
Escherichia coli is a Gram-negative rod-shaped bacterium that is one of the most responsible for hospital-acquired infections, such as gastrointestinal infections, urinary tract infections, bloodstream infections, and neonatal meningitis (Riley 2020). The increasing prevalence of multidrug-resistant (MDR) and extensively drug-resistant (XDR) Gram-negative bacteria is indeed a concerning issue and has become a significant public health concern worldwide (Shen et al. 2018). Many Gram-negative bacteria produce extended-spectrum β-lactamase (ESBL) that can inactivate cephalosporins, monobactams, and penicillins. The production of ESBLs by bacteria poses a significant challenge in treating infections caused by these organisms since it renders many commonly used antibiotics less effective or even ineffective (Onduru et al. 2021). Meanwhile, carbapenemase-producing E. coli isolated from various countries contain unique carbapenemase genes, and their prevalence exhibits considerable global variation. In one study, eight carbapenemases, including KPC, IMP, NDM, VIM, OXA, FRI, GES, and IMI, were identified among the carbapenem-resistant E. coli (CRECs) analyzed, with NDM being the most predominant (52.15%), followed by OXA (30.09%) and KPC (14.72%) (Li et al. 2024b).
The resurgence of polymyxins as a last-resort treatment in the early 21st century was driven by the rising prevalence of MDR bacteria and the lack of new antibiotics (Rodríguez-Santiago et al. 2021). As a cationic polypeptide, it possesses a positive charge that enables it to bind to lipid A’s negatively charged phosphate group in the lipopolysaccharide (LPS) of bacterial outer membranes. This interaction leads to membrane destabilization, causing leakage of cytoplasmic contents and eventual cell lysis (Ling et al. 2020). However, as with other antibiotics, bacteria have developed resistance to polymyxins. Numerous studies have reported colistin-resistant bacteria’s emergence and global spread (Karki et al. 2021).
In 2015, Liu et al. (2016) discovered a plasmid-borne mcr-1 gene that confers polymyxin resistance. An initial report identified mcr-1 in E. coli and Klebsiella pneumoniae isolates from China’s food, animals, and humans (Rodríguez-Santiago et al. 2021). An initial report identified mcr-1 in E. coli and K. pneumoniae isolates from food, animals, and humans in China (Rodríguez-Santiago et al. 2021). The mobile colistin resistance (mcr) genes carried by plasmids have become increasingly widespread worldwide. As of now, 10 mcr variants (mcr-1 to mcr-10) have been identified in over 60 countries (Yin et al. 2022), with mcr-1, mcr-3, mcr-5, mcr-9, and mcr-10 variants being the most prevalent in mcr-positive E. coli (Dadashi et al. 2022). E. coli is the most common host for mcr gene, which are frequently carried by a variety of plasmids, including IncI2, IncHI2 and IncX4 (Shen et al. 2018; Xie et al. 2022). Research has indicated that the ease of horizontal transfer of mcr-family genes contributes to their global dissemination, increasing colistin tolerance in various bacterial species (Ling et al. 2020). Many studies have shown mcr-1-har-bouring E. coli belong to different sequence types, with a high prevalence of ST10, ST131, and ST155 (Wang et al. 2020; Shi et al. 2023; De La Cadena et al. 2023).
Countries across the world are now paying attention to the presence of mcr-1-positive E. coli producing ESBL, focusing on the environment, animals, water sources, and humans (Chotinantakul et al. 2022; Jamil et al. 2022; Lima et al. 2022; Shafiq et al. 2022; Bastidas-Caldes et al. 2023). However, very few studies have focused on the prevalence and characterization of ESBL-producing mcr-1-harbouring E. coli in clinical patients in Shandong Province; only a few studies have been focused on individuals living in rural areas (Bi et al. 2017; Berglund et al. 2018).
The current study aimed to investigate molecular epidemiology of polymyxin B resistance among ESBL-producing E. coli in the region and the presence of the mcr-1 gene. The findings could provide a theoretical evidence for the prevention and control of mcr-1 transmission.
Clinical strains identification. In this study, 668 non-duplicate ESBL-producing E. coli strains isolated from clinical samples at Shandong Provincial Hospital between January and December 2018 were collected for analysis. VITEK® 2 compact system (bioMérieux, France) and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF/MS) were used to identify the isolates. Twenty-four polymyxin B-resistant clinical strains were obtained by colistin agar test (agar plates containing 4 μg/ml of colistin). Then, polymyxin B’s minimum inhibitory concentration (MIC) was determined using the broth microdilution method and interpreted following the guidelines of the Clinical and Laboratory Standards Institute (CLSI 2023). The Ethics Committee of Shandong Provincial Hospital approved the clinical data collection of patients.
Antimicrobial susceptibility testing (AST). Antimicrobial susceptibility testing was conducted utilizing the VITEK® 2 compact system for ampicillin (AMP), ampicillin/sulbactam (SAM), piperacillin/tazobactam (TZP), cefazolin (CZO), cefotetan (CTT), cefatriaxone (CRO), ceftazidime (CAZ), cefepime (FEP), aztreonam (ATM), ertapenem (ETP), ertapenem (IPM), amikacin (AK), gentamicin (CN), tobramycin (TOB), ciprofloxacin (CIP), ciprofloxacin (LEV), nitrofurantoin (NIT), and trimethoprim/sulfamethoxazole (STX), except for polymyxin B (POL), which was analyzed by broth microdilution. E. coli ATCC® 25922™ was used as the reference strain. Results of the susceptibility assays were interpreted using the breakpoints provided by the Clinical and Laboratory Standards Institute (CLSI 2023).
Conjugation assay and confirmation of resistance genes. Sodium azide-resistant E. coli J53 was utilized as the recipient bacterium while mcr-1-harboring strains were employed as the donor strains. Transconjugants carrying the mcr-1 resistance genes were selectively grown on China-blue agar plates supplemented with 4 μg/ml polymyxin B and 100 μg/ml sodium azide. To confirm the transfer of the mcr-1 gene, polymerase chain reaction (PCR) was performed using primers mcr-1-F (5’-AGTCCGTTTGTTCTTGTGGC-3’) and mcr-1-R (5’-AGATCCTTGGTCTCGGCTTG-3’).
Pulsed-field gel electrophoresis (PFGE). The protocols were as described previously (Ma et al. 2020; Tang et al. 2022). 19 clinical strains and 17 transconjugants were selected and analyzed using S1-PFGE to reveal the number and approximate size of the plasmids. The XbaI-digested fragments of Salmonella enterica serovar Braenderup (H9812) were used as size markers. The XbaI-digested genomic DNA samples were subjected to pulsed-field gel electrophoresis (PFGE) using a CHEF Mapper® XA apparatus manufactured by Bio-Rad (USA) to assess the genetic relatedness across the clinical strains. Gel-J software, version 2.0 (Heras et al. 2015), was used to compare the pulsed-field gel electrophoresis (PFGE) patterns. The pulsotypes were categorized into clusters based on a similarity threshold of 80% (Chen et al. 2019).
Whole-genome sequencing (WGS) and analyses. Clinical strains and transconjugants were enriched overnight in Luria-Bertani broth at 37°C. Genomic DNA was extracted from the enriched cultures using a commercially available bacterial DNA extraction kit (Aidlab Inc., China). Both concentration and purity of the extracted genomic DNA were assessed using a NanoDrop™ 1000 Spectrophotometer (Thermo Fisher Scientific, Inc., USA), and the genomic DNA was subjected to next-generation sequencing using an Illumina Hiseq platform at the Novogene (Novogene Co., Ltd., China). Illumina sequences were assembled de novo using the SPAdes v3.10 (Prjibelski et al. 2020).
Multilocus sequence typing (MLST) was analyzed in silico by using Abricate software. FimH typing was performed using the Center for Genomic Epidemiology website tool. Antibiotic resistance genes were identified using the ResFinder database (Zankari et al. 2012). Plasmid replicon types were identified using the Plasmid Finder database (Carattoli et al. 2014).
Genomic DNA of ECO-POL-29 with the most common sequence type among these clinical strains, was sequenced using the Illumina HiSeq platform (Novogene Co., Ltd., China) and a PacBio RSII sequencer (Biozeron Biological Technology Co., Ltd., China). Paired-end short Illumina reads were utilized to correct long PacBio reads using the proovread software. Subsequently, the corrected PacBio reads were assembled de novo using SMARTdenovo software.
Gene annotation of the sequence was initially completed using the Rapid Annotation using Subsystem Technology (RAST) online annotation platform for prokaryotic genomes. The gene annotation results were further refined by manual curation using the BLAST tool. The mobile element annotation was performed using the online IS finder database (Siguier et al. 2006).
Phylogenetic analysis of isolates. Single-nucleotide polymorphism (SNP) calling was conducted with Snippy v3.1, and recombinant variants were filtered out with ClonalFrameML v1.0 (Lu et al. 2019). Recombination-free SNPs were used to construct maximumlikelihood phylogenetic trees with the RAxML software (Stamatakis et al. 2014). The tree file was visualized and the annotated information was edited using iTOL v6 (Letunic and Bork 2024). Serotyping was performed using the EcoSP website tool (Dong et al. 2023).
Analysis of the plasmid harboring mcr-1. The sequences of five plasmids with high sequence similarity (93–97% query coverage and 98.35–99.14% identity) to pECO-POL-29_mcr1 were obtained through the BLASTn analysis. Then these five plasmids were compared with pECO-POL-29_mcr1 by a BLAST Ring Image Generator tool (Alikhan et al. 2011), including pSLy21 (swine, United States) (GenBank accession no. CP016405), pAH01-2 (chicken, China) (GenBank accession no. CP055253), pRIVM_C029515_2 (Homo sapiens, Netherlands) (GenBank accession no. CP081339), pmcr1_IncI2 (Homo sapiens, China) (GenBank accession no. KU761326), and pEC26-mcr-1.9 (Homo sapiens, China) (GenBank accession no. MG946761).
Clinical isolates and antimicrobial susceptibility. Twenty-four polymyxin B-resistant strains were isolated from 668 non-duplicate ESBL-producing E. coli strains, with a positivity rate of 3.59%. Of the 668, 19 clinical strains carried the mobile colistin resistance gene mcr-1, with a positivity rate of approximately 2.8%. Clinical data of patients infected with these strains were shown in Table I.
Clinical data of patients infected with mcr-1 -positive Escherichia coli isolates producing extended-spectrum β-lactamase.
| Clinical isolates | Gender | Department | Specimen | Clinical diagnosis | Antimicrobial therapy |
|---|---|---|---|---|---|
| ECO-POL-4 | male | Intensive Care Unit | urine | aspiration pneumonia, ARDS | cefoperazone sodium/sulbactam sodium |
| ECO-POL-6 | female | Orthopedic Oncology and Joint Surgery Department | urine | recurrence of intradural teratoma | levofloxacin, levofloxacin sodium chloride, ceftazidime |
| ECO-POL-7 | female | Gastroenterology Department | bile | postoperative recurrence of pancreatic cancer | piperacillin/tazobactam |
| ECO-POL-8 | female | Emergency Department | secretion | acute purulent appendicitis | meropenem sodium/sulbactam sodium |
| ECO-POL-9 | female | Urology Department | urine | ureteral stone | – |
| ECO-POL-10 | female | Minimally Invasive Urology Department | urine | left renal abscess | cefotaxime |
| ECO-POL-15 | male | Minimally Invasive Urology Department | urine | post-prostatectomy for prostate cancer | cefotaxime |
| ECO-POL-17 | male | General Surgery Department | puncture fluid | post-pancreaticoduodenectomy | cefoperazone sodium/sulbactam sodium, imipenem/cilastatin |
| ECO-POL-18 | female | Urology Department | urine | bilateral ureteral stones | meropenem |
| ECO-POL-19 | female | Minimally Invasive Urology Department | urine | urinary retention | ceftriaxone sodium |
| ECO-POL-21 | male | Respiratory Department | sputum | severe pneumonia | fluconazole, cefoperazone sodium/sulbactam sodium, carbapenem |
| ECO-POL-22 | male | Intensive Care Unit | drainage fluid | open pelvic fracture with ARDS | imipenem/cilastatin |
| ECO-POL-23 | male | Intensive Care Unit | bronchoalveolar lavage fluid | septic shock | carbapenem, imipenem/cilastatin, daptomycin, meropenem + polymyxin B, vancomycin + fluconazole |
| ECO-POL-24 | male | Trauma and Orthopedics | secretion | traumatic hemorrhagic shock | cefotaxime, clindamycin hydrochloride |
| ECO-POL-25 | female | Spinal Surgery Department | puncture fluid | postoperative infection following lumbar spine surgery | penicillin, cefepime + levofloxacin, imipenem/cilastatin |
| ECO-POL-27 | male | Thoracic Surgery Department | sputum | post-chemotherapy for right upper lobe lung cancer | cefuroxime |
| ECO-POL-28 | male | Intensive Care Unit | pus | gastrointestinal bleeding, gastric ulcer | cefoperazone sodium/sulbactam sodium, linezolid |
| ECO-POL-29 | male | General Surgery Department | ascitic puncture fluid | post-appendectomy for appendiceal cancer, colonic lesion | imipenem/cilastatin |
| ECO-POL-30 | male | Trauma and Orthopedics | secretion | hemorrhagic shock | cefotaxime |
All 19 clinical strains were resistant to ampicillin, cefazolin, ceftriaxone, ciprofloxacin, levofloxacin, and polymyxin B. However, there was high sensitivity to piperacillin/tazobactam, with a sensitivity rate of 89.5% (17/19). The rate of aztreonam resistance was 84.2% (16/19). One of the 19 strains was resistant to ertapenem and imipenem. The rate of amikacin resistance was 26.3% (5/19). The rate of resistance to gentamicin was 73.7% (14/19). The resistance rates to nitrofurantoin and trimethoprim/sulfamethoxazole were 5.3% (1/19) and 73.7% (14/19), respectively.
Prior to conjugation, the recipient strain, J53, was sensitive to all the tested drugs. All transconjugants were sensitive to cefotetan, ertapenem, imipenem, amikacin, ciprofloxacin, levofloxacin, and nitrofurantoin but resistant to polymyxin B. Of the 17 transconjugants, seven were resistant to ampicillin, ampicillin/sulbactam, cefazolin, and ceftriaxone, and six were resistant to ceftazidime and cefepime. Sensitivity to piperacillin/tazobactam and tobramycin was high, with only one strain being intermediate or resistant out of the 17. Resistance rates to gentamicin and trimethoprim/sulfamethoxazole were both 17.6%. The results of the antibiotic susceptibility tests are presented in Table II.
Antibiotic susceptibility testing of Escherichia coli clinical strains, transconjugants, and the recipient bacterium J53AziR.
| Isolates | MIC (μg/ml) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMP | SAM | TZP | CZO | CTT | CRO | CAZ | FEP | ATM | ETP | IPM | AK | CN | TOB | CIP | LEV | NIT | STX | POL | |
| ECO-POL-4 | ≥ 32 | ≥ 32 | ≥ 128 | ≥ 64 | ≥ 64 | ≥ 64 | ≥ 64 | ≥ 64 | 16 | ≥ 8 | ≥ 16 | ≤ 2 | ≤ 1 | ≤ 1 | ≥ 4 | ≥ 8 | 32 | ≥ 320 | 8 |
| ECO-POL-6 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | 32 | ≥ 64 | ≤ 1 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | 40 | 4 |
| ECO-POL-7 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | ≤ 16 | ≥ 320 | 4 |
| ECO-POL-8 | ≥ 32 | 8 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≥ 4 | ≥ 8 | 32 | ≥ 320 | 4 |
| ECO-POL-9 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 4 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | 8 | ≥ 4 | ≥ 8 | ≤ 16 | ≥ 320 | 4 |
| ECO-POL-10 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≥ 4 | 4 | 32 | ≥ 320 | 4 |
| ECO-POL-15 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≥ 64 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≤ 20 | 8 |
| ECO-POL-17 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≥ 64 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 32 | ≤ 20 | 4 |
| ECO-POL-18 | ≥ 32 | ≥ 32 | 16 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | 32 | ≥ 64 | ≤ 0.5 | ≤ 1 | 16 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | ≤ 16 | ≥ 320 | ≥ 64 |
| ECO-POL-19 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | 32 | 32 | ≤ 0.5 | ≤ 1 | ≥ 64 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≥ 320 | 4 |
| ECO-POL-21 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≥ 320 | 4 |
| ECO-POL-22 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≤ 1 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≥ 64 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≥ 320 | 4 |
| ECO-POL-23 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≥ 320 | 4 |
| ECO-POL-24 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | 32 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | 8 | ≥ 4 | ≥ 8 | ≤ 16 | ≥ 320 | 4 |
| ECO-POL-25 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 4 | ≥ 64 | 32 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≥ 4 | ≥ 8 | 128 | ≤ 20 | 4 |
| ECO-POL-27 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 4 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | 8 | ≥ 4 | ≥ 8 | 64 | ≤ 20 | 4 |
| ECO-POL-28 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≤ 1 | 2 | 4 | ≤ 0.5 | ≤ 1 | ≥ 64 | ≥ 16 | ≥ 16 | ≥ 4 | ≥ 8 | 64 | ≥ 320 | 4 |
| ECO-POL-29 | ≥ 32 | 8 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 4 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≥ 4 | ≥ 8 | 32 | ≥ 320 | 4 |
| ECO-POL-30 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | 32 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | 8 | ≥ 4 | ≥ 8 | ≤ 16 | ≥ 320 | 4 |
| J4 | ≥ 32 | ≥ 32 | 64 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 8 |
| J6 | 8 | 8 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J7 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≥ 320 | 4 |
| J8 | 4 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J9 | 4 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 8 |
| J10 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J15 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 4 | 2 | 16 | ≤ 0.5 | ≤ 1 | ≤ 2 | 8I | ≥ 16 | ≤ 0.25 | ≤ 0.25 | 32 | ≤ 20 | 16 |
| J18 | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 8 |
| J19 | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J21 | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J22 | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J23 | 8 | 4 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J24 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≥ 64 | 8 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≥ 320 | 8 |
| J25 | ≥ 32 | 16 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | 16 | ≥ 64 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 4 |
| J27 | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 8 |
| J29 | 4 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 8 |
| J30 | ≥ 32 | ≥ 32 | ≤ 4 | ≥ 64 | ≤ 4 | ≥ 64 | ≥ 64 | 8 | ≥ 64 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≥ 16 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≥ 320 | 8 |
| J53AziR | 8 | ≤ 2 | ≤ 4 | ≤ 4 | ≤ 4 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 1 | ≤ 0.5 | ≤ 1 | ≤ 2 | ≤ 1 | ≤ 1 | ≤ 0.25 | ≤ 0.25 | ≤ 16 | ≤ 20 | 0.125 |
MIC – minimal inhibitory concentrations, AMP – ampicillin, SAM – ampicillin/sulbactam, TZP – piperacillin/tazobactam,
CZO – cefazolin, CTT – cefotetan, CRO – ceftriaxone, CAZ – ceftazidime, FEP – cefepime, ATM – aztreonam, ETP – ertapenem,
IPM – imipenem, AK – amikacin, CN – gentamicin, TOB – tobramycin, CIP – ciprofloxacin, LEV – levofloxacin,
NIT – nitrofurantoin, STX – trimethoprim/sulfamethoxazole, POL – polymyxin B
Plasmid carriage status of the clinical strains and transconjugants. S1-PFGE was used to analyze the number and approximate size of plasmids. Among the 19 strains of mcr-1-positive polymyxin B-resistant ESBL-producing E. coli, there were different numbers of plasmids, ranging from 2 to 6, with sizes roughly estimated between 20 kb and 300 kb. Seventeen transcon-jugants were obtained using conjugation assays. The transconjugants produced 1–4 plasmids with plasmid sizes ranging from approximately 20 kb to 300 kb (Table III).
The number, approximate size and replicon types of plasmids in 19 strains of mcr-1-positive polymyxin B-resistant Escherichia coli producing extended-spectrum β-lactamase and 17 transconjugants.
| Isolates | Plasmid numbers | Size (kbp) | Replicon types |
|---|---|---|---|
| ECO-POL-4 | 3 | ~ 110, ~ 65, ~ 45 | ColRNAI, IncFIA, IncFIC, IncFIC, IncX3 |
| ECO-POL-6 | 6 | ~ 210, ~ 140, ~ 110, ~ 104.5, ~ 78.2, ~ 65 | Col, ColRNAI, ColpVC, IncFIB, IncFII, IncHI1B, IncHI2A, |
| IncHI2, IncI1, IncI2, RepA, p0111 | |||
| ECO-POL-7 | 5 | ~ 220, ~ 138, ~ 100, ~ 65, ~ 54 | IncFIB, IncFIC, IncFII, IncHI2A, IncHI2, IncI2a, |
| RepA_1_pKPC-CAV1321 | |||
| ECO-POL-8 | 4 | ~ 140, ~ 100, ~ 33, ~ 21 | IncFIB, IncFIC, IncX4 |
| ECO-POL-9 | 3 | ~ 140, ~ 100, < 20 | Col156, Col440II, ColRNAI, IncB/O/K/Z, IncFIA, IncFIB1, |
| IncFIC, IncI2, IncX1, IncY | |||
| ECO-POL-10 | 4 | ~ 140, ~ 65, ~ 34, ~ 30 | Col, Col156, Col8282, IncFIB, IncFII, IncI2, IncX4 |
| ECO-POL-15 | 3 | ~ 265, ~ 65, ~ 45 | Col, IncFII, IncHI2A, IncHI2, IncN, IncX1, RepA |
| ECO-POL-17 | 3 | ~ 104.5, ~ 65, ~ 48 | Col440I, ColRNAI, IncI1, IncI2, IncN, IncR, IncX1 |
| ECO-POL-18 | 3 | ~ 110, ~ 104.5, ~ 65 | IncFIB, IncFIC, IncHI2A, IncHI2, IncI2, RepA |
| ECO-POL-19 | 3 | ~ 265, ~ 140, ~ 48 | Col156, Col440I, IncFIB, IncFIC, IncFII, IncHI2A, IncHI2, RepA |
| ECO-POL-21 | 2 | ~ 140, ~ 65 | Col156, IncFIB, IncFIC, IncI2 |
| ECO-POL-22 | 3 | ~ 180, ~ 110, ~ 65 | Col440II, ColRNAI, IncFIB, IncFIC, IncI2, IncI |
| ECO-POL-23 | 2 | ~ 140, ~ 65 | Col156, IncFIB, IncFIC, IncI2 |
| ECO-POL-24 | 2 | ~ 140, ~ 65 | IncFIB, IncI2 |
| ECO-POL-25 | 3 | ~ 140, ~ 95, ~ 65 | IncB/O/K/Z, IncFIB, IncI2, p0111 |
| ECO-POL-27 | 6 | ~ 140, ~ 130, ~ 95, ~ 70, ~ 65, ~ 43 | Col(MG828), ColE10, ColRNAI, IncB/O/K/Z, IncFIB, IncFIC, |
| IncFII, IncI2, IncX1, p0111 | |||
| ECO-POL-28 | 2 | ~ 90, ~ 28 | Col, Col156, Col440I, ColRNAI, IncFIA, IncFIB, IncI, IncX1 |
| ECO-POL-29 | 4 | 128.293, 83.176, 62.869, 30.619 | IncFIB/IncFIC, IncFIC, IncI2, IncX4 |
| ECO-POL-30 | 2 | ~ 130, ~ 60 | IncFIB, IncI2 |
| J4 | 1 | ~ 65 | IncI2 |
| J6 | 1 | ~ 65 | IncI2 |
| J7 | 4 | ~ 103, ~ 100, ~ 65, ~ 60 | IncI2, IncFIB, IncFII, IncFIC |
| J8 | 2 | ~ 100, ~ 33 | IncFIB, IncX4 |
| J9 | 1 | ~ 65 | IncI2 |
| J10 | 3 | ~ 65, ~ 33, ~ 30 | IncI2, IncX4 |
| J15 | 1 | ~ 255 | IncHI2, RepA, IncHI2A, IncN |
| J18 | 1 | ~ 65 | IncI2 |
| J19 | 1 | ~ 255 | IncHI2A, IncHI2, RepA |
| J21 | 1 | ~ 65 | IncI2 |
| J22 | 1 | ~ 65 | IncI2, ColRNAI |
| J23 | 1 | ~ 65 | IncI2 |
| J24 | 2 | ~ 140, ~ 65 | IncI2, IncFIB |
| J25 | 1 | ~ 65 | IncI2 |
| J27 | 1 | ~ 65 | IncI2 |
| J29 | 2 | ~ 65, ~ 33 | IncI2, IncX4 |
| J30 | 2 | ~ 140, ~ 65 | IncI2, IncFIB |
Molecular typing and genotyping of the phylogenetic groups. Of the 19 strains of mcr-1 -positive polymyxin B-resistant E. coli-producing ESBL, there were 14 sequence types (STs). The most common ST was ST155 (n = 4, 21%), followed by ST162 (n = 2), ST393 (n = 2), ST457, ST88, ST450, ST617, ST744, ST354, ST359, ST93, ST224, ST69, and ST167. ECO-POL-21 and ECO-POL-23 shared the same serotype (O29:H12) and ST. Similarly, ECO-POL-24 and ECO-POL-30 had the same serotype (O15: H1) and ST (Fig. 1).
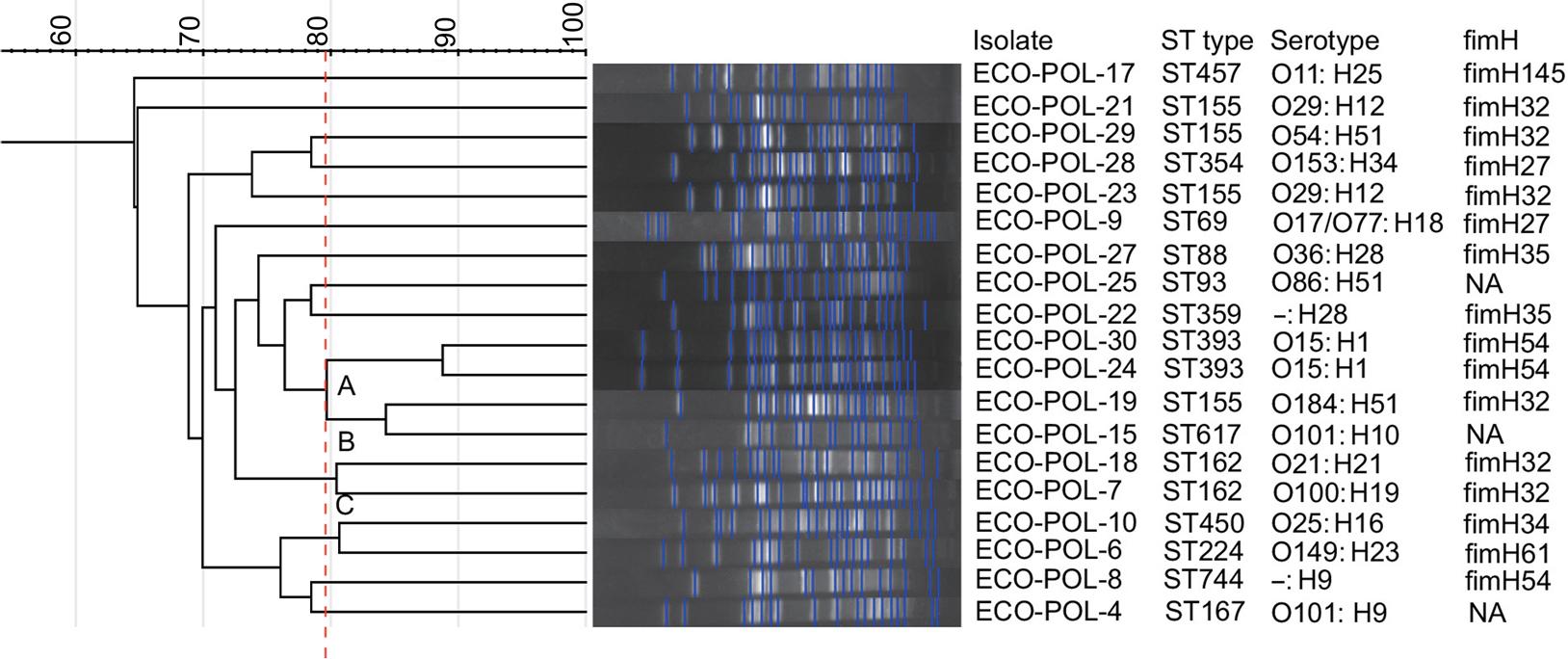
A dendrogram was constructed using the similarity of pulsed-field gel electrophoresis (PFGE) patterns across the 19 strains of mcr-1 -positive polymyxin-B-resistant extended-spectrum β-lactamase-producing Escherichia coli. Names of the isolates, sequence types, serotypes, and fimH types are displayed on the right side.
Based on the PFGE profiles, only three distinct clusters, identified as clone groups A to C, were observed, indicating a lower homology among the 19 isolates. According to the PFGE profile and MLST analysis, there was homology between strains 24 and 30, both of which belong to ST393. The ST162 isolates showed a high degree of homology. The 19 strains were distributed across different branches, suggesting considerable genetic diversity among the strains, with variations in the overall SNPs. According to branch length, ECO-POL-24 and ECO-POL-30 exhibited the most closely related evolutionary lineage (Fig. 2). The predominant replicon type of transconjugants was IncI2 (n = 14) followed by IncFIB (n = 4) (Table III).
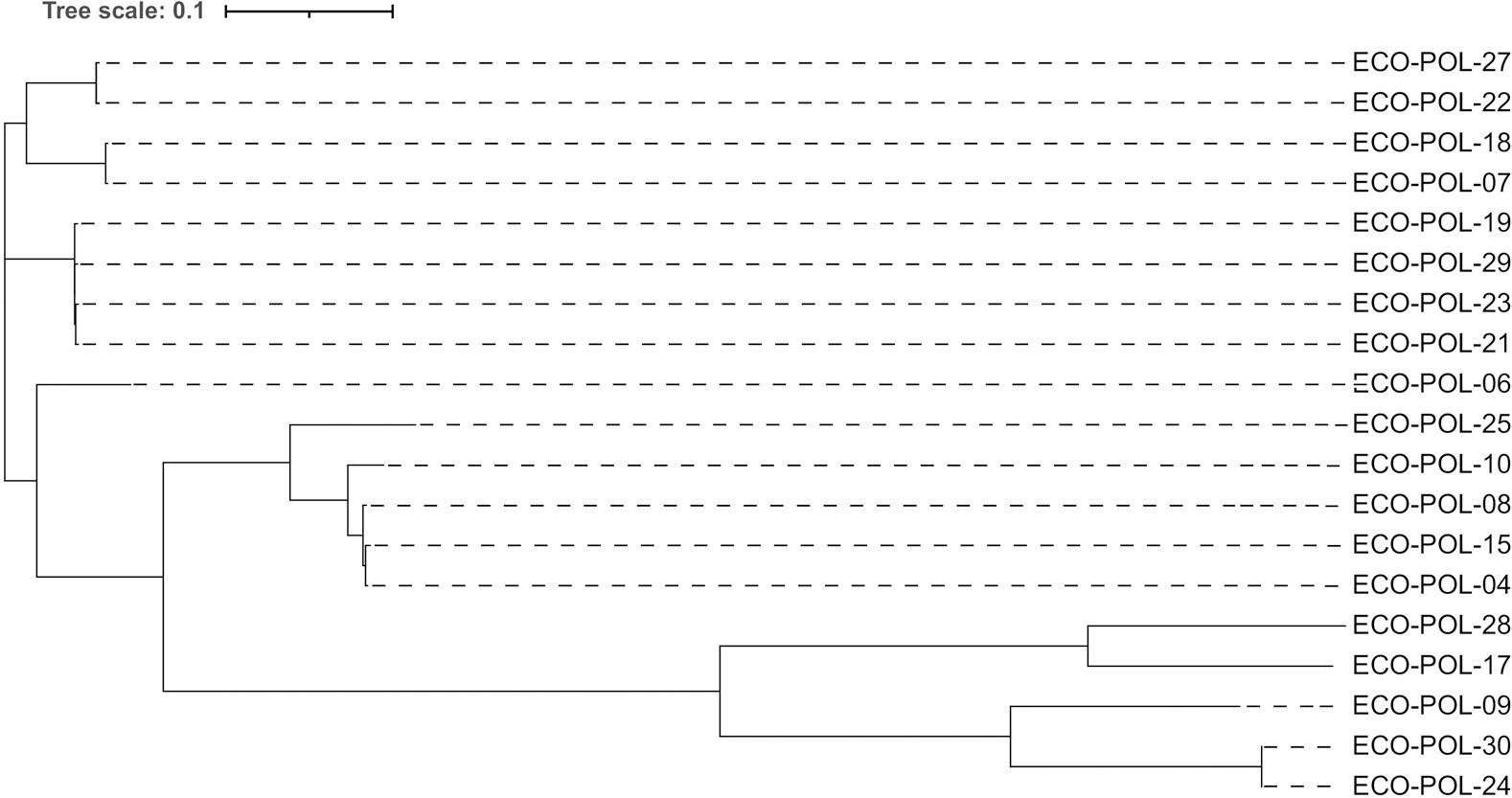
Phylogenetic tree distribution of 19 strains of mcr-1-positive polymyxin-B-resistant extended-spectrum β-lactamase-producing Escherichia coli.
Distribution of drug-resistance genes in clinical strains and transconjugants. Next-generation sequencing and analysis revealed that all 19 clinical strains harbored mcr-1. No other mcr variants were detected. Additionally, the drug resistance genes bla
The carriage rate of bla
Regarding aminoglycoside resistance, 14 isolates harbored APH(6)-Id, followed by APH(3’’)-Ib (13/19), AAC(3)-IV (8/19), and aadA2 (8/19). ECO-POL-19 carried the most aminoglycoside resistance genes, totaling to 8. Bla
DNA sequencing and PCR results showed that all 17 transconjugants carried the drug resistance gene mcr-1. ESBL genes such as bla
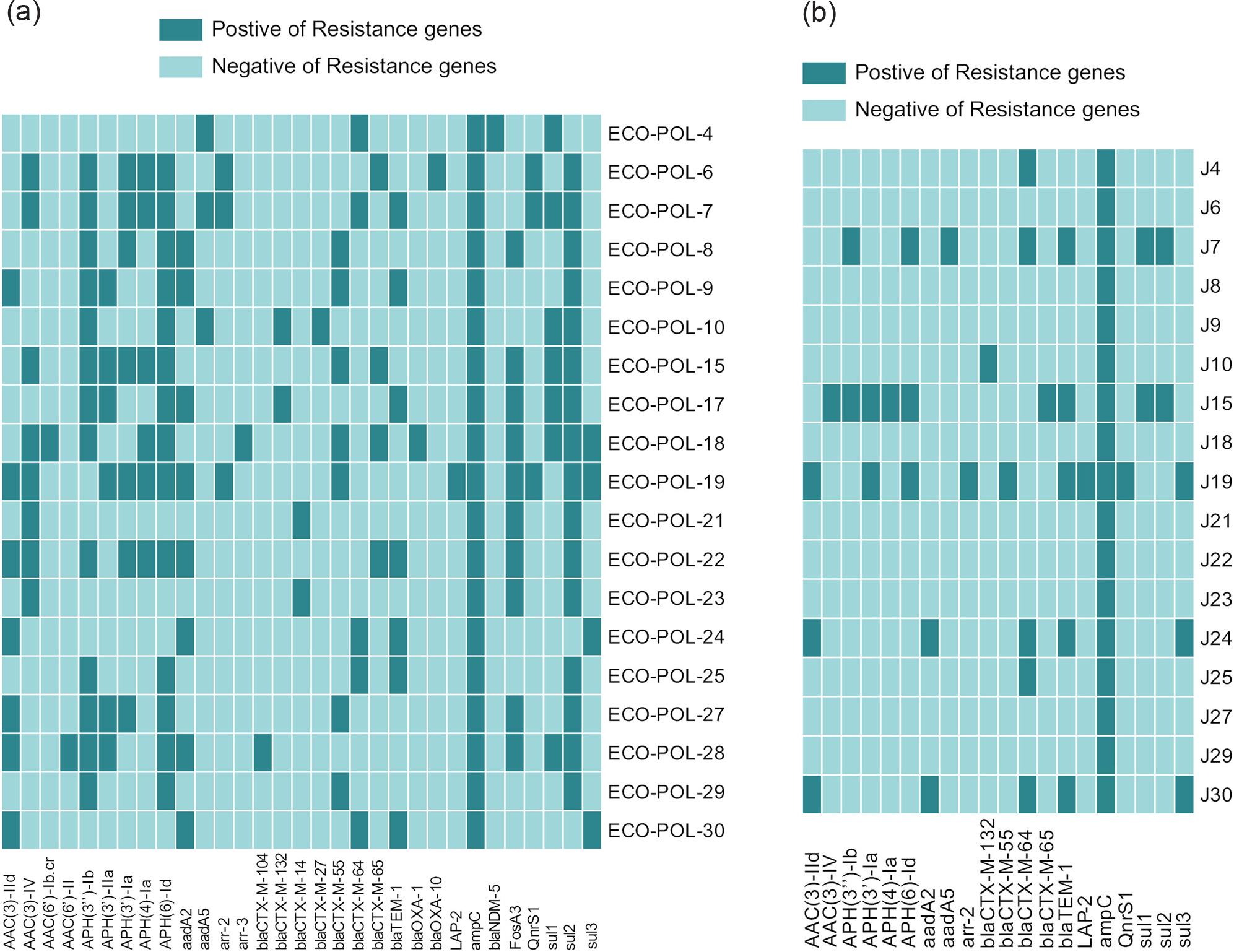
a) Detection of resistance genes in 19 mcr-1-positive clinical strains, b) detection of resistance genes in 17 transconjugants.
Analysis of the plasmid harboring mcr-1. The clinical strain ECO-POL-29 was subjected to wholegenome sequencing (WGS). One chromosome and four plasmids were present in the ECO-POL-29. The plasmid pECO-POL-29_mcr1 was an IncI2-type plasmid with a GC content of 42.7% and 88 open reading frames and the only resistance gene in this plasmid was mcr-1. There was no insertion sequence near mcr-1. The Type IV secretion system (T4SS) was detected in pECO-POL-29_mcr1. A transconjugant harboring pECO-POL-29_mcr1 was successfully obtained using the conjugation assay. Based on the BLASTn results for pECO-POL-29_mcr1, five plasmids from different countries, regions, and hosts were identified and selected. All five plasmids originated from E. coli and shared the same plasmid type IncI2. As indicated in Fig. 4, the main differences across the plasmids were distributed in region 1 (ranging from 7.32 kb to 10.19 kb) and region 2 (ranging from 57.8 kb to 58.9 kb). These regions contain genes associated with proteins of unknown function (Fig. 4).
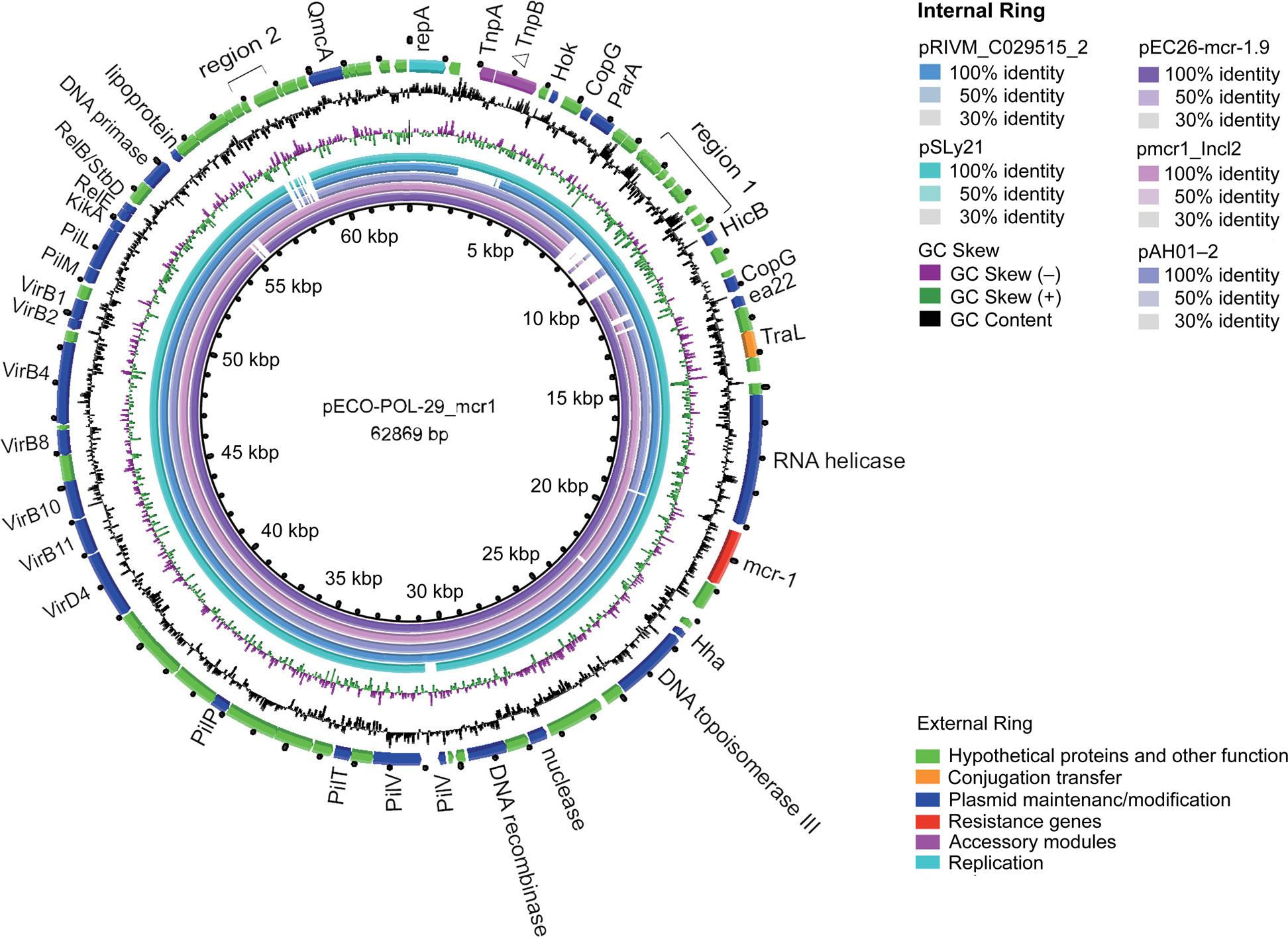
The external ring is the schematic representation of plasmid pECO-POL-29_mcr1 (GenBank accession number PP721192). Genes are marked with different colors based on their functional annotations. The internal five rings show a comparative analysis of mcr-1-harboring plasmids with pECO-POL-29_mcr1, including pSLy21 (sky blue), pAH01-2 (light purple), pRIVM_C029515_2 (light blue), pmcr1_IncI2 (light purple pink), and pEC26-mcr-1.9 (purple) (constructed using BRIG).
Nucleotide sequence accession numbers. The raw reads of all 19 isolates have been deposited in GenBank (BioProject PRJNA1104118). The complete sequence of pECO-POL-29_mcr1 has been deposited in GenBank under accession number PP721192.
E. coli is a significant causative agent of severe clinical bacterial infections with increasing levels of antibiotic resistance(Arafi et al. 2023; Ibrahim et al. 2023). Polymyxins have re-entered our field of view as the last line of defense against infections caused by multidrugresistant (MDR) Gram-negative bacteria (Rhouma et al. 2023). Since the first detection of the mcr-1 gene in 2015, ten mcr variants have been identified to date. Among those, mcr-1 remains the most dominant in clinical isolates (Nang et al. 2022; Rodríguez-Santiago et al. 2022).
Among the 668 strains of ESBL-producing E. coli collected in this study, 24 were multidrug-resistant strains, resistant to polymyxin B, with a resistance rate of 3.59%. According to a review published in 2022, the global prevalence of polymyxins resistance was 1.6% (Binsker et al. 2022). The result of this study was higher than that of the previous review. Among the 668 strains, 19 carried the colistin resistance gene mcr-1, with a positivity rate of approximately 2.8%. According to a tudy in 2016, mcr-1 was detected in only two isolates (4.2%) of 48 ESBL-producing E. coli from a hospital in France (Robin et al. 2017). In a study on healthy children in Central South China, the prevalence of mcr-1 was 0.84%, with one positive sample of over 118 ESBL-producing E. coli (Liu et al. 2022). The 2.8% positivity rate was significantly lower than that of environmental strains (5.43%) (Fan et al. 2020).
It would be worth noting that the mcr gene was associated with other antimicrobial resistance genes, such as ESBLs, carbapenemases, fluoroquinolons, and fosfomycins, making the treatment of infections caused by these bacteria even more difficult (Tian et al. 2020; Rodríguez-Santiago et al. 2022; Di Francesco et al. 2023; Szmolka et al. 2023). In the present study, all 19 mcr-1-positive clinical strains carried bla
Many studies have indicated that the STs of E. coli isolates carrying mcr-1 gene exhibit a high degree of diversity (Liu et al. 2018; Zhang et al. 2019; Vu Thi Ngoc et al. 2022). The STs in this study were also characterized by such diversity. According to a previous study, the most common ST among the isolates carrying the mcr-1 gene was ST10, also commonly found in E. coli isolates carrying ESBL from humans and animals (Shen et al. 2018). In our study, ST155 was the most common sequence type, which was consistent with the description of a previous study (Wang et al. 2020).
The mcr gene carried by plasmids is present in various types of plasmids, such as IncI2, IncHI2, IncX4, IncP, IncF, and uncommon IncY (Li et al. 2022; Protonotariou et al. 2022; Rodríguez-Santiago et al. 2022). The IncI2 plasmid is considered the most successful vector for horizontal dissemination of the mcr-1 gene, according to previous studies (Al Mana et al. 2022; He et al. 2022). However, in the Shanghai region, between 2012 and 2015, the predominant plasmid type among mcr-1-positive E. coli was IncX4, followed by IncI2. After 2015, IncI2 gradually surpassed IncX4 (Xie et al. 2022). Nevertheless, this result differed from those of the main types detected in Hong Kong (IncI1, IncX4, and IncN) (Chan et al. 2018). In the present study, the most common replicon types of transconjugants associated with mcr-1 were IncI2 and IncFIB.
Whole-genome sequencing of ECO-POL-29 revealed that the mcr-1 gene was located on an IncI2-type plasmid approximately 62.8 kb in size, and the plasmid did not carry any other resistance gene. The transconjugant carried only the mcr-1 gene. Additionally, the bla
The genetic background of mcr-1 was found to be highly diverse. Many studies have indicated that the mobilization of the mcr-1 gene is primarily associated with transposition mediated by ISApl1 (Shen et al. 2018; Ding et al. 2021; Zhou et al. 2021). ISApl1 is considered an element that can mobilize genes on plasmids and integrate them into the chromosome (Al Mana et al. 2022). It was first described in Acineto-bacillus pleuropneumoniae as a part of the IS30 family (Tegetmeyer et al. 2008). Previous studies have shown that the ISApl1-mcr-1-pap2-ISApl1 and ISApl1-mcr-1-pap2 structures carrying mcr-1 are involved in ISApl1 (Xie et al. 2022; Li et al. 2024a). Shen et al. showed that most mcr-1 genes are not adjacent to IS elements (Shen et al. 2018). A previous study had reported that 68.3% of mcr-1-positive E. coli strains completely lost ISApl1 (Huang et al. 2021). After analyzing our sequencing results, we found that the mcr-1 gene was not adjacent to ISApl1. Our findings collectively suggested that ISApl1 is not an essential mobile element for the transfer of mcr-1.
According to the data compiled in one study, ISApl1 is not commonly found in IncI2 and IncX4 plasmids (Li et al. 2024a). This description aligned with the genetic environment of pECO-POL-29_mcr1, since the plasmid type of pECO-POL-29_mcr1 is IncI2. Based on the result of pECO-POL-29_mcr1 genomic analysis, there was no mobile element surrounding mcr-1, suggesting that the mechanism by which the plasmid acquired mcr-1 would require further study. The T4SS on the pECO-POL-29_mcr1plasmid plays a crucial role in the spread of the mcr-1 gene, enabling its transmission as a plasmid.
In conclusion, our study uncovered the presence and spread of mcr-1-carrying ESBL-producing E. coli in the region and provided their genomic characteristics. As we increasingly recognise the importance of “One Health” taking more measures to monitor and stem the prevalence and transmission of mcr-1 would be essential.