituting fish meal with some plant proteins on SQS and LSS of certain fish species are shown in Table 6.
HMGCR is a key enzyme in the mevalonate pathway for cholesterol biosynthesis. HMGCR is the rate-limiting enzyme in cholesterol biosynthesis, and its expression is influenced by dietary cholesterol levels, which are typically lower in plant-based diets than in fish meals. These upregulations activate genes such as low-density lipoprotein receptor (LDLR), liver X receptor (LXR), and ATP-binding cassette transporters (ABC family). Zhu et al. (2018) investigated the impact of replacing fish meal with soy protein concentrate in rainbow trout (Oncorhynchus mykiss) diets. The study found an increase in HMGCR expression in the liver, suggesting enhanced cholesterol biosynthesis. In a study on Nile tilapia, researchers explored the effects of replacing fish meal with soybean meal and corn gluten meal. Liland (2011) found that HMGCR mRNA expression was significantly higher in fish fed plant-based diets compared to those fed traditional fish meal-based diets. A study conducted by Mellery et al. (2015) replaced fish meal with a blend of soy protein and rapeseed protein concentrate in rainbow trout diets. The results demonstrated a significant upregulation of HMGCR expression in the liver tissues of fish fed plant-based diets. The up-regulation of HMGCR was directly linked to the need for increased endogenous cholesterol synthesis, compensating for the reduced cholesterol intake from plant-based diets to maintain homeostatic cholesterol levels (Caballero-Solares et al., 2018). The increase in HMGCR expression in response to plant-based diets can be attributed to several mechanisms. One of the primary regulatory pathways is through sterol regulatory element-binding proteins (SREBPs), particularly SREBP-2, which regulates cholesterol biosynthesis. When dietary cholesterol is low, as in plant-based diets, SREBP-2 is activated and induces the expression of HMGCR to increase cholesterol synthesis (Bou et al., 2017; Zhu et al., 2020). Additionally, the presence of phytosterols in plant-based diets, which compete with cholesterol for absorption, may further exacerbate the need for endogenous cholesterol synthesis. Summary of research on the effect of substituting fish meal with some plant proteins on HMGCR expression of some fish species is shown in Table 7.
Effect of replacing fish meal with plant protein on HMG-CoA reductase
| Species studied | Plant ingredients used | HMGCR remarks | References |
|---|---|---|---|
| Yellowtail (Seriola quinqueradiata) | Soy protein concentrate | Elevated HMGCR expression as an adaptive response to plant-based protein diets | Maita et al. (2006) |
| European sea bass (Dicentrarchus labrax) | Soybean meal, wheat gluten and white sweet lupin | High stimulation of HMGCR genes due to cholesterol biosynthesis | Geay et al. (2011) |
| Atlantic salmon (Salmo salar) | Pea protein, soy protein | Increased expression of HMGCR due to reduced dietary cholesterol | Kortner et al. (2013) |
| Atlantic salmon (Salmo salar L.) | Lupin meal and wheat gluten | Capacity for cholesterol synthesis was up-regulated simultaneously with the mRNA expression of HMGCR | Gu et al. (2014) |
| Turbot (Scophthalmus maximus) | Soybean meal and wheat gluten | Peaked mRNA expression of HMGCR in the intestine | Gu et al. (2017) |
| Atlantic salmon (Salmo salar) | Soy protein concentrate | Significant increase in HMGCR expression as a compensatory mechanism due to low dietary cholesterol | Bou et al. (2017) |
| Rainbow trout (Oncorhynchus mykiss) | Corn gluten, soybean meal, wheat gluten and white lupin, peas and rapeseed meal | Increased hepatic mRNA expression of HMGCR in both juveniles and ongrowing fish. | Lazzarotto et al. (2018) |
| Atlantic salmon (Salmo salar) | Rapeseed meal, soy protein | Higher HMGCR expression was observed as cholesterol synthesis compensation mechanism. | Caballero-Solares et al. (2018) |
| Japanese seabass (Lateolabrax japonicus) | Soy protein concentrate | HMGCR expression was significantly elevated with plant protein diets replacing fish meal, indicating increased cholesterol synthesis. | Zhang et al. (2019 b) |
| Rainbow trout (Oncorhynchus mykiss) | Corn gluten, wheat gluten, soybean meal, soybean protein concentrates light white lupin, and dehulled pea | Hepatic mRNA expression of HMGCR increased, suggesting a promotion of the synthesis of cholesterol regulated by SREBP-2 | Zhu et al. (2020) |
| Common carp (Cyprinus carpio L.) | Soybean meal and cottonseed protein concentrate | Peaked the liver mRNA expression of HMGCR | Yao et al. (2021) |
| Largemouth bass (Micropterus salmoides) | Partial inclusion of soybean protein concentrate | Enhanced the mRNA levels of HMGCR | Chen et al. (2024) |
| Red seabream (Pagrus major) | Soybean meal, corn gluten, soy protein concentrates | Elevated expression of HMGCR in the liver | Murashita et al. (2024) |
| Largemouth bass (Micropterus salmoides) | Soybean, rapeseed, and cottonseed protein | Increased expression of HMGCR due to enhanced cholesterol synthesis and efflux | Yao et al. (2024) |
| Nile tilapia (Oreochromis niloticus) | Corn gluten, soybean meal, cottonseed meal, and rapeseed meal | The plant protein peaked the hepatic mRNA expression of HMGCR, however, it was suppressed by cholesterol and bile acid supplementation | Jiang et al. (2024 b) |
LDLR is a critical component of cholesterol homeostasis, and it is responsible for mediating the uptake of cholesterol-containing lipoproteins from the bloodstream into cells. Replacing fish meal with plant protein sources generally results in an upregulation of LDLR expression in fish. Cai et al. (2022) found that gibel carp (Carassius auratus gibelio) diets containing plant proteins exhibited marked upregulation of LDLR expression. This increase in LDLR activity helped maintain cholesterol homeostasis despite the lower dietary cholesterol input. Murashita et al. (2024) replaced fish meal with plant proteins in red sea bream (Pagrus major) diets and observed a significant increase in LDLR expression. Furthermore, in a study on red sea bream, fish fed diets with increasing amounts of fermented rapeseed meal had significantly elevated LDLR mRNA expression (Dossou et al., 2018). All these studies agree with other research in Table 6.
The upregulation of LDLR in fish fed plant-based diets is largely driven by a need to balance cholesterol homeostasis. Elevated LDLR mRNA expression indicates a physiological response to the lower cholesterol levels in the diet (Ye et al., 2019). Low dietary cholesterol levels reduce the feedback inhibition on LDL receptor expression, leading to increased LDLR activity to boost cholesterol uptake (Bou et al., 2017). This is often accompanied by changes in other genes involved in lipid metabolism, such as HMG-CoA reductase (HMGCR), which is upregulated to synthesize cholesterol endogenously (Caballero-Solares et al., 2018; Hussain et al., 2024; Ye et al., 2019). Moreover, the presence of phytosterols further reduces cholesterol availability, triggering a compensatory upregulation of LDLR to increase the efficiency of cholesterol transport from the bloodstream (Ye et al., 2019). Table 8 gives a summary of research on the effect of substituting fish meal with some plant proteins on LDLR of some fish species.
Effect of replacing fish meal with plant protein on LDLR, ABC family, SREBP, and LXR expressions
| Species studied | Plant ingredients used | LDLR | ABC family transporter expression | SREBP expression | LXR | References |
|---|---|---|---|---|---|---|
| Atlantic salmon (Salmo salar) | Plant proteins | LDLR mRNA expression was significantly elevated in fish fed plant protein diets. | Significant upregulation of ABCG5 and ABCG8 in fish fed plant protein diets. | Increased expression of SREBP-2 and its downstream genes, including HMGCR. | Upregulation of LXR expression to promote cholesterol efflux. | Liland (2011) |
| Gilthead sea bream (Sparus aurata) | Soy protein concentrate | Significant increase in LDLR expression. | Significant increase in ABCA1 expression. | Increased expression of SREBP-2 and its target genes. | Increased LXR expression linked to cholesterol homeostasis. | Couto et al. (2014) |
| Rainbow trout (Oncorhynchus mykiss) | Soy protein and rapeseed protein concentrate | Upregulation of LDLR as a compensatory mechanism to lower cholesterol intake. | Upregulation of ABCA1 and ABCG5/G8 expression in response to plant proteins. | Upregulation of SREBP-2 linked to cholesterol biosynthesis in response to plant proteins. | Upregulation of LXR expression to compensate for reduced dietary cholesterol intake. | Mellery et al. (2015) |
| Atlantic salmon (Salmo salar) | Soy protein concentrate | Elevated LDLR expression due to reduced dietary cholesterol availability. | Increased ABCA1 expression in response to plant-based diets. | Upregulation of SREBP-2 expression in response to reduced dietary cholesterol. | Upregulation of LXR expression in response to the lower cholesterol content of the diet. | Bou et al. (2017) |
| Tilapia (Oreochromis niloticus × Oreochromis aureus) | Rubber seed meal | Decreased LDLR expression | Deng et al. (2017) | |||
| Rainbow trout (Oncorhynchus mykiss) | Totally plant-based diet | Lower ABCG8 expression | Increased SREBP-2 expression | Reduced LXRa activity | Zhu et al. (2018) | |
| Atlantic salmon (Salmo salar) | Soy protein concentrate, rapeseed meal | Higher LDLR expression observed to support cholesterol uptake in low-cholesterol diets. | Higher ABCA1 expression due to the lower cholesterol content of plant-based diets. | Increased SREBP-1 expression driving fatty acid synthesis due to lower lipid content. | Increased LXR activity leading to enhanced cholesterol removal via efflux transporters. | Caballero-Solares et al. (2018) |
| Japanese seabass (Lateolabrax japonicas) | Soy protein, maize gluten, wheat gluten | Significant increase in LDLR activity to compensate for low dietary cholesterol. | Increased expression of ABCG5 and ABCG8 as a compensatory response. | Increased SREBP-2 expression driving cholesterol and fatty acid synthesis. | Increased LXR expression to maintain cholesterol and fatty acid metabolism. | Zhang et al. (2019 b) |
| Rainbow trout (Oncorhynchus mykiss) | Plant protein sources | The expression of LDLR was significantly increased | ABCA1, ABCG5 and ABCG8 showed elevated expression levels | SREBP-2 was upregulated | LXR showed increased expression | Zhu et al. (2020) |
| Tilapia (Oreochromis niloticus) | Corn gluten meal, soybean meal, cottonseed meal and rapeseed meal | Significant upregulation of LDLR expression in fish fed plant protein diets | Significant increase in ABCG5 expression | Significant increase in ABCG5 expression | Increased LXR expression in response to plant protein | Jiang et al. (2024 b) |
LXR is a nuclear receptor that plays a central role in regulating cholesterol, lipid metabolism, and inflammatory responses in fish. LXRs regulate the expression of genes involved in cholesterol efflux, such as ATP-binding cassette transporters (ABC family) and fatty acid synthesis genes. LXR regulates cholesterol homeostasis by promoting cholesterol efflux through the upregulation of transporters like ABCA1, ABCG5, and ABCG8, which are responsible for removing excess cholesterol from cells. LXR also regulates lipogenesis by activating genes involved in fatty acid synthesis, including SREBP-1. Given that plant-based diets tend to be lower in cholesterol and higher in phytosterols compared to fish meals, LXR activity is often modulated in response to these dietary changes. Replacing fish meal with plant protein sources can alter the expression of LXR and its target genes due to differences in the sterol and fatty acid content of plant proteins. Fish adapt to these changes by modulating LXR activity to maintain lipid homeostasis. Takakuwa et al. (2023) found that replacing fish meal with plant protein concentrate in red sea bream diets led to an increase in LXR expression. This was accompanied by higher expression of cholesterol transporters, indicating an enhanced cholesterol efflux in response to the reduced cholesterol content of the plant-based diet. Zhu et al. (2020) reported that rainbow trout fed plant protein exhibited increased LXR expression. This upregulation was linked to enhanced cholesterol efflux through the activation of ABCA1 and ABCG5, as the fish adapted to the lower cholesterol levels in the plant-based diet. In a study by Caballero-Solares et al. (2018), Atlantic salmon fed diets containing terrestrial alternatives to fish meal (plant protein) showed increased LXR activity. The upregulation of LXR in these fish was linked to enhanced expression of ABCA1 and other transporters involved in cholesterol removal, suggesting a compensatory mechanism in response to the low cholesterol content of plant-based diets. Bou et al. (2017) investigated the effects of replacing fish meal with soy protein concentrate on LXR expression in Atlantic salmon. The study found that LXR expression was upregulated. This up-regulation led to increased expression of cholesterol efflux transporters, including ABCA1 and ABCG5. Liang et al. (2024) studied the effects of soy protein concentrate-based diets on LXR expression in largemouth bass. The fish exhibited increased LXR expression, which promoted the upregulation of genes involved in cholesterol efflux, including ABCG5 and ABCG8. This response was associated with the higher phytosterol content of the plant-based diet, which competes with cholesterol for absorption, triggering an adaptive response via LXR. In Atlantic salmon, Morais et al. (2011) found that feeding plant-based diets led to increased LXR expression, which activated SREBP-1 and promoted fatty acid synthesis. Table 8 gives a summary of research on the effect of substituting fish meal with some plant proteins on LXR of some fish species.
ABC transporters are a large family of proteins involved in the transport of various molecules across cellular membranes, including lipids, sterols, and xenobiotics. These transporters, especially ABCA1, ABCG1, ABCG5, and ABCG8, play a critical role in cholesterol homeostasis by mediating cholesterol efflux from cells, thus maintaining lipid balance in fish.
ABCA1 and ABCG1 is one of the key transporters involved in the export of cholesterol and phospholipids to lipid-poor apolipoproteins, facilitating the formation of high-density lipoproteins (HDL). This process is crucial for the reverse cholesterol transport pathway, where excess cholesterol is removed from tissues and transported to the liver for excretion. ABCA1 helps prevent cholesterol accumulation in cells, aligning with catabolic processes that reduce cholesterol levels. ABCG1 works alongside ABCA1 to export cholesterol to HDL particles. It also contributes to reverse cholesterol transport, ensuring cholesterol is cleared from peripheral tissues and ultimately eliminated. In fish fed plant-based diets, ABCA1 expression tends to be up-regulated to compensate for the lower dietary cholesterol intake. Bou et al. (2017) reported that replacing fish meal with soy protein concentrate led to an increase in ABCA1 expression in Atlantic salmon. This upregulation was attributed to the reduced availability of dietary cholesterol, which prompted the fish to increase cholesterol efflux to maintain cellular lipid balance. In a study by Caballero-Solares et al. (2018), Atlantic salmon (Salmo salar) fed diets containing plant proteins exhibited a marked increase in ABCA1 expression.
The ABCG5 and ABCG8 transporters are expressed in the liver and intestines and they are involved in the biliary excretion of cholesterol, a crucial step in cholesterol catabolism (Plosch et al., 2006). The ABCG5 and ABCG8 transporters form a heterodimer that plays a key role in transporting cholesterol and phytosterols across the cellular membrane (Dikkers and Tietge, 2010). These transporters help regulate the excretion of sterols into bile and are particularly important in fish fed plant-based diets due to the higher levels of phytosterols in plant ingredients (Dikkers and Tietge, 2010; Wang et al., 2015). Mellery et al. (2015) showed that rainbow trout fed diets containing soy protein and rapeseed protein exhibited an increase in ABCG5 and ABCG8 expression. Moreover, Liang et al. (2024) found that largemouth bass (Micropterus salmoides) fed diets with soy protein concentrate meal exhibited increased expression of ABCG5 and ABCG8 in the liver. In all these researches, the upregulation was attributed to the higher levels of phytosterols in the plant-based diet, which necessitated the enhanced excretion of excess sterols to maintain lipid balance (Gu et al., 2014; Kopylov et al., 2021; Lifsey et al., 2020; Sissener et al., 2018; Takase and Ushio, 2018; Zhu et al., 2018). Plant-based diets, especially those containing ingredients such as soy, rapeseed, and corn gluten, tend to have higher levels of phytosterols, which can influence the expression of ABC transporters involved in cholesterol and sterol transport (Couto et al., 2014; Gu et al., 2014, 2017; Takakuwa et al., 2023). Phytosterols compete with cholesterol for absorption in the intestine, reducing cholesterol uptake and increasing the need for cholesterol efflux through ABCA1. This leads to the up-regulation of ABCA1 to maintain cellular cholesterol levels. Furthermore, the higher levels of phytosterols in plant-based diets stimulate the expression of ABCG5 and ABCG8, which are responsible for excreting excess phytosterols and cholesterol. This mechanism helps fish maintain lipid homeostasis by preventing the accumulation of excess sterols in the body. Summary of research on the effect of substituting fish meal with some plant proteins on ABC transporters of some fish species is shown in Table 8.
ACAT is responsible for esterifying free cholesterol into cholesterol esters for storage within cells. This enzyme helps regulate intracellular cholesterol levels by storing excess cholesterol in the form of esters. ACAT activity is elevated when fish consume diets rich in cholesterol. This enzyme helps manage excess cholesterol by converting it into a storage form, preventing an overload of free cholesterol in cells. Several studies, such as those by Liang et al. (2024), highlight that plant-based diets rich in soybean or corn gluten led to increased ACAT expression. The rise in ACAT activity is an adaptive mechanism to store the excess free cholesterol as esters, preventing intracellular cholesterol overload. This was observed in fish like juvenile largemouth bass and gilthead sea bream. Fish fed plant proteins need to convert more cholesterol into esters, which reflects an attempt to maintain cellular cholesterol balance (Jiang et al., 2024 b; Valente et al., 2011). However, this increased activity might also indicate lipid storage issues and the onset of fatty liver over time (Katan et al., 2019; Liland, 2011). Studies like those by Deng et al. (2013), Tong et al. (2020) and Koven et al. (2016, 2023) show that cholesterol supplementation improves ACAT activity in fish fed plant proteins, such as soy protein concentrate. The addition of cholesterol helps balance the lipid metabolism disrupted by plant-based diets, ensuring proper lipid storage without excessive sterol accumulation. Cholesterol supplementation can mitigate the negative impacts of plant proteins on ACAT, stabilizing cholesterol esterification and bile acid synthesis. This strategy also reduces metabolic stress, especially in fish species heavily dependent on dietary cholesterol. Research by He et al. (2022) and Takase and Ushio (2018) emphasizes that diets containing phytosterols (naturally present in plant-based diets) suppress ACAT expression. Phytosterols mimic cholesterol, competing with it for metabolism but do not activate ACAT efficiently. This can impair the esterification process, leading to the accumulation of free cholesterol in tissues (He et al., 2022). Some studies, such as Liang et al. (2024) and Yuan et al. (2020), suggest that plant oils cause only moderate changes in ACAT activity. These oils, when combined with plant proteins, maintain lipid metabolism without triggering significant sterol accumulation or suppressing ACAT function. Diets with balanced lipid sources (for example, plant oils mixed with fish oils) reduce the metabolic burden on ACAT and ensure efficient cholesterol regulation without severe disruptions in metabolism (Ge et al., 2023). Studies like Xu et al. (2020) report that taurine supplementation alongside plant proteins helps normalize ACAT activity. Taurine plays a role in bile acid synthesis, improving cholesterol metabolism and alleviating the metabolic stress caused by plant protein diets. Fish species, such as Japanese seabass, benefited from taurine supplementation, maintaining both lipid balance and bile acid synthesis. This strategy shows promise for optimizing plant-based diets in aquaculture. Table 9 presents a summary of research on the effect of substituting fish meal with some plant proteins on ACAT activity in some fish species.
Effect of replacing fish meal with plant protein on ACAT activity
| Species studied | Plant ingredients used | Effects on ACAT | References |
|---|---|---|---|
| Rainbow trout (Oncorhynchus mykiss) | Soybean meal, soy protein concentrate | Improved ACAT activity with dietary cholesterol supplementation | Deng et al. (2013) |
| Atlantic salmon | Microalgae and plant proteins | Partial restoration of ACAT activity | Kousoulaki et al. (2016) |
| Hybrid tilapia (Oreochromis niloticus × Oreochromis aureus) | Rubber seed meal | Elevated ACAT activity for lipid storage | Deng et al. (2017) |
| Tiger puffer (Takifugu rubripes) | Soy protein isolate, wheat gluten | Elevated ACAT activity with taurine addition | Xu et al. (2020) |
| Hybrid grouper | Soy protein concentrate | ACAT upregulation under plant protein diets | Chen et al. (2020) |
| Nile tilapia | Corn gluten, soybean concentrate | Increased ACAT expression and lipid storage | Li et al. (2023) |
This enzyme catalyzes the first step in the conversion of cholesterol into bile acids in the liver. It plays a critical role in regulating cholesterol levels by facilitating the excretion of cholesterol as bile acids (Li and Chiang, 2009). CYP7A1 activity is influenced by the amount of dietary cholesterol. Higher dietary cholesterol can increase the need for bile acid production, leading to higher activity of CYP7A1 (Caponio et al., 2020). This enzyme helps balance excess dietary cholesterol by converting it to bile acids for excretion (Just et al., 2018).
Several studies, including those by Zhang et al. (2019 b) on Japanese seabass and Zhao et al. (2020) on common carp, report that plant-based diets tend to reduce CYP7A1 expression. This decrease in enzyme activity corresponds to lower bile acid synthesis, which may compromise the ability to metabolize and excrete excess cholesterol. Moreover, accumulation of cholesterol in hepatic tissues potentially leads to metabolic disorders like fatty liver in some species. This suggests that fish meal contains components such as certain amino acids, cholesterol, or other nutrients, that are crucial for maintaining optimal bile acid metabolism. Kortner et al. (2014) found that supplementing plant-based diets with dietary cholesterol helped restore CYP7A1 activity. In this study on Atlantic salmon, supplemental cholesterol (1–1.5%) stimulated the production of bile acids, balancing cholesterol metabolism disrupted by the absence of fish meal. This implies that cholesterol from fish meal plays a vital role in modulating CYP7A1. External cholesterol sources can partially compensate for the effects of plant protein diets on cholesterol metabolism. Wei et al. (2020) studied Amur sturgeon and found that a complete replacement of fish meal with plant protein reduced CYP7A1 expression, which impaired bile acid homeostasis and intestinal health. The study highlighted those changes in bile acid metabolism also affected immune function, demonstrating a complex link between lipid metabolism and immunity. This suggests that bile acids, regulated by CYP7A1, may have secondary effects beyond digestion, particularly in immune modulation and liver function. The study on Nile tilapia by Li et al. (2023) emphasized that diets high in plant protein altered the fish’s cholesterol turnover. The suppression of CYP7A1 led to cholesterol accumulation, negatively affecting lipid metabolism and energy homeostasis. In some cases, the altered metabolism resulted in higher body adiposity and metabolic imbalances. These findings illustrate the importance of bile acid synthesis not only for excreting excess cholesterol but also for maintaining lipid equilibrium within the body. Chen et al. (2024) observed that largemouth bass fed mixed protein diets, including plant proteins, exhibited inhibited CYP7A1 expression. This suppression disrupted bile acid receptor signaling pathways (for example, FXR-RXR pathways), which play a role in cholesterol metabolism feedback mechanisms. The disruption reduced the fish’s ability to regulate cholesterol levels, illustrating the complexity of lipid regulation pathways. In some cases, partial replacement of fish meal with plant proteins was found to mitigate these negative effects. For example, Murashita et al. (2024) reported that mixed diets containing both animal and plant proteins maintained better bile acid metabolism, including CYP7A1 expression, compared to full plant-based diets. This suggests that gradual substitution or inclusion of specific supplements could help balance the metabolic effects. The studies collectively highlight the complex relationship between fish meal, plant proteins, and CYP7A1 expression. Plant protein diets reduce CYP7A1 expression, affecting the synthesis of bile acids and disrupting cholesterol metabolism. Cholesterol supplementation or mixed diets help restore bile acid metabolism, mitigating the negative impacts of fish meal replacement. Long-term disruption of CYP7A1 and bile acid metabolism can lead to fatty liver, metabolic disorders, and compromised immune function in fish. Gradual substitution strategies or additional lipid supplements may offer solutions to balance lipid metabolism in plant-based aquafeeds. CYP7A1 plays a pivotal role in cholesterol metabolism, and its suppression from plant protein diets can lead to various metabolic disturbances. While dietary cholesterol and partial substitution strategies show promise in mitigating these effects, further research is required to optimize plant-based diets for aquaculture, ensuring both growth performance and metabolic health.
While the upregulation of these cholesterol metabolic genes and enzymes in response to plant-based diets helps fish maintain cholesterol and lipid homeostasis, the long-term effects of these diets on fish health must be carefully monitored. Studies have shown that prolonged reliance on plant protein diets may lead to hepatic lipid accumulation and fatty liver disease in some species, as a result of altered lipid metabolism (Bou et al., 2017; Caballero-Solares et al., 2018; Mellery et al., 2015). Careful balancing of lipid content, the inclusion of essential fatty acids, and minimizing the anti-nutritional effects of phytosterols in plant-based diets are important to ensure optimal health and growth in aquaculture species. Table 10 contains research on the effect of substituting fish meal with some plant proteins on CYP7A1 activity in some fish species.
Effect of replacing fish meal with plant protein on CYP7A1 activity
| Species studied | Plant ingredients used | Effects on CYP7A1 | References |
|---|---|---|---|
| Atlantic salmon (Salmo salar L.) | Soy protein concentrates with 1–1.5% cholesterol | Restored CYP7A1 activity with cholesterol supplementation | Kortner et al. (2014) |
| Atlantic salmon (Salmo salar L.) | Microalgae and organic minerals | Enhanced lipid metabolism and improved CYP7A1 activity | Kousoulaki et al. (2016) |
| Gilthead seabream (Sparus aurata L.) | DL-methionine supplemented soy protein concentrate | Maintained CYP7A1 activity at optimal levels | Kokou et al. (2016) |
| Turbot (Scophthalmus maximus) | High plant protein with taurocholate supplementation | Partial restoration of CYP7A1 expression | Gu et al. (2017) |
| Atlantic salmon (Salmo salar L.) | Varied lipid sources (plant vs fish oils) | Differential effects on CYP7A1 activity | Hixson et al. (2017) |
| Atlantic salmon (Salmo salar L.) | Phytosterol-enriched plant oils | Altered CYP7A1 expression but no fatty liver development | Sissener et al. (2018) |
| Rainbow trout (Oncorhynchus mykiss) | Rapeseed meal, cottonseed meal | Reduced CYP7A1 activity and disrupted cholesterol metabolism | Zhu et al. (2018) |
| Japanese seabass (Lateolabrax japonicas) | Soy protein isolate, wheat gluten, corn gluten meal | Reduced CYP7A1 expression and compromised bile acid metabolism | Zhang et al. (2019 b) |
| Amur sturgeon (Acipenser schrenckii) | Full plant protein diet | Decreased CYP7A1 expression and impaired bile acid homeostasis | Wei et al. (2020) |
| Tiger puffer (Takifugu rubripes) | Soy protein isolate with taurine supplementation | Increased CYP7A1 expression | Xu et al. (2020) |
| Common carp (Cyprinus carpio) | Lupin meal and pea protein | Reduced CYP7A1 activity | Zhao et al. (2020) |
| Red seabream (Pagrus major) | Soy protein concentrate | Reduced CYP7A1 expression but maintained lipid homeostasis with fish oil supplementation | Takakuwa et al. (2023) |
| Nile tilapia | Corn gluten and soy protein concentrate | Suppression of CYP7A1 expression and reduced bile acid synthesis | Li et al. (2023) |
| Gilthead seabream (Sparus aurata) | Bile salt-supplemented plant diet | Improved bile acid metabolism and enhanced CYP7A1 expression | Ruiz et al. (2023) |
| Largemouth bass (Micropterus salmoides) | Mixed plant proteins (soy and corn) | Suppressed CYP7A1 and bile acid receptor signaling | Chen et al. (2024) |
Replacing fish meal with plant protein in aquafeeds supports sustainability but poses metabolic challenges, particularly in cholesterol regulation. Fish fed plant-based proteins upregulate LDLR and HMGCR to compensate for reduced dietary cholesterol, but phytosterols in these feeds inhibit cholesterol absorption. While these mechanisms maintain short-term cholesterol balance, long-term reliance on plant diets can result in hepatic fat deposition, altered lipid profiles, and impaired growth.
To mitigate these effects, strategic supplementation with cholesterol, essential fatty acids, and balanced lipid sources is crucial. A blend of plant and animal proteins tailored to species-specific needs will ensure optimal fish health and performance. Future research should refine plant-based feeds to achieve sustainable aquaculture without compromising fish welfare.