The serine protease thrombin (coagulation factor IIa) functionally links blood clotting with inflammatory signaling [1], a concept captured by the term “thrombo-inflammation”. Thrombin directly stimulates cytokine production in various cell types via protease-activated receptors (PAR), independent of its hemostatic action [1, 2]. In cardiac fibroblasts, thrombin for example promotes the production of interleukin (IL)-1β and IL-6 [3, 4]. In the post-infarcted heart, neutrophil-expressed PAR4 contributes to cardiomyocyte apoptosis in the acute phase, but in the chronic phase critically regulates resolution and repair [5]. IL-6 also exerts highly context-dependent action on myocardial inflammation and resolution, healing and repair, with cardiomyocytes serving as a relevant intrinsic source [6, 7]. IL-6 is one of the most prominent transcriptional targets downstream of IL-β [8], the key effector cytokine of the NLRP3 (NOD-, LRR- and pyrin domain-containing protein 3) inflammasome. This signaling platform was initially characterized in myeloid cells but has also been demonstrated to be functionally active in cardiomyocytes [9, 10]. Cardiomyocytes express the prototypical thrombin receptor PAR1 [11-13] and release modest amount of IL-1β in response to thrombin [14]. Thrombin-stimulated secretion of IL-6 has to date not been reported in cardiomyocytes. One study did show IL-6 release by H9c2 rat cardiomyoblasts, but this was in cells subjected to hypoxic injury followed by exposure a pathologically high concentration of thrombin (5 U/mL) [15]. We therefore sought to establish if cardiomyocytes produce IL-6 in response to the more physiological thrombin concentration of 1 U/mL) [16], without prior hypoxia, and how this relates to cardiomyocyte-neutrophil cross-talk.
The mouse atrial cardiomyocyte HL-1 cell-[17] and Claycomb culture medium were purchased from Merck KGaA/Sigma-Aldrich (Taufkirchen, Germany). Cells were maintained under standard culture conditions as described [9]. HL-1 cells at passage 7-10 were seeded in Claycomb medium in 6-well culture plates at a density of 1x106/well. Two days after seeding, at a density of approximately 80-90%, HL-1 were stimulated with 1 U/mL of human α-thrombin (American Diagnostica GmbH, Pfungstadt, Germany), 20 ng/mL oxidized human serum albumin (oxAlb, Biocat GmbH, Heidelberg, Germany) or phosphate-buffered saline (PBS) vehicle for 24 h.
Peripheral blood was obtained from healthy human donors with informed consent and approval by the institutional ethics committee (20-9697-BO) and in accordance with the Declaration of Helsinki. ethylenediaminetetraacetic acid (EDTA)-anticoagulated blood was immediately processed for preparation of neutrophils and autologous platelet-rich plasma (PRP). PRP was obtained by centrifugation of whole blood at 1650 x g, 5 min, and stored at 37°C until use (resuspension and stimulation of concurrently isolated neutrophils). Neutrophils were isolated using PolymorphPrepTM as instructed by the manufacturer (PROGEN, Heidelberg, Germany). Neutrophil purity of >90% was verified using the DxH 520 Hematology Analyzer as per manufacturer instruction (Beckman Coulter GmbH, Krefeld, Germany). Neutrophils resuspended in autologous PRP were divided into two tubes, each containing 20-30 x103 neutrophils in 100 μL, and stimulated for 3 h in a humidified 37°C incubator with PBS vehicle or with thrombin plus 1.25 mM GPRP amide (H-Gly-Pro-Arg-Pro-NH2, Bachem AG, Bubendorf, Switzerland) to prevent coagulation of plasma. Following centrifugation (460 x g, 5 min), cleared supernatants were used for stimulation, a fraction was stored at -80°C for later analysis.
Freshly conditioned PRP from control or thrombin-stimulated neutrophils was diluted 1:4 with Claycomb HL-1 cell culture medium. PRP from thrombin-stimulated neutrophils was supplemented with 50 μM of the CD36 inhibitor Sulfo-N-Succinimidyl Oleate Sodium (SSO, Merck) or vehicle (0.1% dimethylsulfoxide, DMSO); PRP from control neutrophils was also supplemented with 0.1% DMSO. PRP were then added to HL-1 cells for 24 h.
Male C57Bl/6J were obtained from Janvier Labs (Le Genest St. Isle, France) and housed at 22°C, 50-60% relative humidity and a 12h light/dark cycle with free access to water. At 8 weeks of age, mice were randomized to receive standard chow or a high fat diet (HFD, #S7200-E010, ssniff Spezialdiäten GmbH, Soest, Germany) for 8 weeks. Over this time, HFD-fed mice were confirmed to develop progressive adiposity, insulin resistance, glucose intolerance [18] and myocardial NLRP3 inflammasome activation [3], compared to chows. At age 16 weeks, mice were anesthetised by intraperitoneal injection of ketamine/xylazine (100/5 μg/g body weight). Once anesthesia was confirmed by loss of toe-pinch reflexes, mice were sacrificed by cervical dislocation. The body cavity was carefully opened, hearts were removed and rinsed in ice-cold PBS. The septum-free LV was dissected and snap-frozen in liquid nitrogen. The study was approved by the local animal ethics authority (Landesamt für Natur, Umwelt und Verbraucherschutz, LANUV Nordrhein-Westfalen, Az. 81-02.04.2017.A458 and 81-02.04.2022.A297) and was performed in accordance with the ARRIVE and IMPROVE guidelines.
Snap-frozen cell supernatants and LV lysed in Kranias buffer [3] were assessed using the following kits: OxiselectTM Myeloperoxidase (MPO) Chlorination Activity Assay (Catalog Nr. STA-803) and OxiselectTM Advanced Oxidation Protein Products (AOPP) Assay (Catalog Nr. STA-318), both from Cell Biolabs (Heidelberg, Germany); Human IL-6 Quantikine ELISA Kit (Catalog Nr. D6050B) and Mouse IL-6 Quantikine ELISA Kit (Catalog Nr. M6000B-1), both from R&D Systems, Wiesbaden, Germany. IL-6 mRNA was determined by qPCR relative to the housekeeper gene ribosomal 18S using Qiagen Validated Quantitect Primer Assay (Qiagen, Hilden, Germany) as described [3, 4]. IL-6 and CD36 total protein levels were determined in LV lysates relative to g-tubulin using the LI-COR Odyssey platform as described [19]. Monoclonal antibodies against IL-6 and CD36 were obtained from Santa Cruz Biotech (Heidelberg, Germany), primary antibody against g-tubulin was from ThermoFisher Scientific (Waltham, MA, USA). Infrared-coupled secondary antibodies were from LI-COR Biosciences (Bad Homburg, Germany).
Data show mean ± standard deviation (SD), as fold of control or as absolute values where appropriate. Each n represents one individual mouse or one independent cell culture experiment, performed with single wells per condition. Analysis by qPCR, immunoblot or ELISA were performed with three technical replicates, the mean of which was taken as n=1. Statistical differences between two groups were determined by the Mann-Whitney U-test. For comparison amongst three groups, the Kruskall-Wallis one-tailed ANOVA on ranks was performed, with Dunn’s multiple comparison procedure applied as appropriate. GraphPad Prism version 10 was used to generate figures and perform statistical tests. P<0.05 was accepted as statistically significant.
Thrombin applied directly to HL-1 cardiomyocytes did not influence either IL-6 transcript levels or its secretion (Fig. 1A, B). Oxidised albumin by contrast increased Il6 mRNA to 2.2±0.9 fold of control (P<0.05, Fig. 1C)) and the amount of secreted IL-6 from 24±6 to 47±14 ng/mL supernatant (P<0.05, Fig. 1D). To determine if thrombin can indirectly stimulate cardiomyocyte IL-6 production via neutrophil secretion products, human neutrophils were stimulated with and without thrombin for 3h in autologous PRP. Conditioned medium from thrombin-stimulated neutrophils contained more active MPO than controls (364±62 versus 221±40 mU/mL, P<0.05, Fig. 2A) and formed more advanced oxidation protein products (AOPP; 125±7 ng/mL versus 90±16, P<0.05, Fig. 2B). Exposure of HL-1 cells to conditioned medium from thrombin-stimulated neutrophils increased Il6 mRNA in HL-1 to 1.9±0.7 fold compared to conditioned medium from control neutrophils (P=0.1, Fig. 2C). In the presence of the CD36 inhibitor SSO, IL-6 transcript levels were reduced to 1.4±0.3 fold of control but this effect was not statistically significant (Fig 2C). Secreted IL-6 amounted to 9.4±4 ng/mL in HL-1 cells exposed to conditioned medium from control neutrophils; with conditioned medium from thrombin-stimulated neutrophils this increased to 21.6±5 ng/mL (P<0.05, Fig. 2D). SSO restored IL-6 secretion nearly to control levels (11.2±5 ng/mL, P=0.08 versus thrombin-stimulated, Fig. 2D). The CD36 inhibitor alone was without effect on either parameter (data not shown).
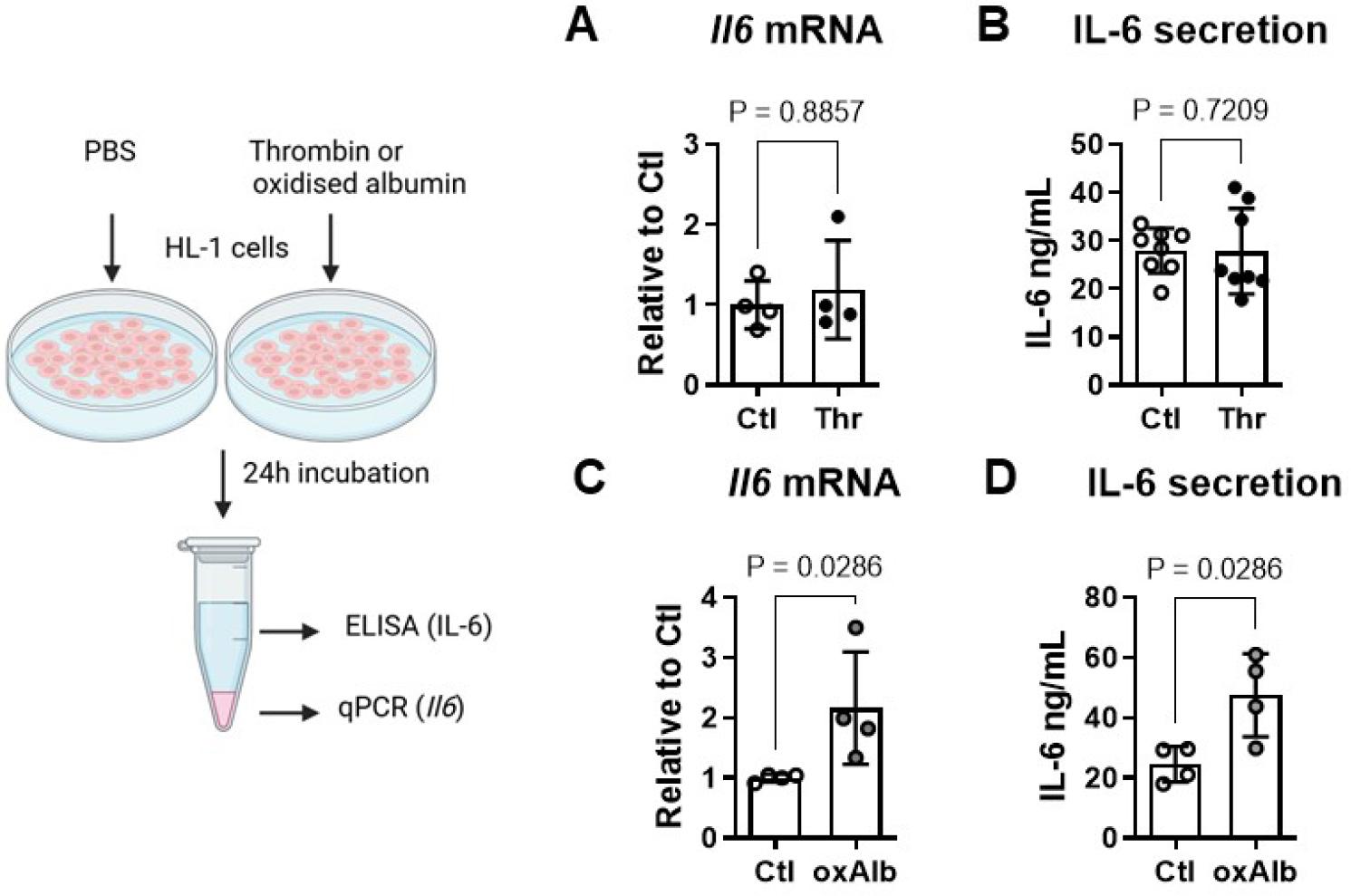
Thrombin does not directly regulate IL-6 in cardiomyocytes at physiological concentrations. (A) Exposure of HL-1 atrial cardiomyocytes to thrombin (Thr, 1 U/ml, 24 h) did not influence either Il6 mRNA expression or (B) IL-6 secretion, compared to control (Ctl, PBS vehicle). (C) Oxidized albumin (oxAlb, 20 ng/mL, 24 h) by contrast increased both Il6 mRNA and (D) secreted IL-6 compared to PBS control in HL-1 cardiomyocytes. Data show mean ± SD.
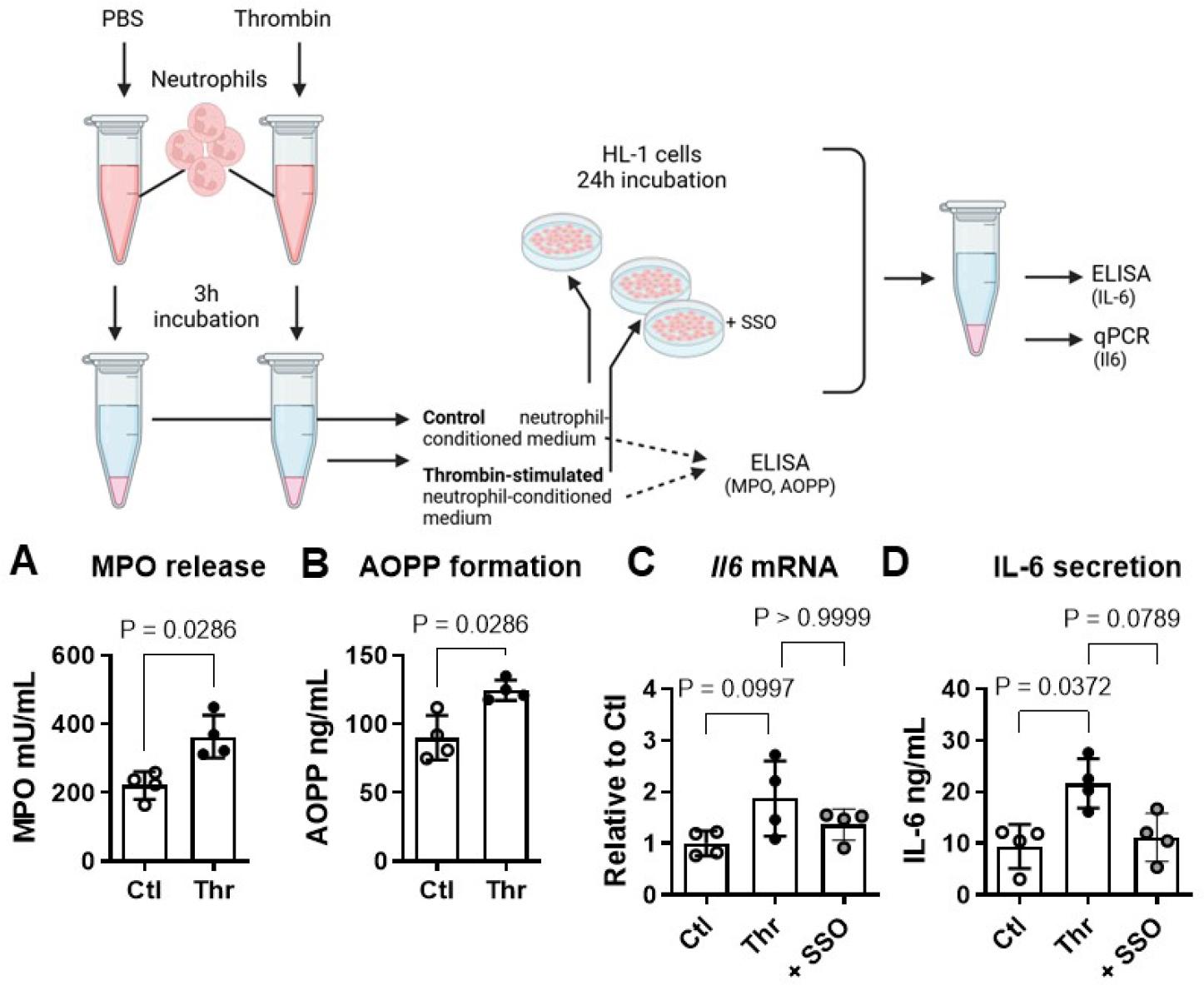
Thrombin indirectly regulates cardiomyocyte IL-6 via neutrophil-derived oxidation products. (A) In the presence of thrombin (Thr, 1 U/mL, 3h), human neutrophils in autologous platelet-rich plasma (PRP) produced more extracellular myeloperoxidase (MPO) activity and (B) Advanced Oxidation Protein Products (AOPP) compared to unstimulated control (Ctl) neutrophils. (C) Exposure of HL-1 atrial cardiomyocytes to PRP conditioned with thrombin-stimulated neutrophils increased Il6 transcript levels and (D) IL-6 secretion compared to PRP conditioned with control neutrophils. In the presence of the CD36 inhibitor SSO both effects were attenuated. Data show mean ± SD.
LV from mice fed a high fat diet (HFD) exhibited higher levels of MPO activity than LV from chow-fed mice (644±120 versus 445±139 mU/mL, P<0.05, Fig. 3A). AOPP content was 381±105 and 221±124 ng/ml respectively in LV from HFD-versus chow-fed mice (P=0.06, Fig. 3B). LV from HFD-mice also showed higher expression of CD36 total protein (1.8±08 fold of chow, P=0.06, Fig. 3C) and of IL-6 total protein (1.5±0.5 fold of chow, P<0.05, Fig. 3D).
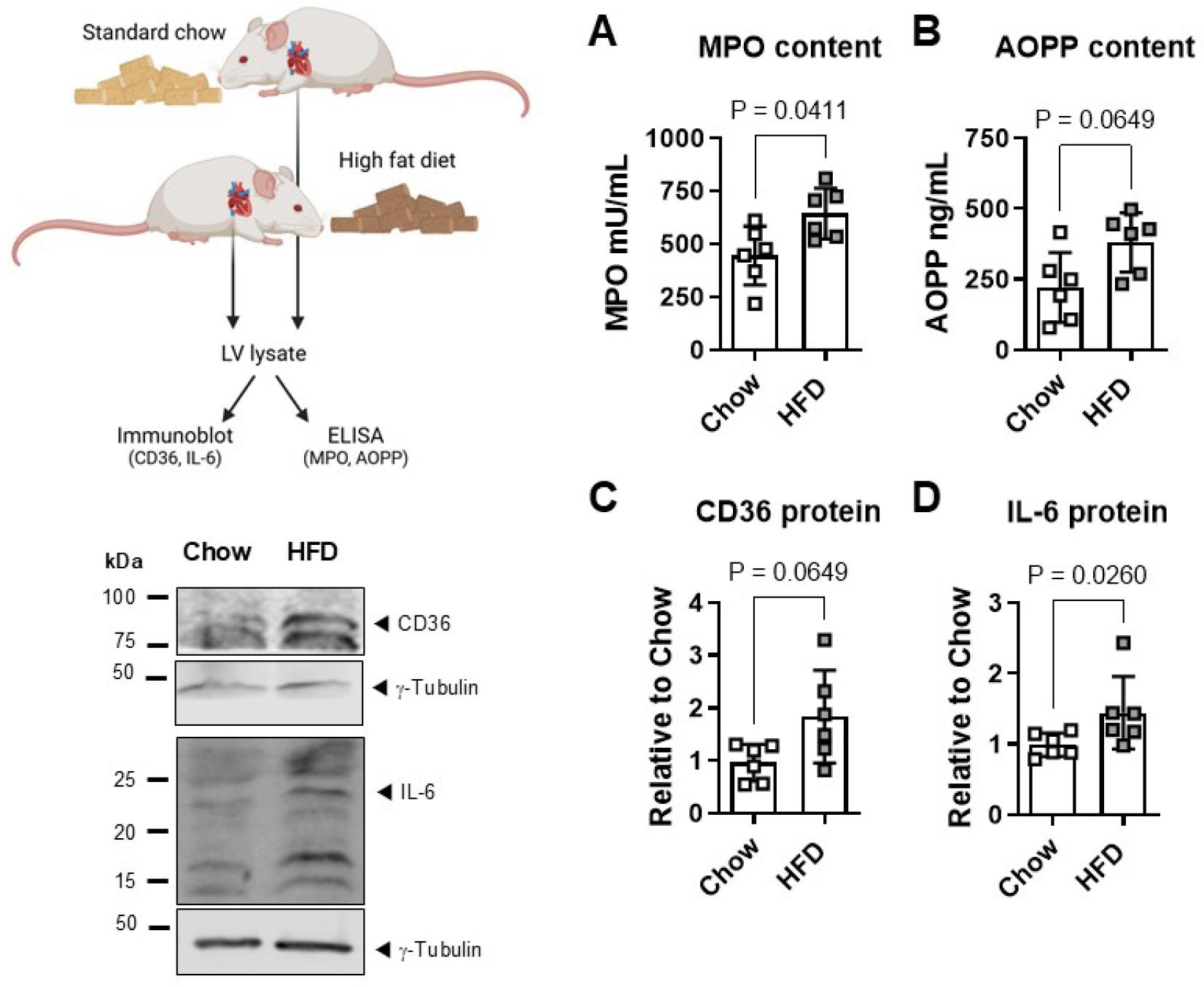
High fat diet increases myocardial IL-6. (A) Left ventricular (LV) lysates from mice fed a high fat diet (HFD) displayed higher myeloperoxidase (MPO) activity and (B) Advanced Oxidation Protein Products (AOPP) than LV from chow-fed controls. (C) Immunoblot also showed more abundant CD36 and (D) IL-6 total protein expression in LV myocardium of HFD-versus chow-fed mice. Immunoblots were normalized to γ-tubulin. Data show mean ± SD. This image was created with Biorender.com.
Thrombo-inflammation, a term that conceptually links the coagulant cascade with inflammatory and immune responses, is a critical feature and candidate platform for therapeutic intervention in cardiovascular diseases [20]. Thrombin is able to directly influence the phenotype and function of cardiac fibroblasts and cardiomyocytes in a hemostasis-independent manner via protease-activated receptors (PAR) [2-4, 13, 14]. In HL-1 atrial cardiomyocytes, thrombin-activated PAR4 elicits the expression, maturation and release of IL-1β [14]. Whether IL-6 is also regulated in response to thrombin is not known. We have noted increased Il6 mRNA in thrombin-stimulated mouse cardiac fibroblasts under high glucose conditions [4], while others have reported that a supra-physiological concentration of thrombin (5 U/mL) stimulates IL-6 release from hypoxia-primed rat H9c2 rat cardiomyoblasts [15]. We here show that a more physiologically relevant thrombin concentration (1 U/mL), reflecting what is released during the clotting process [16], does not elicit IL-6 transcription or secretion in unprimed HL-1 atrial cardiomyocytes, unlike what we previously saw in terms of IL-1β [14]. To confirm that the HL-1 atrial cardiomyocytes are in fact able to produce IL-6 we exposed HL-1 cells to oxidised albumin (oxAlb), a prototypic Advanced Oxidation Protein Product (AOPP) that is taken up into cells via the CD36 scavenger receptor and initiates inflammatory and redox stress [21-24]. This reactive species did increase IL-6 transcript levels and IL-6 release in HL-1 atrial cardiomyocytes, verifying that the machinery for IL-6 production operates in these cells.
Myeloid cells are key drivers of thrombo-inflammation [25] and of inflammation-resolution in the heart [26]. Neutrophil-secreted myeloperoxidase (MPO) is a critical mediator of hypochloric acid formation and the subsequent generation of AOPP like oxAlb [27-29]. To assess if neutrophil-derived AOPP may provoke IL-6 production in cardiomyocytes, we first incubated freshly isolated human neutrophils in autologous PRP in absence and presence of thrombin. We found increased externalization of active MPO together with higher AOPP formation in conditioned medium from thrombin-stimulated compared to control neutrophils. To the best of our knowledge this is the first demonstration of this phenomenon. The neutrophil-conditioned PRP was then added to HL-1 recipient cells for 24h. HL-1 cells exposed to supernatant from thrombin-stimulated neutrophils displayed higher levels of IL-6 mRNA and greater IL-6 secretion than cells exposed to PRP conditioned with control neutrophils. To confirm that AOPP contained in neutrophil-conditioned medium was the culprit mediator of IL-6 production, we employed the specific CD36 inhibitor SSO. In the presence of SSO, the stimulatory effect of PRP from thrombin-stimulated neutrophils on IL-6 expression and release was reduced, significantly in the case of IL-6 secretion.
Thrombo-inflammation and cardiac injury and disease are hallmarks of visceral obesity [30-32]. We have previously reported increased abundance of PAR4 thrombin receptors in adipose tissue and myocardium of mice with HFD-induced obesity and could link PAR4 causally with local tissue inflammation [3, 18]. LV biopsies archived from these mice were now assessed specifically for the markers we observed in the in vitro studies. LV from HFD-fed mice exhibited greater MPO activity and AOPP content than chow-fed mouse LV, together with a higher abundance of the scavenger receptor CD36. This suggests that the machinery for local AOPP generation and cellular uptake is present in obese myocardium. In keeping with the association between AOPP and IL-6 production in our in vitro system, we could also detect greater IL-6 expression in HFD-fed mouse myocardium compared to chow controls.
Myocardial-level thrombo-inflammation likely involves the activation of neutrophils by thrombin, leading to the oxidative modification of circulating and tissue-bound proteins such as albumin. The oxidized products may be taken up by cardiomyocytes via CD36 scavenger receptors to trigger inflammatory signaling, including the transcription and release of IL-6. A proposed scheme is depicted in Fig. 4. In this way the thrombin/PAR system may indirectly modulate local inflammation-resolution pathways, particularly in the obese heart, and could provide a platform for therapeutic intervention.
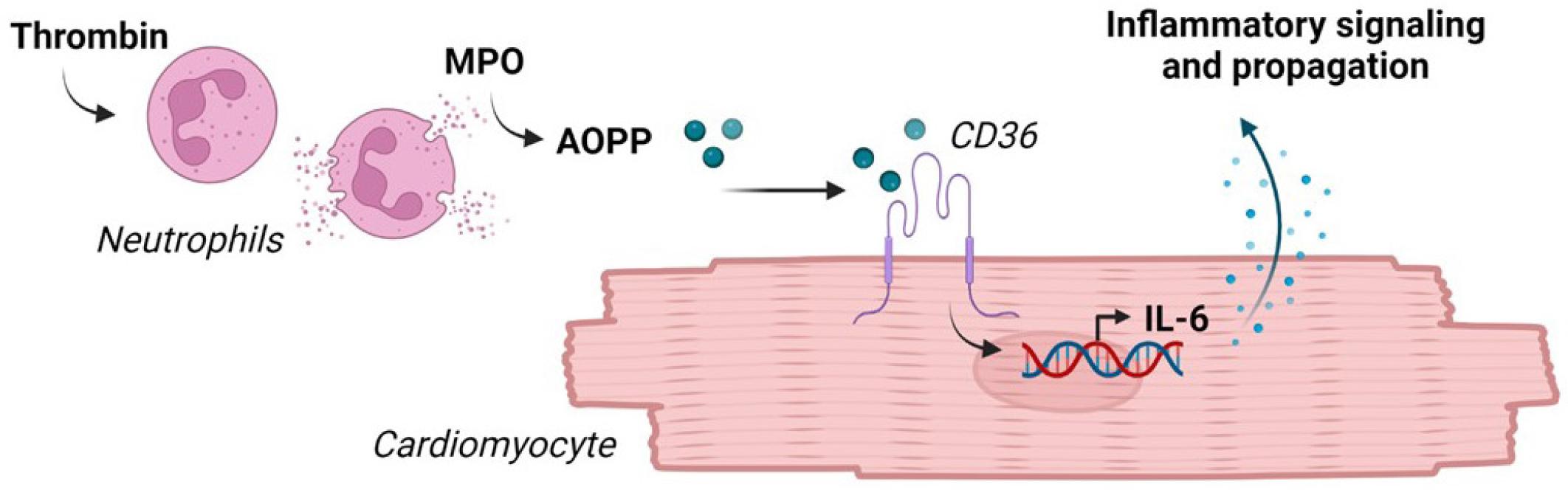
Schematic depiction of working hypothesis: Thrombin stimulates MPO release and AOPP formation by neutrophils. AOPP uptake via CD36 scavenger receptors provokes IL-6 transcription and secretion by cardiomyocytes, to modulate local inflammatory events.