
Cereals are various grasses from the Poaceae (Gramineae) family. They are the world’s most important crops, grown in large quantities and used as staple foods. Due to a program known as the Green Revolution, which focused on fighting famine and increasing yield per plant, there was a significant increase in the production of high-yielding cereal crops worldwide in the second half of the twentieth century (FAO, 2017). In 2019, global production of the most important cereals – namely maize, wheat, rice and barley – amounted to 1,148, 768, 755 and 159 million tons respectively (FAOSTAT, 2019).
Weeds interfere with human activities (Vencill, 2002) and are considered strong competitors for crops due to their rapid germination, early growth, high seed production, and long-time persistence, leading to high population numbers (Baker, 1974). For weed regulation, policy making and biodiversity conservation, surveys and assessments of noxious weed lists are important tools that have been developed for a range of areas and purposes (Skinner et al., 2000).
Egypt has an area of about one million km2, with only 4% of the land being cultivated and the rest of the country is a desert. Barley and wheat (winter crops) and maize and rice (summer crops) are the most important components of crop rotation in the agro-ecosystems. Since prehistoric times, wheat and barley have been the most important cereals cultivated (Fahmy, 1997). The increase of human population in Egypt in contrast to the decline of ancient cultivated areas due to disturbances necessitates an expansion of cultivated areas. Therefore, several desert areas are being reclaimed and the total cultivated area of the country is being increased (Abd El-Ghani et al., 2017).
Weed communities in old world agroecosystems have evolved over centuries, supporting unique vegetation (Holzner & Immonen, 1982). Cropping systems that use mainly annual or even perennial crops are often accompanied by weeds, which are unintentionally growing plants adapted to the conditions of arable farming. These weeds contribute to plant diversity in their sites as they also provide ecosystem services or are themselves targets of nature conservation (Médiène et al., 2011). In general, weeds are ubiquitous in arable soils, and their abundance and occurrence vary geographically and climatically. Based on their phenology, weeds in Egypt can be divided into 3 main groups: (1) winter weeds, which are restricted to the cooler months of the year, (2) summer weeds, which tend to be restricted to the warmer months of the year, and (3) all-year weeds, which are present and biologically active throughout the year (Abd El-Ghani et al., 2015).
Several studies have focused on identifying the most important factors influencing the diversity and distribution of weed composition (Lososová et al., 2004; Pinke et al., 2009; Šilc et al., 2009). Crop type (Fried et al., 2008), crop cycle (Barralis, 1976; Cardina et al., 2017), crop seasonality (Mahgoub, 2021), soil type (Dale et al., 1992; Mahgoub, 2019a), geographical gradients and climatic characteristics (Fried et al., 2008), surrounding landscape (Fanfarillo et al., 2023) and management practices and ecological conditions (Marshall et al., 2003; Sher & Al-Yemeny, 2011; Stevenson et al., 2017) were included among the factors controlling the species richness and composition of weed communities in arable fields.
A wealth of studies on the composition and diversity of weed flora in cereals have been conducted worldwide, for example in Nepal (Nowak et al., 2016), Germany (De Mol et al., 2015), western and central Italy (Abbate et al., 2013), Iran (Hassannejad & Ghafarbi, 2013), Finland (Salonen et al., 2011), central and western France (Gaba et al., 2010), New Zealand (Rahman et al., 2006) and Argentina (Poggio et al., 2004). In Egypt, several quantitative studies have been conducted on weed composition, diversity and soil properties of croplands, including cereals, in different bioclimatic regions (Al-Sherif et al., 2018; El-Ghani et al., 2013; Salama et al., 2016; Salama et al., 2017). In most of these studies, multivariate analysis techniques were applied to assess the role of soil properties in the relationship between crops and weeds. However, few quantitative studies have been conducted on the role of other environmental factors controlling weed composition in association with crops (Alsherif, 2020; Mahgoub, 2021).
Therefore, the present study focuses on the diversity and distribution of weed species associated with common cereals in arable lands of different bioclimatic regions in Egypt and to assess the effects of crop type, crop seasonality, region and physicochemical soil properties on weed species composition.
Egypt consists of 29 administrative governorates and can be divided into climatically and geographically different regions: Upper and Lower Egypt, the western and eastern deserts and the Sinai Peninsula. The southern highlands of Upper Egypt comprise the area of the Nile Valley from the southern border with Sudan to the north of the Beni Suef governorate, while Lower Egypt, or the Nile Delta, stretches from the Giza governorate to the Mediterranean Sea.
The cultivated land in the study area can be divided into old lands (Nile Delta and Valley), that have been cultivated since ancient Egypt, and new reclaimed desert land (Wadi El-Natrun), which has been cultivated in the last six decades. The old land is very fertile, while the new reclaimed land is less fertile, more fragile and has limited access to high quality water for irrigation (Bakr & Bahnassy, 2019). The studied area included five governorates (localities) in three major bioclimatic regions (Figure 1): 1 – The Nile Delta region: the governorates of El-Sharkiya and El-Menoufiya, 2 – The Nile Valley: the governorates of Beni Suef and Qena, and 3 – The Western Desert: the Wadi El-Natrun depression (referred to as Natrun in the text; the governorate of El-Buhaira).
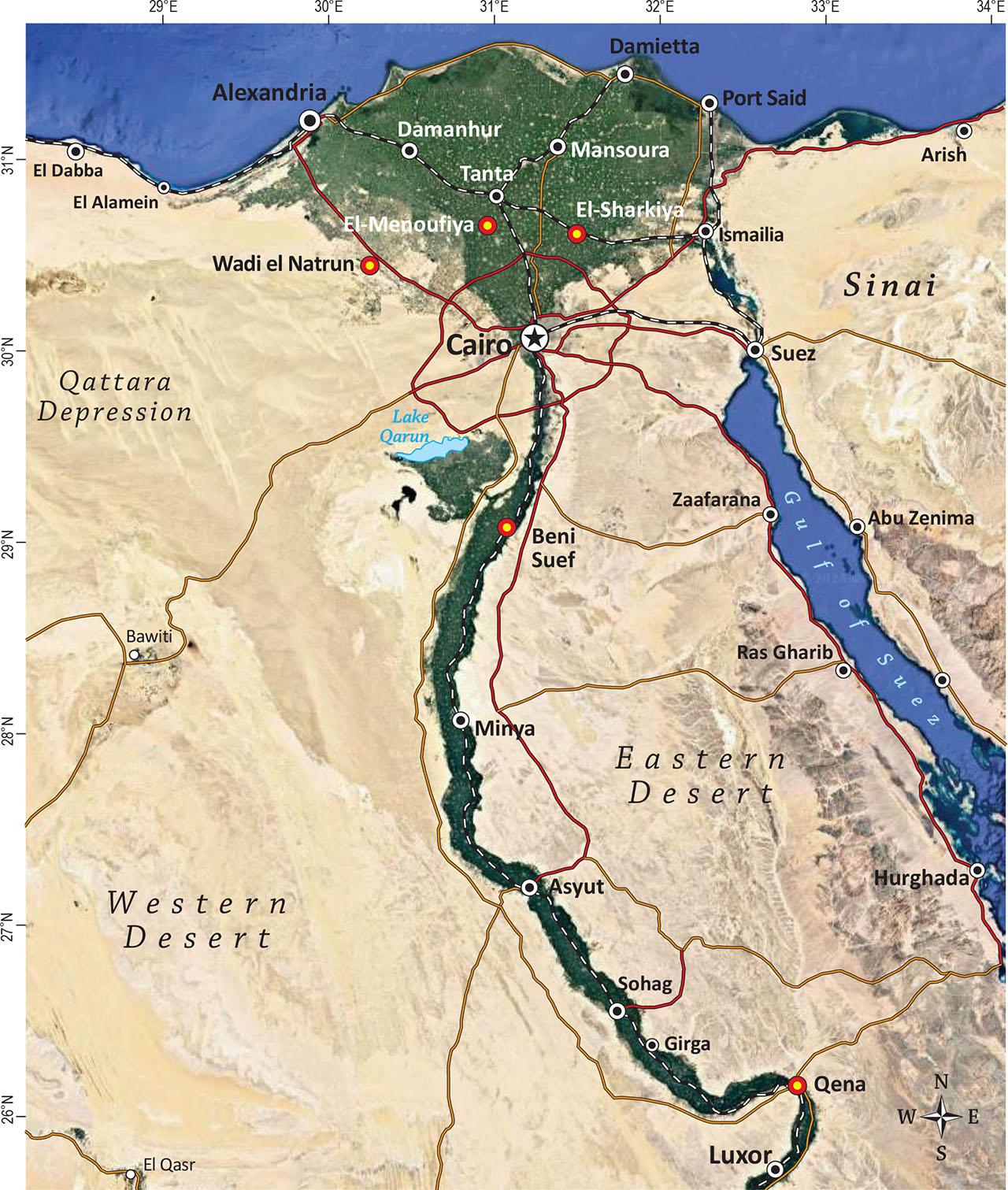
Map with the five selected regions (governorates), represented by red dots.
Slika 1: Zemljevid s petimi izbranimi območji (upravnimi enotami), prikazanimi z rdečimi točkami.
The Nile Valley has an extremely dry climate: high evaporation, high temperature and negligible rainfall (1.3 to 5.4 mm per year), low relative humidity (Table 1). The Qena governorate in the southern Egypt has a subtropical, rainless climate, but in winter there can be torrential rainfall coming from the Red Sea Mountains. The climatic aridity gradually decreases northward towards the delta (annual rainfall ranges between 20.8 and 45.5 mm per year), as does the rest of Egypt’s northern coast, which is the wettest region in the country and has relatively moderate temperatures. Natrun is a hyperarid bioclimatic area with hot summers and mild winters characteristic of the Egyptian desert (Ayyad & Ghabbour, 1986). In the driest areas (Natrun), the highest precipitation was measured in January and the lowest in April (4.9 and 0.8 mm respectively).
Selected geographical, agricultural and climatic information (mean values of the 10 years from 2011 to 2021) for the nearest meteorological station in the studied regions.
Tabela 1: Izbrani geografski, kmetijski in klimatski podatki (povprečne vrednosti za deset let med 2011 in 2021) za najbližje meteorološke postaje v raziskovanih območjih.
| Region (Governorates) | Latitude | Longitude | Total Area (km2) | Cultivated Area (%) | Elevation (m asl) | Tmax (°C) | Tmin (°C) | Tmean (°C) | Rainfall (mm) |
|---|---|---|---|---|---|---|---|---|---|
| El-Sharkiya | 30°35′ | 31° 30′ | 4180 | 81 | 13.00 | 29.6 | 15.5 | 22.5 | 33.5 |
| El-Menoufiya | 30°36′ | 30°1′ | 2543 | 57 | 17.90 | 29.4 | 15.3 | 22.4 | 34.6 |
| Beni Suef | 29°4′ | 31°6′ | 1322 | 91 | 30.40 | 29.9 | 15.4 | 22.6 | 7.80 |
| Qena | 26°10′ | 32°43′ | 1796 | 75 | 72.60 | 31.2 | 17.1 | 24.1 | 1.00 |
| El-Buhaira | 30°58′ | 30°33′ | 281.7 | 13.4 | −23.00 | 29.2 | 15.4 | 22.3 | 41.4 |
Field data on floristic composition were collected during two seasonal visits (summer and winter) between March 2019 and May 2021 at the study area sites to survey the weed species associated with the main cereal crops and record their distribution patterns. Two hundred and sixty-one relevés (fields) in five visited governorates were selected to represent the agroecosystem of different bioclimatic regions in Egypt. The selection of the studied fields was based on: (i) non-application of herbicides and (ii) conventional irrigation through a network of irrigation canals. A three-year crop rotation is typically implemented in Egypt's agricultural areas as follows: (1) temporary Egyptian clover (or fallow fields) – cotton; (2) wheat – maize (or rice in the northern Delta) and (3) permanent Egyptian clover (or broad beans) – maize. in order to grow all crops in a single year, an area is typically divided into three portions (El-Khshin et al., 1980). winter crops are sown in September through November; cotton is planted in February through March; maize and rice are planted in April through May. For all crops except cotton (which lasts7–8 months), crop longevity is 5–6 months.
Within each governorate, the number of relevés varied depending on the area under cultivation (Beni Suef – 26 relevés, El-Menoufiya – 90 relevés, Natrun – 28 relevés, Qena – 84 relevés, El-Sharkiya – 33 relevés). Four main cereals were analysed: barley (Hordeum vulgare L.) and wheat (Triticum aestivum L.) as winter crops, rice (Oryza sativa L.) and maize (Zea mays L.) as summer crops. We sampled 107 relevés for wheat, 100 for maize, 38 for rice and 16 for barley (Table S1). The area of each relevé was 20m × 100m (Salama et al., 2017). To avoid the effects of field edges (Marshall & Arnold, 1995), relevés were made at least 10 m from the field edge or road verges in a homogeneous weed vegetation within the field.
Within each relevé presence/absence of each species was recorded (Table S2). The specimens were identified in the herbarium of Cairo University (CAI). The nomenclature followed Boulos (1995–2009). Life form and chorotype categories were determined according to Raunkiær (1934) and Salama et al. (2016, 2017), respectively (Table S3).
The soil samples were collected from all 261 sampling plots. Three soil samples were taken from a depth of 0–25 cm, mixed to a composite sample, air-dried and then sieved through a sieve (2 mm) to remove debris. Soil water extracts (100 g soil in 500 ml distilled water) were prepared to measure pH (pH meter; Model Lutron pH 206), electrical conductivity (conductivity meter; Model Corning, NY, 16542, USA), bicarbonates (Allen et al. 1974), chloride (Davey & Bembrick, 1969), calcium and magnesium (Jackson, 1967) using titration methods. Sulfates were determined using a turbidimetric technique (spectrophotometer; Model 1200; Bardsley & Lancaster (1965)), while potassium and sodium were determined using the flame photometer technique. Organic matter was determined using the Walkley and Black (1934) method. Soil particles of different sizes (sand, silt and clay) were quantified using the international pipette method (Pansu & Gautheyrou, 2007).
Species richness was enumerated as number of species per relevé (α-diversity). We defined the species pools of the four different bioclimatic regions and crop types for comparative analyses as number of all species sampled that could be easily compared. Beta (β) diversity was measured to estimate the variation among local assemblages due to the species turnover between sample plots or habitats (Koleff et al., 2003). The data set was partitioned according to the groups (bioclimatic region, crop type, crop seasonality). β-diversity was calculated as the mean Whittaker dissimilarity for all pairs of relevés using the JUICE program (Tichý, 2002). The confidence intervals of Sørensen similarity index were measured using bootstrap technique sampling (500 samples) from that partition. The phi-coefficient (φ) index (Chytrý et al., 2002) as a fidelity measure was calculated to detect diagnostic species for particular region or crop. Fisher’s exact test was applied to determine the significance of fidelity, after standardising the size of all groups to equal size (p<0.05; Tichý & Chytrý, 2006). We used the phi threshold value of 0.30 for diagnostic species.
The rule introduced by Lepš & Šmilauer (2003) was used to select an appropriate ordination method for direct gradient analysis. Detrended correspondence analysis (DCA) showed a short gradient in species composition (axis length 3.31 SD), so we used linear methods for further analysis. Redundancy analysis (RDA) was performed to separate the effects of crop type, crop seasonality, bioclimatic regions and soil variables to evaluate the response of weed species to these ecological factors. To exclude multicollinearity between the explanatory variables, the variance inflation factor (VIF) was calculated for all variables; therefore, Cl and Na were removed from further analysis. Monte Carlo permutation (999 permutations) was used to determine the statistical significance of a certain group of variables. Ordination analyses were performed in R (R Development Core Team, 2012) with the package vegan (Oksanen et al., 2019). In combination with RDA, variation partitioning was employed to estimate the linear partial effect of the four factors under investigation. The gross effect represented the variation explained by the predictor as the only explanatory variable, while the net effect was calculated using one specific explanatory variable and all others as covariates (Lososová et al., 2004). The explained variation was compared by adjusted R2 values. ANOVA (999 permutations, permutation test for RDA under reduced model) was applied to test the significance of the variables.
The species richness and beta diversity were analysed using non-parametric Kruskal–Wallis tests and Games-Howell post-hoc tests in program SPSS.
A total of 250 plant species were recorded in the 261 relevés of the surveyed arable fields in the five bioclimatic regions of Egypt. The plant species were represented by 47 families; Poaceae (20.8%), Asteraceae (13.25), Fabaceae (9.2%), Brassicaceae (7.6%), Chenopodiaceae (5.2%) and Euphorbiaceae (4.4%) were the most common, accounting for about 60.3% of the total flora. Among the growth forms, annual herbs dominated, accounting for 66.6%. The percentage distribution of life forms in the current study revealed that therophytes ranked first (163 species, 65%), followed by cryptophytes, hemicryptophytes, phanerophytes, and chamaephytes (Figure 2). Chorologically, more than half (55%) of the recorded species belong to Mediterranean (15.6%), Saharo-Arabian (15.5%), Cosmopolitan (13.2%) and Irano-Turanian (11.1%) chorotypes (Figure 3).
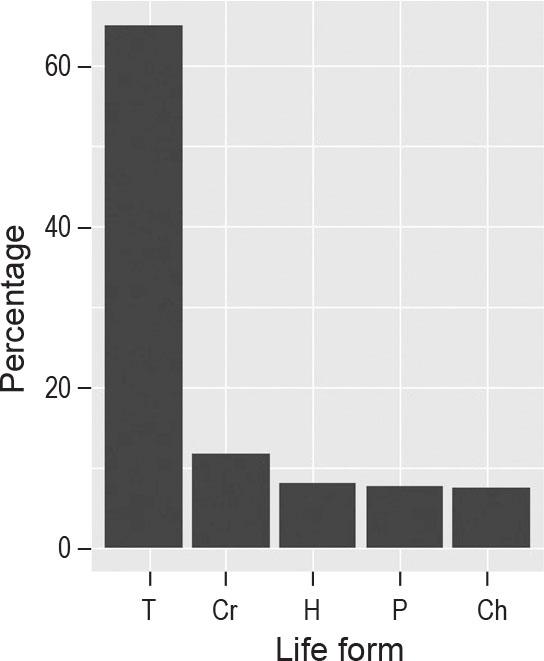
Life form spectrum of weed vegetation associated with cereals in study areas. T = Therophytes, Cr = Cryptophytes, H=Hemicryptophytes, P = Phanerophytes, CH = Chamaephytes.
Slika 2: Spekter življenskih oblik plevelne vegetacije v žitih na obravnavanih območjih. T = terofiti, Cr = kriptofiti, H = hemikriptofiti, P = fanerofiti, CH = hamefiti.
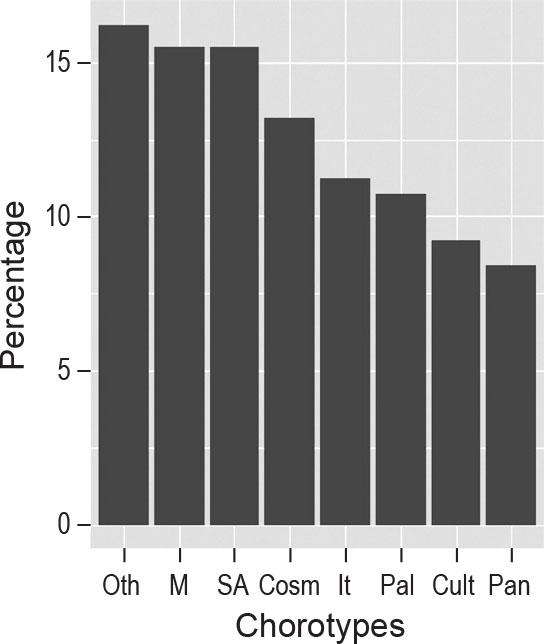
Chorotypes of weed vegetation associated with cereals in study areas. M = Mediterranean, SA = Saharo-Arabian, Cosm = Cosmopolitan, IT = Irano-Turanian, Pal = Paleotropical, Cult = Cultivated, Pan = Pantropical, Oth = Other.
Slika 3: Horotipi plevelne vegetacije v žitih na obravnavanih območjih. M = Sredozemski, SA = Saharo-Arabski, Cosm = kozmopolitski, IT = Irano-Turanski, Pal = Paleotropski, Cult = Gojeni, Pan = Pantropski, Oth = Ostali.
The most frequent species were Cynodon dactylon, Sonchus oleraceus, Convolvulus arvensis and Chenopodium murale. The total number of species differed from one crop to another. The highest number (species pool) was 184 species in wheat, followed by 155 species in maize, 114 species in rice, and 112 species in barley. The total number of species in winter crops was 197 and in summer crops 188. Fifty-one species were found in all four cereals, of which some species were more frequent in one (or more) crops than in the others. Cynodon dactylon, Sonchus oleraceus, Chenopodium murale, Malva parviflora and Melilotus indicus were most frequent in the winter crops (wheat and barley, frequency=76–53%), and Cynodon dactylon, Echinochloa colona and Portulaca oleracea were common in the summer crops (rice and maize, frequency=73–58%). Species characteristic of wet and moist soils, e.g. Echinochloa crus-galli and Cyperus difformis, were the most frequent weeds in rice fields.
Species fidelity using the phi (φ) coefficient index showed the diagnostic species for each region, 83 weed species were diagnostic for El-Sharkiya, 9 for Beni Suef, 3 for Qena, 4 for El-Menoufiya and 2 for Wadi Natrun (Table S4). While 60 weeds were reported as diagnostic species for different crops, 49 species were diagnostic for barley, 6 for rice, 3 for wheat and 2 for maize. Maize fields were characterized by Corchorus olitorius and Dine-bra retroflexa (Table S5). On the other hand, 32 species were diagnostic of barley, among them Lepidium sativum, Medicago orbicularis, Medicago laciniata and Anagallis arvensis var. arvensis, and 6 species of rice fields (Table S5).
Species richness showed considerable differences between regions and crops (Figure 4). The regions studied showed similar average species richness (10.0–14.2 species per plot), except for El-Sharkiya governorate (located in the Nile Delta), that deviated (47.6 species per plot). Among the crops studied, barley had the highest species richness (average 33.0 species per plot) with the largest range (18–72 species), while species richness was relatively low in the other cereals (average 12.1–16.6 species per plot), and the differences were not statistically significant. In terms of crop seasonality, the differences in species richness between winter and summer crops were not statistically significant (average 13.1–18.7 species per plot).
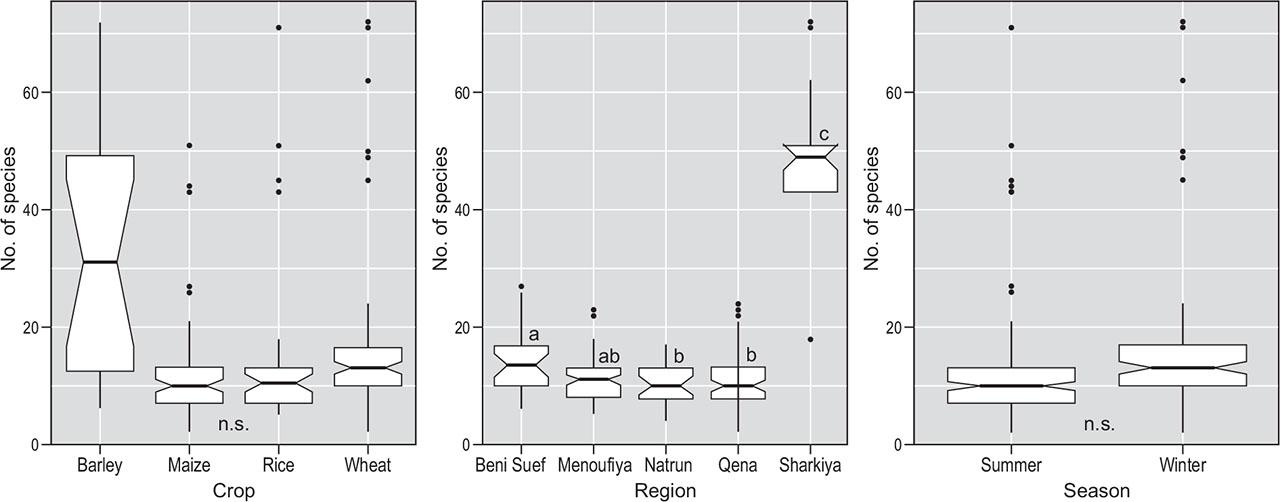
Species richness (species number per relevé) in different crops, bioclimatic regions and seasons. Box-plots: horizontal line represents median, box’s upper and lower borders 1st and 3rd quartiles, vertical line represent the highest and lowest value excluding outliers and dots the outliers. Letters display pairwise treatment comparisons with Kruskal-Wallis test and Games-Howell posthoc test.
Slika 4: Vrstna pestrost (število vrst na vegetacijski popis) v različnih poljščinah, bioklimatskih regijah in letnih časih. Škatle z brki: vodoravne črte predstavljajo mediano, zgornji in spodnji rob škatel predstavljata prvi in tretji kvartil, navpična črta največjo in najmanjšo vrednost brez osamelcev, točke predstavljajo osamelce. Črke predstavljajo parne primerjave s Kruskal-Wallisovim testom in Games-Howell posthoc testom.
The beta diversity index (Figure 5) showed statistically significant differences between all bioclimatic regions with the lowest beta diversity by far in El-Sharkiya governorate. Among crop types, the lowest diversity was found in rice and the highest in maize. All were statistically significantly different. The difference in Beta-diversity between the two crop seasonalities (summer/winter) was not statistically significant.
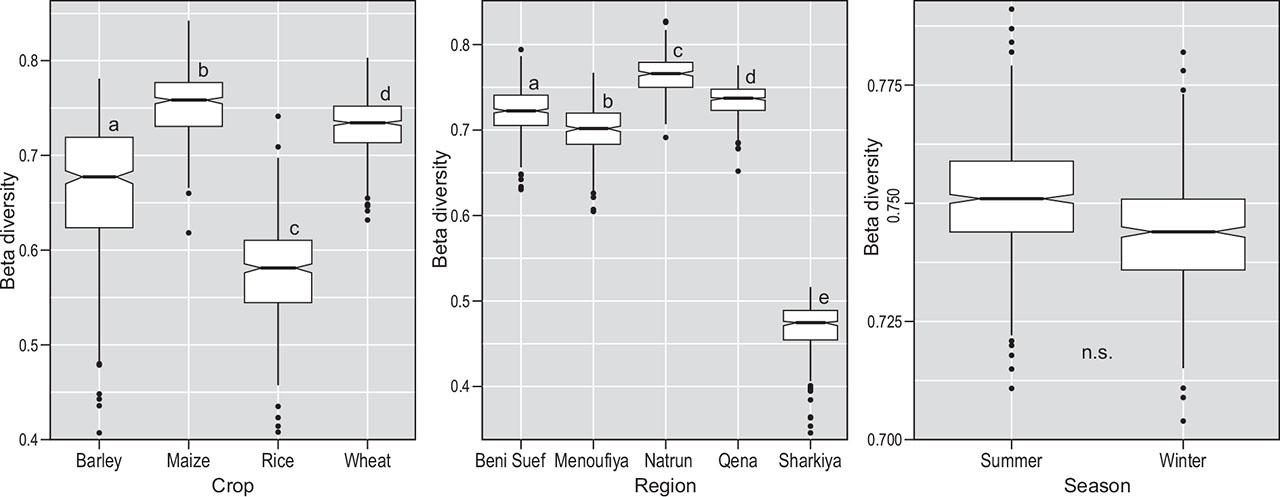
Beta diversity along three gradients (bioclimatic regions, crops and seasons). Sørensen index was calculated by bootstrap method. Letters display pairwise treatment comparisons with Kruskal-Wallis test and Games-Howell posthoc test.
Slika 5: Beta diverziteta vzdolž treh gradientov (bioklimatsko območje, kulturna rastlina, letni čas). Sørensenov indeks smo izračunali z metodo bootstrap. Črke predstavljajo parne primerjave s Kruskal-Wallisovim testom in Games-Howell posthoc testom.
Soils in rice fields differed the most, being statistically different from the other crops in almost all the characteristics studied (Table S6). The differences in soil properties are more pronounced between the different regions. Higher organic matter was found in El-Sharkiya, Qena and especially Natrun and differed significantly from El-Menoufiya and Beni Suef. The regions also differed significantly in the proportion of fine sand (FS) (Table S6).
Detrended Correspondence Analysis (DCA) of species composition was employed as a preliminary analysis and showed long gradients for the first two axes (Figure 6). The first axis was related to crop type and seasonality, while the second axis was related to other explanatory variables (particularly pH and Mg).
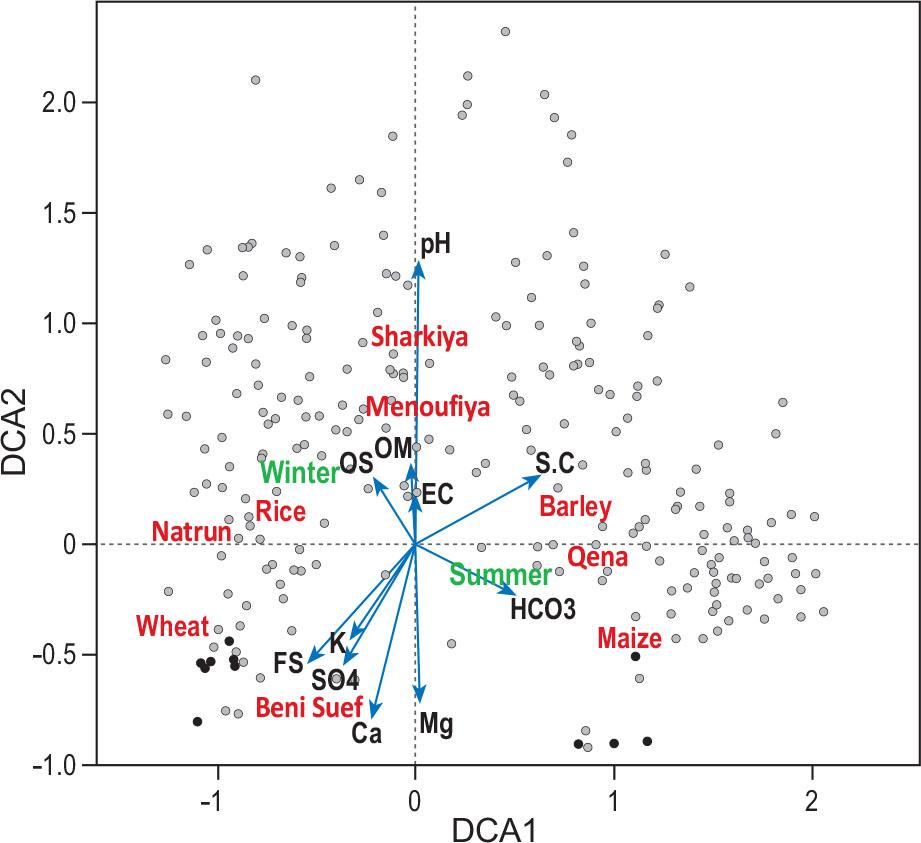
DCA analysis of plots and explanatory variables passively projected onto the diagram of weed community in Egypt. Numerical variables are presented as blue arrows, categorical variables as centroids in red text and sampling plots as grey points. Eigenvalues of first two axes: 0.5077 and 0.3183. Abbreviations: CS – coarse sand, FS – fine sand, S.C – silt and clay, OM – organic matter, pH – soil reaction, EC – electrical conductivity.
Slika 6: Analiza DCA popisnih ploskev in razlagalnih spremenljivk pasivno projiciranih na diagram plevelnih združb v Egiptu. Numerične spremenljivke so prikazane kot modre puščice, kategorične spremenljivke kot centroidi z rdečim tekstom in popisne ploskve kot sive točke. Lastne vrednosti za prvi dve osi: 0.5077 in 0.3183. Okrajšave: CS – debelozrnat pesek, FS – drobnozrnat pesek, S.C – melj in glina, OM – organska snov, pH – reakcija tal, EC – električna prevodnost.
Redundancy analysis (RDA) summarized the variation in species composition of weed communities in relation to the following explanatory variables: crop type, crop seasonality, bioclimatic regions and soil properties (Figure 7). In the RDA ordination, the first two axes explained 22.4% and 12.1% of the total variation, respectively. The vectors length and position of centroids showed that weed composition was mostly affected by bioclimatic region (highest explained variance of weed composition; 23.1%), followed by crop type (12.4%) and crop seasonality (7.6%). Among soil variables fine sand, silt and clay were the most important. The RDA results which were displayed in ordination diagrams (Figure 7) indicated that most positively associated species with the first axis were Beta vulgaris ssp. maritima, Chenopodium album and most negatively associated were Trianthema portulacastrum and Dactyloctenium aegyptium.
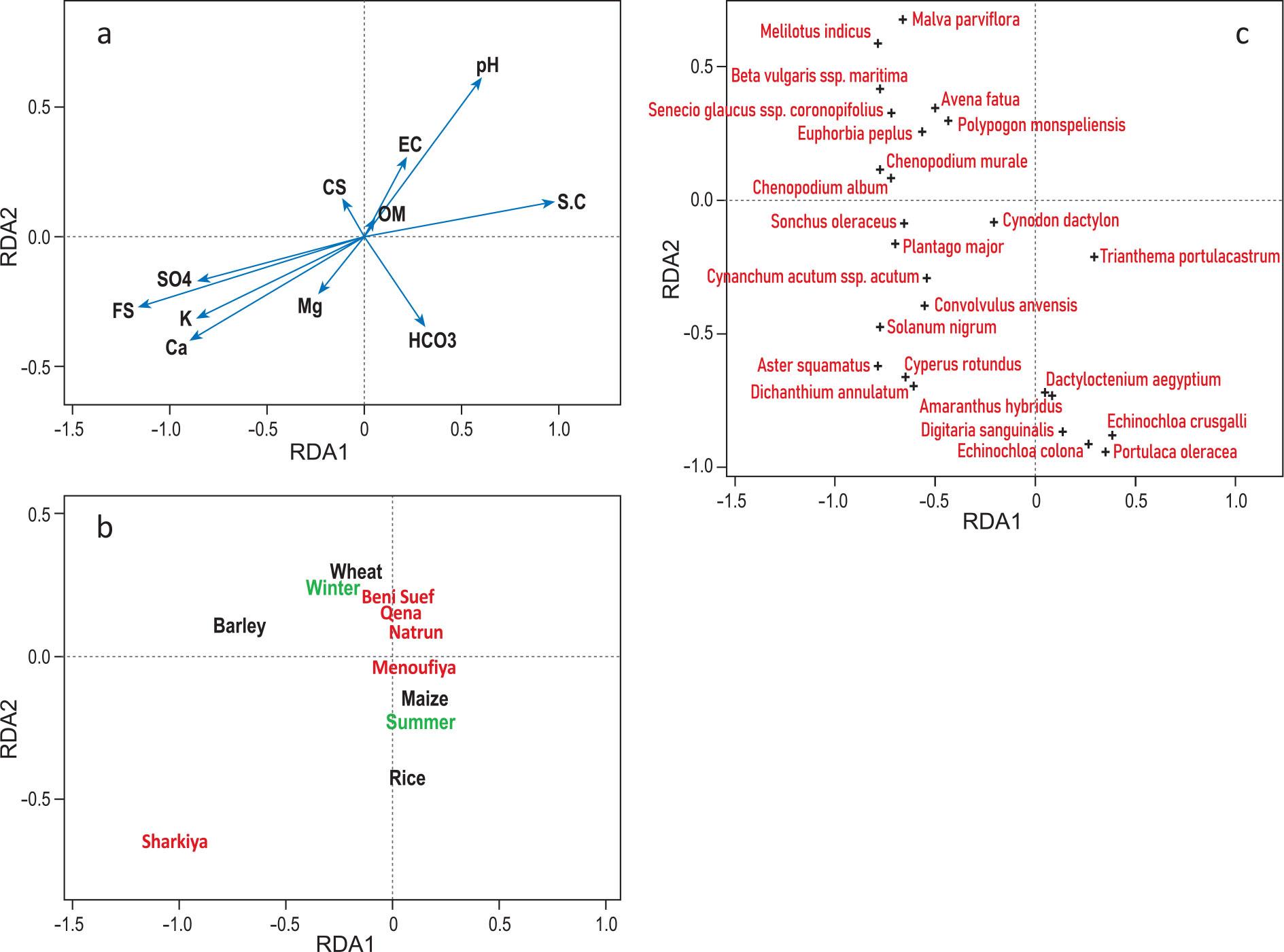
The ordination diagrams of the reduced RDA containing (a) soil variables as vectors, (b) crop, region and season as factors and (c) species. Only the species with the highest fit were presented (25 species with highest frequencies and highest fit along the first axis). For the soil variables abbreviations see Figure 6.
Slika 7: Ordinacijski diagrami reducirane RDA, ki vsebujejo: (a) talne spremenljivke kot vektorji, (b) kulturna rastlina, območje in letni čas kot faktorji in (c) vrste. Prikazane so samo vrste, ki imajo največji fit (25 vrst z največjo frekvenco in fitom vzdolž prve osi). Za talne spremenljivke glej Slika 6.
In the redundancy analysis (RDA) model, the variation partitioning between explanatory variables (net effects) clarified that bioclimatic region (9.4%) and crop type (2.4%) were again the most important variables, while crop seasonality was not significant (Table 2). The partitioning results (not shown) indicated percentage contributions of groups of explanatory variables (region, crop and soil) to the variation in weed species composition showing that region and crop type have higher values (9.4 and 2.4 respectively), while the soil variables have lower net effect.
Gross and net effects of the explanatory variables on the weed vegetation resulting from partial RDA analysis with single variable. RDA explained variation 0.366. For the soil variables abbreviations see Figure 6.
Tabela 2: Bruto in neto učinek razlagalnih spremenljivk na plevelno vegetacijo za posamezno spremenljivko v parcialni RDA. Pojasnjena varianca v RDA je 0.366. Za talne spremenljivke glej Slika 6.
| Gross effect | Net effect | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | df | Explained variance % | R2adj. | F | p-value | Explained variance % | R2adj. | F | p-value |
| CS | 1 | 0.881 | 0.0050 | 2.3023 | 0.0220 * | 0.245 | −0.0002 | 0.9289 | 0.4460 |
| FS | 1 | 3.782 | 0.0341 | 10.1800 | 0.0010 *** | 0.245 | −0.0002 | 0.9291 | 0.4790 |
| S.C | 1 | 3.527 | 0.0315 | 9.4679 | 0.0010 *** | 0.245 | −0.0002 | 0.9287 | 0.4880 |
| OM | 1 | 0.730 | 0.0035 | 1.9032 | 0.0600 . | 0.491 | 0.0024 | 1.8626 | 0.0330 * |
| pH | 1 | 2.495 | 0.0212 | 6.6278 | 0.0010 *** | 0.334 | 0.0008 | 1.2663 | 0.1200 |
| EC | 1 | 0.696 | 0.0033 | 1.8162 | 0.0580 . | 0.320 | 0.0006 | 1.2138 | 0.1820 |
| HCO3 | 1 | 1.100 | 0.0072 | 2.8799 | 0.0050 ** | 0.302 | 0.0004 | 1.1453 | 0.2330 |
| SO4 | 1 | 2.044 | 0.0167 | 5.4057 | 0.0020 ** | 0.277 | 0.0001 | 1.0523 | 0.3470 |
| Ca | 1 | 2.153 | 0.0177 | 5.6975 | 0.0010 *** | 0.233 | −0.0003 | 0.8837 | 0.6160 |
| Mg | 1 | 1.102 | 0.0072 | 2.8864 | 0.0110 * | 0.421 | 0.0017 | 1.5959 | 0.0730 . |
| K | 1 | 2.187 | 0.0181 | 5.7894 | 0.0010 *** | 0.383 | 0.0013 | 1.4516 | 0.0640 . |
| Crop | 3 | 12.103 | 0.1108 | 11.7950 | 0.0010 *** | 2.410 | 0.0201 | 4.5740 | 0.0010 *** |
| Season | 1 | 7.629 | 0.0727 | 21.3900 | 0.0010 *** | 0.000 | 0.0000 | 0.0000 | |
| Region | 4 | 22.430 | 0.2122 | 18.5060 | 0.0010 *** | 9.359 | 0.0888 | 8.9702 | 0.0010 *** |
In our study, we found 250 species in 261 sampled fields (197 in winter and 188 in summer crops). Weed species number in winter crops is comparable to findings from Mediterranean region (Arslan, 2018; Chakkour et al., 2023; Cirujeda et al., 2011), although also higher values were reported in Italy (Fanfarillo et al., 2023; Fanfarillo et al., 2020), and in traditional cereal fields in Algeria and NW Morocco (Deil, 1997; Kazi Tani et al., 2010). Compared to other studies of weeds in summer crops (maize) (Fanfarillo et al., 2019), in Egypt species richness is higher. Slightly higher species richness could be explained by inclusion of rice fields with specific flora into the analysis, although rice fields are usually species poor (Pinke et al., 2014).
Beta diversity is very different between regions, with the most uniform weed vegetation in El-Sharkiya. Although climate and latitude are the main drivers of species composition (Fanfarillo et al., 2023), in case of El-Sharkiya, we could hypothesise that low beta diversity is due to agricultural measures. Among crop types, rice had lower beta diversity although species richness was comparable to other crops. Special environmental conditions (stagnant water) are the reason for less diverse and uniform vegetation.
The 250 recorded species in the investigated cereals belong to 47 families. Poaceae, Asteraceae, Fabaceae, Brassicaceae, Chenopodiaceae, and Euphorbiaceae were the largest six species-rich families and also represent the most common families in the Mediterranean and North African flora (Quézel, 1978) and, specifically, in the Mediterranean arable flora (Fanfarillo et al., 2020). This is partially in agreement with the findings in the agroecosystems of different bioclimatic regions of Egypt: in Nile Delta (Abd Al-Azeem, 2003; Abd Alla, 2007; Mashaly, 2003; Mashaly et al., 2013), in Nile Valley (Alsherif, 2020; Mustafa, 2002; Salama et al., 2017), in Oases of the Western Desert (Abd El-Ghani et al., 2015), and in reclaimed lands (Shehata & El-Fahar, 2000; Soliman, 1996). These plant families were frequently recorded during many surveys of weed flora in other parts of the Africa: Algeria (Eddoud et al., 2018), Morocco (Chakkour et al., 2023), Ethiopia (Firehun & Tamado, 2006) and Arabic peninsula: Saudi Arabia (Gomaa, 2012), Oman (El-Sheikh, 2013).
Poaceae had the highest number of species (52 species) because of their ability to flourish in numerous ecosystems and their tolerance towards salinity, freeze, drought and various abiotic stresses (Landi et al., 2017). They occur in different habitats in Egypt, including salt marshes and arid areas and they are particularly common in old lands in the Nile Valley (Boulos & Fahmy, 2007).
In terms of plant longevity, annuals comprised the majority (168 species, 66.6%) of the total flora. Their dominance can be explained by their enormous reproductive capacity, morphological, ecological and genetic adaptability under high levels of disturbance, such as agricultural activities (Grime, 2002). The high number of annual species throughout the favourable seasons can be explained by the short life cycles of farm crops, and also prevailing dry climate conditions, low water availability, substrate instability, and agrotechnical measures (Shaltout & El Fahar, 1991; Storkey, 2010). Life-form spectra recorded in current study are similar to other studies on Egyptian agro-ecosystem (Al-Sherif et al., 2018; Salama et al., 2016). Similar result was reached by Da Costa et al. (2007) who indicated that annual plants were the predominant growth form in semi-arid and arid regions. On the contrary, the relatively high number of trees and shrubs (13.5%) found in our study reflects seedlings originating from the neighbouring land.
Phytogeographically, Egypt lies at a junction of floristic elements from four separate regions (El Hadidi, 1993), namely the Euro-Afro Asiatic Mediterranean, the Afro-Asiatic Saharo-Arabian, the African Sudano-Zambezian, and the Asiatic Irano-Turanian. The chorological analysis showed that Mediterranean and Saharo-Arabian elements represent one third of the recorded weed species. El-Demerdash et al. (1997) suggest that Egypt has a transitional Mediterranean character between the pure Mediterranean and the Saharo-Arabian territory. The Mediterranean species dominated the weed flora and showed the highest number in the Nile Delta regions, followed by the Nile Valley, and Natrun indicating more mesic environmental conditions. Similar results were reported in the cultivated areas in Nile Delta (El Saied & Bedair, 2018) and Nile Valley (El-Ghani et al., 2013; Salama et al., 2016). The Saharo-Arabian species were highly represented in Nile Valley and Natrun (22.1% and 16.8%, respectively) compared to Nile Delta (5%). The Saharo-Arabian elements are indicators of arid hot climate, as they have a high tolerance to the extreme harsh environmental conditions. This is in agreement with results of Abd El-Ghani et al. (2015) in Natrun, Salama et al. (2017) in Qena governorate, Al-Sherif et al. (2018) in Fayoum depression of the Western Desert, and Gomaa (2012) in Saudi Arabia. The other floristic elements comprised 34.1% in the whole dataset, reflecting their capability to penetrate the study area. Willcox (2012) indicated the increase of weed migration among Mediterranean region, Middle East and Europe in recent decades due to increased migration of people and transcontinental commerce.
Egypt is ecologically a very diverse country, although majority of land is desert. Nevertheless, agriculture is the most important sector of the Egyptian economy (El-Ramady et al., 2018). But most of the agriculture is concentrated in the Nile Delta, Valley and a number of oases where soil and site conditions are very similar. Therefore, important impact of crop type on weed diversity was expected.
Crop type had second most profound effect on floristic composition of arable fields, which supports the evidence that crop is the primary factor influencing the weed community (Cimalová & Lososová, 2009; Fried et al., 2008; Mahgoub, 2019b; Nagy et al., 2018; Nowak et al., 2015; Šilc et al., 2009). Most previous studies included a wide variety of crops (cereals and also root crops), with very diverse management practices, particularly in terms of time and frequency of disturbance. In contrast, the current study concentrated only on cereal crops, making it even more unexpected that the crop type is the second most significant factor in the weed floristic composition. Importance of the crop type, at least in Europe, also increases along the gradient from north towards the south (Šilc et al., 2009).
The species pool varied among crop types, although we must consider uneven number of relevés in various crops (Table S1); wheat had the highest number of weeds, followed by maize and rice. On the contrary, barley had the lowest number of species and the largest range of species richness, which could be explained by different land intensity in crops.
The most frequent species Cynodon dactylon, Sonchus oleraceus, Convolvulus arvensis and Chenopodium murale indicated their wide ecological amplitudes, while others were limited to a specific crop with very narrow amplitude. This finding is in line with recent researches in Egypt (Mahgoub, 2019a; Shaheen, 2002), which found that these species were widely distributed (ubiquitous) in significant quantities, owing to heterogeneity and phenotypic plasticity. Cynodon dactylon, according to Håkansson (1982), is one of the world’s worst weeds, conquering most temperate and subtropical agro-ecosystems, and growing practically on all soil types, rapidly invading cultivated area. Sonchus oleraceus is a weed of a wide range of environments including cultivated areas as it grows and flowers very quickly and produces numerous light seeds with no or low innate dormancy, that easily disperse by wind or water (Peerzada et al., 2019). Chenopodium murale, one of the dominant recorded species, is a well-known weed with its high growth rate, high reproductive capacity, and environmental flexibility. In agreement with our results, Chenopodium murale is a strong competitor to other weeds in different conditions (regions and crops). Additionally, it has a negative effect on various crops by accumulating potassium, magnesium, nitrogen, and phosphorus more efficiently than other weed species (Qasem, 1997). Some weeds showed higher frequency occurrences in certain crops. The habitat preference phenomena may be the reason of restricted distribution of some weeds. A weed with a life cycle similar to that of a crop will typically be a more successful competitor (Zimdahl, 2007). In line with El-Hadidi & Kosinová (1971) and Abd El-Ghani & Amer (1990), Anagallis arvensis var. caerulea, Malva parviflora, and Melilotus indicus were diagnostic species of wheat crop.
Results also indicated the occurrence of certain species of wet and moist soils (e.g., Cyperus difformis, Echinochloa colona, Echinochloa crus-galli) associated with rice fields with its high water needs. Turki and Sheded (2002) and El-Kassaby et al. (2011) enumerated the most common weeds associated with rice fields in the Nile Delta, and suggested that rice fields represent a transitional ecosystem between the agro-ecosystem and the natural water ecosystem. The first comprehensive contribution to the weed flora of the rice fields in Egypt was that of Imam & Kosinová (1972) who reported commonly occurring 14 weed species, 11 of them being specific for rice crops only, the remaining three being present in rice as well as in various other crops. The rice weeds included both aquatic weeds, e.g. Lemna minor, L. gibba, Najas minor and Utricularia inflexa and weeds having life cycles similar to that of rice, e.g. Cyperus difformis, Echinochloa colona, Echinochloa crus-galli, Paspalidium geminatum, Paspalum distichum. In our studied rice fields, typical aquatic weeds were missing but several other diagnostic species were those, characteristic of wet habitats.
Region is a complex variable used as surrogate for regional climate, but it is difficult to separate also the effect of crop and farming methods related to geographical gradient. The relationship between climate and vegetation has been well established. A bioclimate among other factors determines vegetation, region’s species distribution, irrigation needs and suitability of land for cultivation of specific crop types (Belda et al., 2014; Trewartha & Horn, 1980). Therefore, climatic classification of regions is important for vegetation distribution as each species has a climatic niche (Bailey, 2009; Feddema, 2005). In present study, region proved to be the most important factor. These findings corroborate those of Mahgoub (2019a, 2019b), who found that climate was the most important variable influencing weed vegetation for Nile Delta, although variables were not partialled out in those analyses.
Since the early 1960s, several large parts in the Egyptian deserts (Western, Eastern and Sinai) were converted into cultivated areas as a result of land reclamation projects (Salama et al., 2017). In the current study, the highest numbers of species (species richness per plot and species pool) were found in the old lands of the Nile Delta (El-Sharkiya) and Valley (Qena), as there are more favourable conditions for reproduction, germination and growth. These regions also have a long history of agriculture. The reclaimed areas of Natrun recorded the lowest species number due to the lack of water and poor soil (Abd El-Ghani et al., 2015). Unlike the Nile’s old cultivated fields, which are watered by an extensive network of irrigation systems through continuous flood irrigation from the river, reclaimed desert areas are often irrigated by recent irrigation techniques (e.g. sprinkle and drip) and mainly depend on underground water using pumps.
The vegetation in the agro-ecosystem of the reclaimed lands (as those of Natrun) consisted mainly of weeds similar to those growing in the cereals of the old lands, in addition to number of endemic xerophytes (e.g. Deverra tortuosa, Echinops spinosus, Farsetia aegyptia, Heliotropium digynum, Zygophyllum album, Z. coccineum, Z. simplex), which were able to persist with the reclamation processes. On the other hand, the availability of irrigation water in reclaimed desert creates suitable habitat for growing and flourishing of some rare desert plants.
Although a number of studies showed soil factors are very important determinants of weed species composition (Andreasen & Skovgaard, 2009; Dale et al., 1992; Fanfarillo et al., 2020; Nagy et al., 2018), they were not significant in our study. This may be attributed to the relatively homogeneous soil types in Egypt compared to other agricultural lands. Andreasen & Skovgaard (2009) already mentioned that it is sometimes difficult to find the true connection between soil properties and weed species composition, since every field has its own history with regard to crop rotation, soil cultivation, herbicide use, etc.
The only significant soil variable after partialling out of other variables was organic matter. This is related to old lands in the most fertile part of the country being rich in minerals and organic matter which enhance the vigorous growth of weeds compared to new arable areas. Additionally, old lands (Nile Delta and Valley) have silty and clayey soil texture, and new reclaimed lands have sandy soils. Crops in the reclaimed areas are also more similar; rice is not grown there as sandy soils cannot hold water during the flooded period.
The meaning of the season depends on the authors’ interpretation, and could represent the time of sampling (Šilc et al., 2009) or the sowing season (winter versus summer) (Pinke et al., 2012), so the caution is required when comparing the previous terms.
In our case, the season was defined based on sowing period for winter and summer crop types, and it was the least important variable for weed species composition. This contradicts results by Mahgoub (2021) for Nile delta where crop seasonality and diversification were more important than soil variables.