
Cryptosporidium is a genus of apicomplexan protozoa infecting a broad spectrum of vertebrate hosts, including fish, amphibians, reptiles, birds and mammals (8). Cryptosporidiosis may progress to a chronic, life-threatening human disease characterised by severe diarrhoea, particularly in children and immunocompromised individuals (19, 20). Approximately 40 species of Cryptosporidium have been identified, with C. hominis being the predominant cause of human infections; however, zoonotic transmission from animal-associated species such as C. canis, C. felis, C. suis, C. erinacei, C. parvum, C. meleagridis and C. cuniculus is also well documented (8). Avian cryptosporidiosis was first documented in 1929, when Tyzzer identified oocysts in the caecal epithelium of chickens (46). In contrast to mammals, where the parasites are largely confined to the intestinal tract, avian species frequently harbour infections in the cloacal bursa and respiratory system. In chickens and turkeys, Cryptosporidium has been detected in the sinuses, trachea, bronchi, cloaca and bursa of Fabricius (11). The most frequently detected species in poultry are C. baileyi, C. meleagridis, C. galli and C. avium (8). Among wild birds, six species (C. andersoni, C. parvum, C. meleagridis, C. avium, C. baileyi and C. galli) and five genotypes (goose I and II and avian I, III and VI) have been documented (48). Cryptosporidium proventriculi (formerly avian genotype III) has been reported in Psittaciformes, Passeriformes, toucans and migratory ducks (14, 31, 38). The zoonotic species C. meleagridis, a major cause of human cryptosporidiosis, has additionally been identified in wild Anatidae (22) and Columbidae (14, 24). Oocyst shedding is influenced by species, host age and immune status, but typically is in low abundance, which along with the small size of oocysts complicates detection. Conventional diagnosis relies on concentration methods (sucrose flotation or sedimentation) followed by acid-fast staining (Ziehl–Neelsen or Kinyoun) (18). Immunoassays, such as ELISA and rapid immunochromatographic (IC) tests, allow high throughput or rapid screening (4). Molecular approaches, including nested, multiplex and real time PCR as well as PCR and restriction fragment length polymorphism, represent the current gold standard for species identification and genotyping (8, 11, 18, 39, 51). This study aimed to detect Cryptosporidium in different species of asymptomatic and clinically affected pet birds by applying two molecular methods in nested PCR form and a rapid immunochromatographic assay.
Between March 2024 and April 2025, faecal samples from 63 pet birds comprising 52 parrots, 8 pigeons and 3 canaries were tested in a private veterinary laboratory in Warsaw. There were 22 female birds and 41 males. All samples originated from a single veterinary clinic and were collected during routine visits, which were supplemented by clinical examination and veterinary interview. Each faecal sample was collected individually from each bird and no samples were pooled. The birds were placed in a transport container lined with clean paper towels, and fresh faeces were collected for analysis. All birds included in the study were kept in cages or indoor aviaries without access to the external environment or direct contact with free-living wild birds. None of the pigeons were feral birds.
To facilitate statistical analyses and evaluate age-related trends, the 63 birds included in the study were divided into four age categories. The categorisation was designed to achieve a balance in group size while preserving biologically relevant developmental stages. The following age groups were established: juveniles of ≤1 year (n = 11), early adults of >1 to 2 years (n = 14), mature adults of >2 to 5 years (n = 22) and senior birds >5 years (n = 16). This stratification allowed meaningful statistical comparisons while accounting for developmental and physiological differences associated with age, and differed little from that adopted in previous avian epidemiological and microbiome studies (45).
Cloacal, crop, skin and feather-calamus swabs were cultured on blood agar and MacConkey agar (GRASO Biotech, Starogard Gdański, Poland). Plates were incubated aerobically at 37°C for 24–48 h. Bacterial isolates were identified based on colony morphology, Gram staining, biochemical characteristics, and, when necessary, by MALDI-TOF (Microflex LT MALDI-MS Biotyper System; Bruker Daltonics, Bremen, Germany).
Parasitological screening was conducted using direct smear and zinc sulphate flotation techniques (7). For the direct smear, ~100 mg of faeces was mixed with 0.9% saline, and a drop was examined microscopically at 100× and 400× magnification. For zinc sulphate flotation, samples were mixed with saturated ZnSO4 solution at a specific gravity of 1.31 g/cm3, centrifuged for 5 min at 3,000 rpm and examined after 25 min using a coverslip placed on the meniscus. Cryptosporidium spp. was detected with a commercial rapid immunoassay (Simple Crypto; Operon, Zaragoza, Spain) per the manufacturer’s instructions. The test demonstrated an estimated sensitivity and specificity of 99.9% relative to microscopic examination. Diagnostic performance was further evaluated against a one-tube nested real-time PCR assay, used as the reference method.
For molecular analysis, 50–100 mg of fresh faeces was collected and stored frozen. Isolation of DNA was performed using the commercial Genomic Mini AX Stool kit (A&A Biotechnology, Gdańsk, Poland) according to the manufacturer’s protocol. Extracted DNA was resuspended in 200 μL of Tris elution buffer and frozen at –20°C for further tests.
A nested PCR was performed according to a previously described protocol (51), using two primer pairs. The first round employed 5′-TTCTAGAGCTAATACATGCG-3′ and 5′-CCCTAATCCTTCGAAACAGGA-3′ primers (expected product: 1325 bp), and the second round used 5′-GGAAGGGTTGTATTTATTAGATAAAG-3′ and 5′-AAGGAGTAAGGAACAACCTCCA-3′ (expected product: ~840 bp) primers. Each 25 μL reaction included StartWarm HS-PCR Mix (A&A Biotechnology), 1.5 μL of each primer (5 μM), 2 μL of template (genomic DNA or first-round product) and nuclease-free water. The PCR conditions were: initial denaturation at 95°C for 3 min; 35 cycles at 95°C for 30 s, 55°C for 45 s and 72°C for 60 s; and a final extension at 72°C for 10 min. The reactions were run on a MultiGene OptiMax thermal cycler (Labnet, Edison, NJ, USA). Genomic DNA of Cryptosporidium parvum and ultrapure water served as positive and negative controls, respectively. Nested PCR products were analysed by 2% agarose gel electrophoresis. Amplified products were purified and Sanger sequenced bidirectionally by a commercial provider (Oligo IBB, Warsaw, Poland). Chromatograms were edited using Chromas 2.6.6 (Technelysium, South Brisbane, QLD, Australia) and MEGA 7 (21). Consensus sequences assembled from forward and reverse reads were compared to NCBI GenBank references using BLAST.
The assay used two primer pairs: the first, described by Johnson et al. (17), amplified a ~428 bp fragment of the Cryptosporidium 18S ribosomal RNA (rRNA) gene and the second, by Santana et al. (39), validated a one-tube nested real-time PCR protocol for avian faecal samples. The reaction was performed in a 20 μL total volume, per the original protocol (39), using 10 μL of EvaGreen RT PCR Mix (A&A Biotechnology), 0.5 μL of each of the NRT18SF (5′-GTTGTTGCAGTTAAAAAGCTCGTAGTTGGATT-3′) and NRT18SR (5′-ACTTTGATTTCTCATAAGGTGCTGAAGG-AGT-3′) primers at 2.5 μM and CPB-DIAGF (5′-AAGCTCGTAGTTGGATTTCTG-3′) and CPB-DIAGR (5′-TAAGGTGCTGAA-GGAGTAAGG-3′) primers at 5 μM, 2 μL of DNA and 6 μL of ultrapure water. Reactions were run on a Gentier 48E Real-Time PCR System (TianLong Biotechnology, Xi’An, China), with fluorescence monitored during the nested PCR stage. The thermal cycling comprised a first step at 95°C for 5 min and 20 cycles of reaction at 95°C for 15 s and 70°C for 30 s, and a second step of 35 cycles of reaction of 95°C for 15 s, 62°C for 30 s and 72°C for 20 s. High-resolution melting curve analysis followed, working with data from 60°C to 95°C at 0.5°C increments and 5-s steps. The decrease in fluorescence intensity with increasing temperature reflected the denaturation of PCR amplicons, and the distinct melting profiles differentiated species based on their melting temperatures. This method enabled discrimination of closely related Cryptosporidium species based on differences in DNA sequence composition. The positive control was the DNA of C. parvum and the negative control was ultrapure water.
Descriptive statistics were used to summarise the prevalence of Cryptosporidium infection among the examined birds. To assess the association between sex (male vs female), age and infection status (positive vs negative; Cryptosporidium species vs host species), as well as the relationship between infection, clinical signs and co-infections, two statistical tests were applied. A chi-squared (χ2) test of independence was used for a 2 × 2 contingency table, and Fisher’s exact test was performed to confirm the reliability of the results owing to the relatively small sample size. A P-value < 0.05 was considered statistically significant. All statistical analyses were performed using Statistica 13.1 software (Dell, Round Rock, TX, USA).
The validation of the rapid immunochromatographic test was conducted using the one-tube nested real-time PCR as the reference method. This allowed the classification of results as true positives (TP), false positives (FP), true negatives TN) and false negatives (FN). Based on these values, the sensitivity and specificity of the rapid test were calculated using the following formulae: sensitivity = TP / (TP + FN) and specificity = TN / (TN + FP).
Microscopic examination revealed the presence of protozoa in two pigeons: Eimeria spp. oocysts in one individual and Trichomonas spp. trophozoites in the other. No helminth eggs were detected in any of the examined samples. Using the one-tube nested real-time PCR assay, Cryptosporidium spp. DNA was detected in 33.3% of the faecal samples (21/63), which were those from 16 parrots and 5 pigeons. The nested PCR method identified Cryptosporidium spp. in 25.4% of samples (16/63), which were those from 13 parrots and 3 pigeons. In comparison, the antigen detection test yielded positive results in 30.2% of cases (19/63). None of the faecal samples from canaries tested positive in the applied methods. Additionally, co-infections were observed in two pigeons: one with Eimeria spp. and Cryptosporidium spp., and the other with Trichomonas spp. and Cryptosporidium spp. Sequencing of the products obtained from both the nested PCR and the one-tube nested real-time PCR enabled the identification of the parasite species. Cryptosporidium proventriculi was detected in 15 samples from parrots, while C. meleagridis was identified in 5 samples from pigeons and 1 from a parrot (Figs 1–3). The complete dataset is available in Supplementary Table S1.
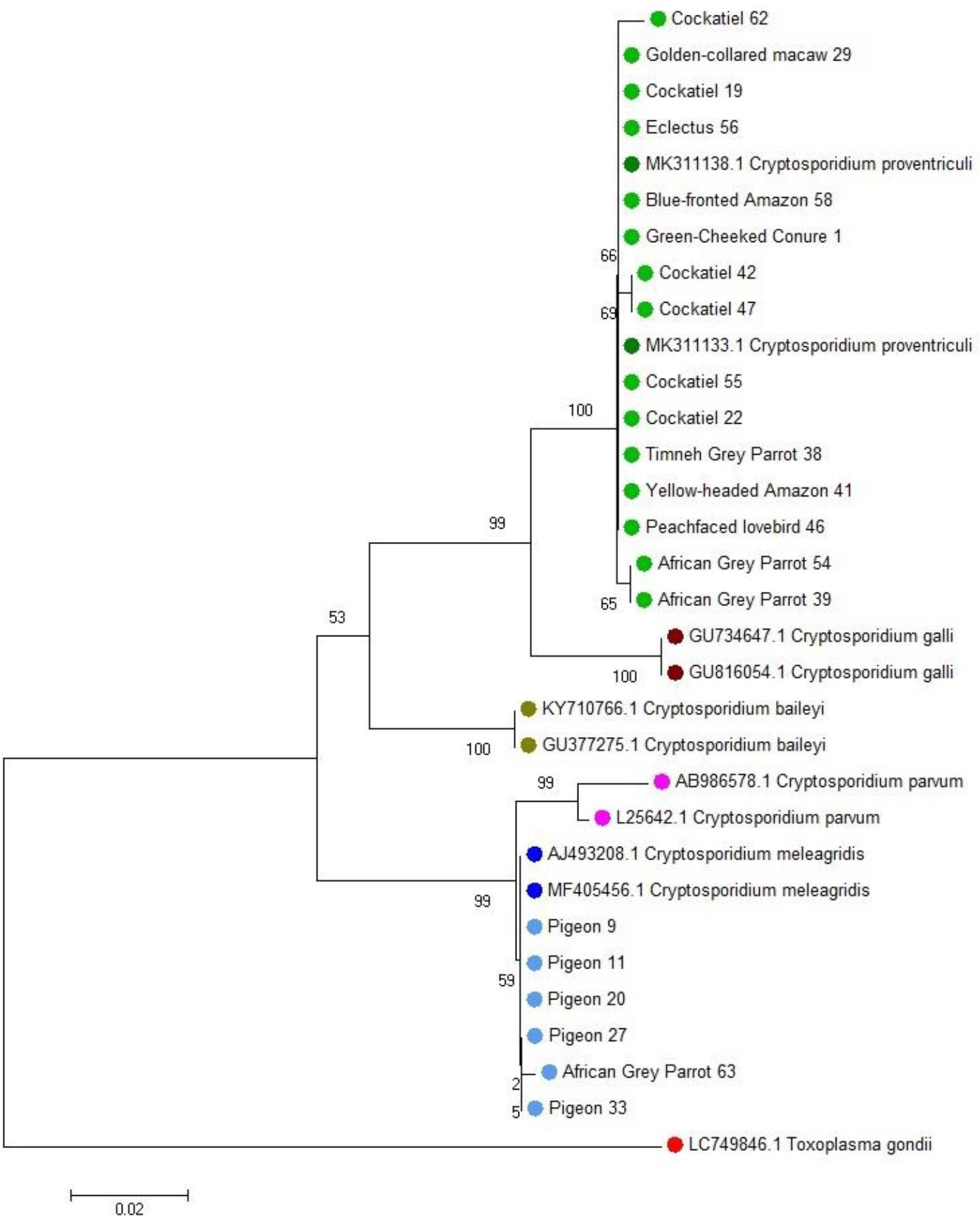
Phylogenetic relationships of 32 nucleotide sequences of Cryptosporidium species isolated from pet birds. The analysis was performed using the neighbour-joining method based on nucleotide sequences of the small-subunit ribosomal RNA gene. The evolutionary distances were computed using the Kimura two-parameter method and are the numbers of base substitutions per site. Two-letter and six-digit codes are GenBank accession numbers. Toxoplasma gondii was used as the outgroup
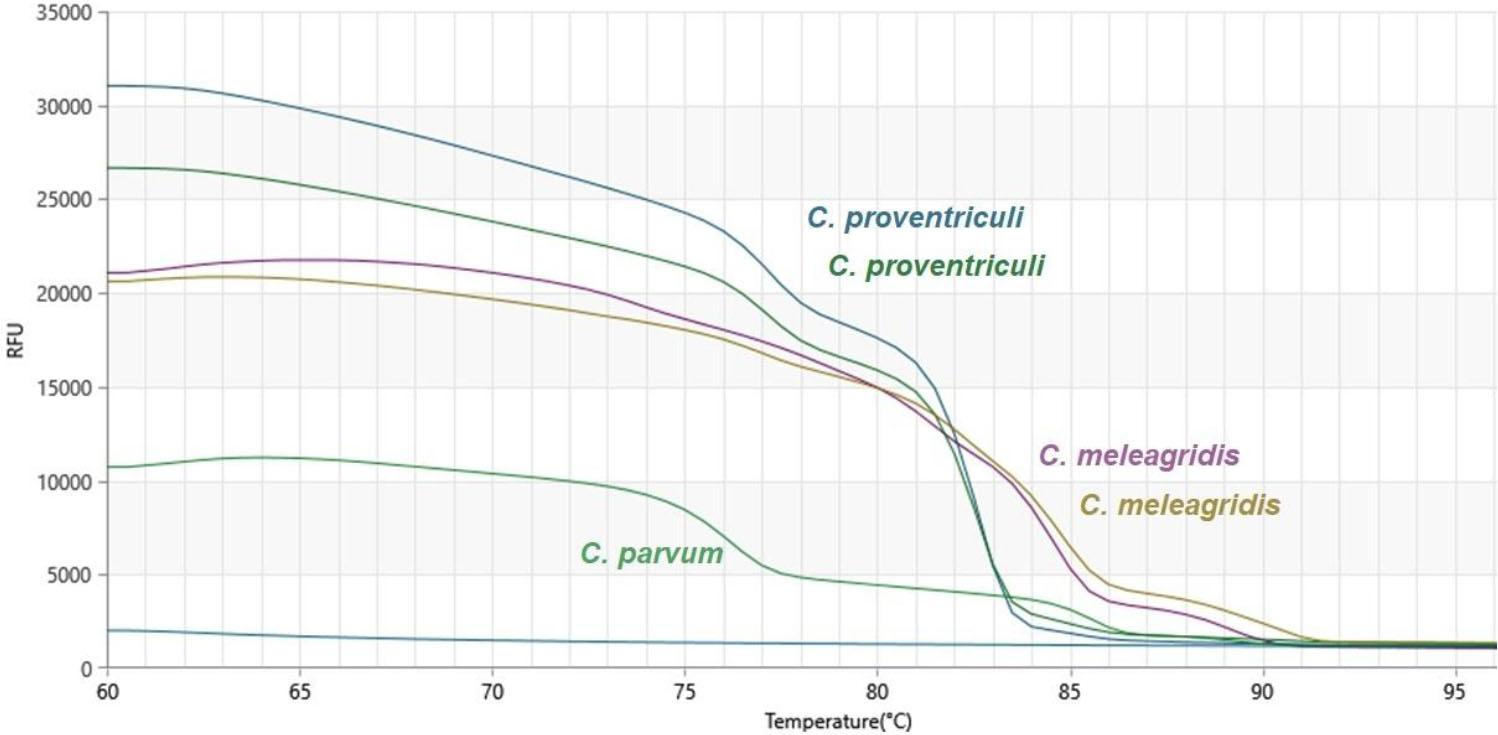
Melting curves obtained during qPCR-high-resolution melting analysis of the products of amplification of isolates of Cryptosporidium spp. from the faeces of pet birds. The paired curves for C. meleagridis and C. proventriculi indicate technical replicates or distinct isolates. RFU – relative fluorescence units
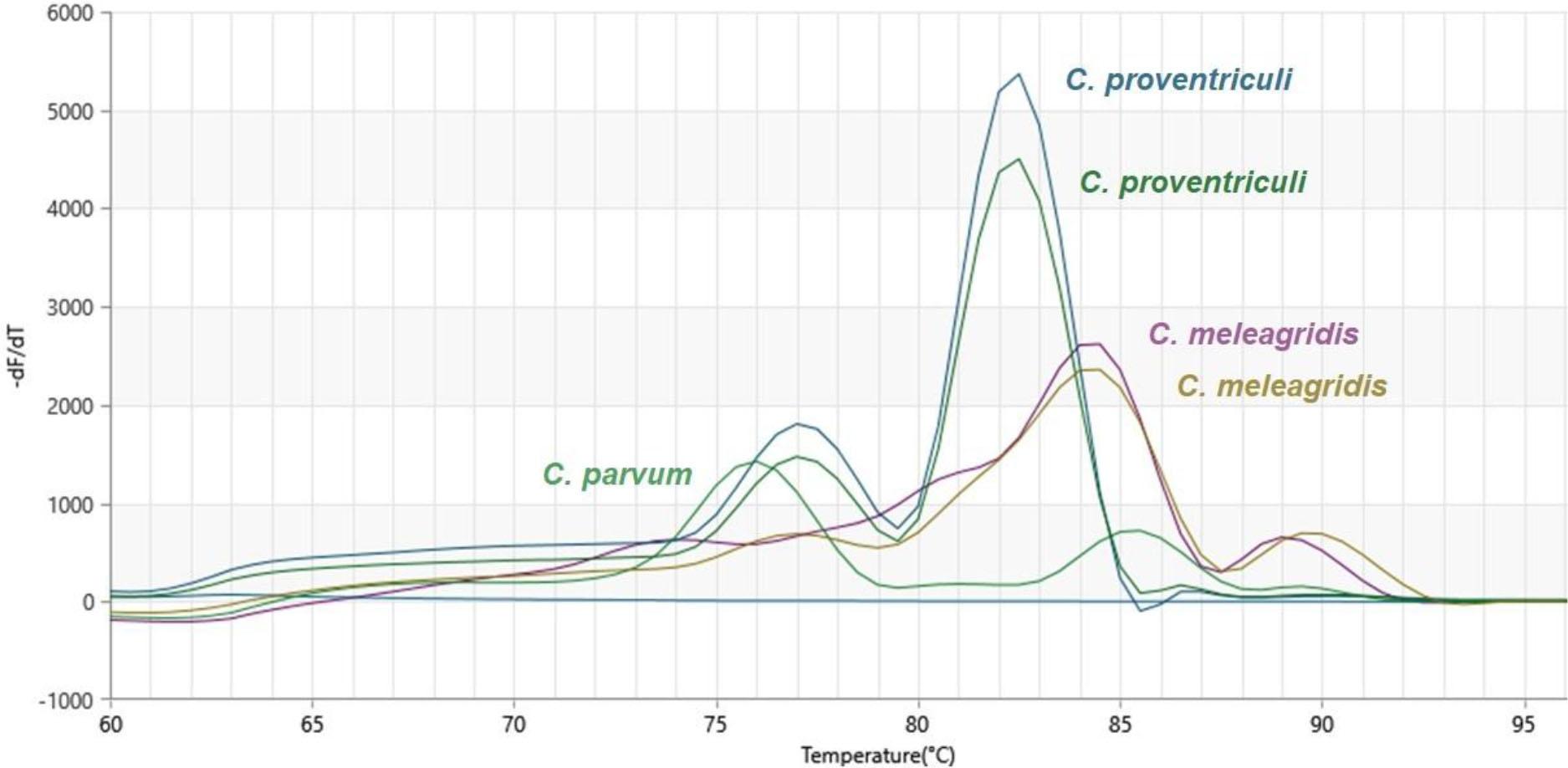
Derivative melting curve analysis of the products of amplification of isolates of Cryptosporidium spp from the faeces of pet birds. The paired curves for C. meleagridis and C. proventriculi indicate technical replicates or distinct isolates. –dF/dT – negative first derivative of fluorescence with respect to temperature
Cryptosporidium spp. was detected in 21 of the 63 birds (33.3%), all of which exhibited various clinical symptoms. Among the remaining 42 birds, 28 showed clinical signs despite testing negative for Cryptosporidium spp., while 14 birds had no symptoms and tested negative. The chi-squared test revealed a statistically significant association (χ2 = 7.17, P-value = 0.0074). Moreover, Fisher’s exact test confirmed an important relationship (P-value = 0.0026), indicating that Cryptosporidium-infected birds were significantly more likely to present with clinical signs than uninfected individuals. These findings support the hypothesis that Cryptosporidium infection was significantly associated with clinical symptoms in the examined birds (P-value < 0.01). Fisher’s exact test also revealed a statistically significant association between Cryptosporidium infection and concurrent bacterial or other protozoan infections (P-value = 0.0002, odds ratio = 9.17), suggesting that co-infections were substantially more likely in Cryptosporidium-positive birds. Although clinical signs were observed in most birds with bacterial infections (either alone or in co-infection), chi-squared analysis did not reveal a statistically significant association between infection type and clinical symptom occurrence (χ2 = 3.12, P-value = 0.21).
Diarrhoea, poor skin condition, vomiting and/or regurgitation and sneezing and/or upper respiratory tract disorders were selected from the most frequently observed clinical signs for statistical analysis in relation to Cryptosporidium infection. In the cohort of 63 pet birds examined, diarrhoea was observed in 6 individuals, of which 4 tested positive for Cryptosporidium. Among the 57 birds without diarrhoea, 15 were positive for the parasite. Statistical analysis using the chi-squared test revealed a significant association between diarrhoea and Cryptosporidium infection (χ2 = 4.20, degrees of freedom (df) = 1, P-value < 0.05), suggesting that the presence of Cryptosporidium is significantly associated with the occurrence of diarrhoea in the studied population. This finding supports the hypothesis that Cryptosporidium infection may have contributed to gastrointestinal disturbances in these birds. Skin in poor condition was identified in 5 birds, all of which tested positive for Cryptosporidium, whereas 16 out of 58 birds with healthy skin were infected. The association between poor skin condition and Cryptosporidium infection was highly significant (χ2 = 10.84, df = 1, P-value < 0.01), indicating a potential link between the parasite and dermatological manifestations. This may reflect systemic effects of the infection or compromised health status in affected individuals. Conversely, only 2 of the 8 birds noted with vomiting and/or regurgitation tested positive for Cryptosporidium, while 19 of the 55 birds without these clinical signs were infected. The chi-squared test did not demonstrate a statistically significant relationship between vomiting/regurgitation and Cryptosporidium infection (χ2 = 0.28, df = 1, P-value > 0.05), suggesting that these symptoms were unlikely to be directly associated with the parasite in this cohort. Similarly, birds exhibiting sneezing and/or upper respiratory tract disorders showed an identical non-significant result (χ2 = 0.28, df = 1, P-value > 0.05), indicating no significant association between respiratory symptoms and Cryptosporidium infection. These findings imply that while Cryptosporidium infection may have been linked to certain clinical signs such as diarrhoea and dermatological changes, other symptoms observed in the examined birds were likely attributable to different aetiologies or multifactorial causes.
Infection with Cryptosporidium spp. was detected in 9 of the 22 female birds (40.9%) and 12 of the 41 males (29.3%). Statistical analysis was conducted to assess the association between sex and infection status. A chi-squared test of independence revealed no statistically significant relationship between sex and Cryptosporidium infection (χ2 = 0.87, df = 1, P-value = 0.35). Fisher’s exact test also confirmed the lack of significant association (P-value = 0.41). These results indicate that, within this sample, sex was not an important factor influencing the prevalence of Cryptosporidium infection.
The prevalence of Cryptosporidium infection in the youngest group was 45.5%, in the early adult group was 28.6%, in the mature adult group was 31.8% and in the senior group was 31.3%. A chi-squared test of independence showed no statistically significant association between age group and Cryptosporidium infection (χ2 = 0.92, df = 3, P-value = 0.82), indicating that age was not an important factor influencing the likelihood of infection in this population.
In this study, C. proventriculi was detected exclusively in parrots (Psittacidae family, including Psittacus, Nymphicus and other genera), with 15 out of 52 parrots (30.8%) but none of the 8 pigeons (Columbidae) testing positive. A Fisher’s exact test was performed to evaluate whether the occurrence of C. proventriculi differed significantly between the two families. The test showed a statistically significant association (P-value = 0.048), indicating that C. proventriculi was significantly more frequent in parrots than pigeons. Within the Psittacidae family itself, C. proventriculi was detected in 6 out of 20 cockatiels (Nymphicus hollandicus; 30.0%) and 9 out of 43 individuals of other genera (20.9%), although Fisher’s exact test did not reveal a statistically significant difference between these groups (P-value = 0.327). These results suggest that although C. proventriculi was most frequently identified in cockatiels, the difference in prevalence was not statistically significant within the studied population (Table 2).
Results of the PCR test for the presence of two Cryptosporidium spp. in the studied bird group
| Species | n | C. proventriculi | C. meleagridis | ||
|---|---|---|---|---|---|
| n (+) | % | n (+) | % | ||
| African grey parrot (Psittacus erithacus) | 3 | 2 | 66.7 | 1 | 33.3 |
| Blue-and-yellow macaw (Ara ararauna) | 2 | 0 | 0.0 | 0 | 0.0 |
| Blue-fronted amazon (Amazona aestiva) | 2 | 1 | 50.0 | 0 | 0.0 |
| Blue-headed pionus (Pionus menstruus) | 1 | 0 | 0.0 | 0 | 0.0 |
| Budgerigar (Melopsittacus undulatus) | 4 | 0 | 0.0 | 0 | 0.0 |
| Canary (Serinus canaria) | 3 | 0 | 0.0 | 0 | 0.0 |
| Cockatiel (Nymphicus hollandicus) | 20 | 6 | 30 | 0 | 0.0 |
| Eastern rosella (Platycercus eximius) | 2 | 0 | 0.0 | 0 | 0.0 |
| Moluccan eclectus (Eclectus roratus) | 1 | 1 | 100 | 0 | 0.0 |
| Golden-collared macaw (Primolius auricollis) | 1 | 1 | 100 | 0 | 0.0 |
| Green-cheeked conure (Pyrrhura molinae) | 3 | 1 | 33.3 | 0 | 0.0 |
| Peach-faced lovebird (Agapornis roseicollis) | 4 | 1 | 25 | 0 | 0.0 |
| Pigeon (Columba livia) | 8 | 0 | 0.0 | 5 | 62.5 |
| Plum-headed parakeet (Psittacula cyanocephala) | 2 | 0 | 0.0 | 0 | 0.0 |
| Rainbow Lorikeet (Trichoglossus moluccanus) | 2 | 0 | 0.0 | 0 | 0.0 |
| Ring-necked parakeet (Psittacula krameri) | 1 | 0 | 0.0 | 0 | 0.0 |
| Senegal parrot (Poicephalus senegalus) | 1 | 0 | 0.0 | 0 | 0.0 |
| Sun conure (Aratinga solstitialis) | 1 | 0 | 0.0 | 0 | 0.0 |
| Timneh grey parrot (Psittacus erithacus timneh) | 1 | 1 | 100 | 0 | 0.0 |
| Yellow-headed amazon (Amazona oratrix) | 1 | 1 | 100 | 0 | 0.0 |
| Total | 63 | 15 | 23.8 | 6 | 9.52 |
The nested real-time PCR identified 21 positive and 42 negative individuals. The results were divided into groups (Table 3), and the assay’s sensitivity and specificity were 66.7% and 88.1%, respectively.
Diagnostic performance of the immunochromatographic assay evaluated against one-tube nested real-time PCR for detection of Cryptosporidium spp. in avian faeces
| Positive (ref. method n = 21) | Negative (ref. method n = 42) | |
|---|---|---|
| IC rapid assay positive | 14 | 5 |
| IC rapid assay negative | 7 | 37 |
Infections caused by Cryptosporidium spp. have been reported in numerous bird species in the Passeriformes, Psittaciformes and Columbiformes orders worldwide (31, 32, 33, 34, 38). Cryptosporidium proventriculi, previously referred to as avian genotype III, appears to be the most prevalent species infecting parrots in both the New and Old Worlds (9, 14, 16, 23, 25, 26, 27, 29, 30, 33, 34, 35, 37, 38). In our study, C. proventriculi was detected exclusively in faecal samples from parrots. This parasite species is a known cause of proventricular inflammation and chronic vomiting in some psittacine birds. Makino et al. (25) confirmed C. proventriculi infection in 13 of 37 peach-faced lovebirds using PCR, noting chronic vomiting, weight loss and radiographic signs such as isthmus enlargement, narrowed lumens and thickened proventricular walls. In our study, infected parrots presented with vomiting, regurgitation, obesity, diarrhoea, beak overgrowth and feather degradation. Dermatitis and feather plucking were noted, with one bird having a wing wound. Laboratory tests indicated bacterial infections of the skin, intestines and crop. In some cases, ultrasound and blood analysis confirmed hepatic steatosis, cirrhosis and hepatitis. Ravich et al. (35) performed a histopathologic analysis of 34 psittacine birds – 12 cockatiels, 18 lovebirds (2 Agapornis roseicollis and 16 of unknown species) and four parrotlets (Forpus sp.). Although histopathological examinations of birds that had died suddenly revealed the presence of Cryptosporidium and associated mucosal hyperplasia of the proventriculus, other lesions were also observed, suggesting that the Cryptosporidium infection was secondary and should not have been considered the primary cause of death. However, the researchers emphasised that in 11 birds, the lesions were exclusively associated with gastrointestinal cryptosporidiosis. Cryptosporidium galli and C. proventriculi are considered two of the most pathogenic Cryptosporidium species in birds and cause clinical signs related to the gastrointestinal tract such as vomiting, diarrhoea and weight loss (11, 28, 33, 37, 41). In our study, a statistically significant association between diarrhoea and C. proventriculi infection was observed, consistently with the findings of Pangeossi et al. (33), who reported a similar association in a group of 100 cockatiels (P-value = 0.017). The same authors also identified a significant relationship between Cryptosporidium infection and the birds’ origin, noting that owned birds had a higher prevalence of infection than those from pet shops. In our study, all examined birds were privately owned.
Cryptosporidium galli has been identified in poultry and birds of the Passeriformes order (29, 30, 39). Studies conducted by da Silva et al. (41) indicate that C. galli is commonly found in great-billed seed finches (Oryzoborus maximiliani), lesser seed finches (Oryzoborus angolensis), ultramarine grosbeaks (Cyanocompsa brissonii) and rusty-collared seedeaters (Sporophila collaris). However, its virulence appears to be low. Pathogenic effects were observed only in one aviary, where bird mortality was associated with coinfection with E. coli. In our study, no sample was C. galli positive. Although no cases of mortality were observed among our patients, the vast majority of birds infected with Cryptosporidium presented with concurrent bacterial infections of the skin, intestines or crop, which may be considered co-infections with associated clinical manifestations. Clinical signs were frequently observed in the study birds with bacterial infections, whether present alone or in combination with other pathogens; however, statistical analysis did not show a significant association between the type of infection and the occurrence of clinical symptoms.
Cryptosporidium meleagridis was first described in young turkeys (43), but has also been found in chickens, ducks, pet birds and wild birds (2, 5, 12, 13, 23, 26, 34, 48). In our study, it was detected in five pigeons and one African grey parrot. Reports on Cryptosporidium genotypes in pigeons are limited (24, 32, 41) and correspondingly, those focusing on pigeons in Poland are sparse (13). As synanthropic birds common in urban and rural areas, pigeons may play a key role in pathogen transmission. In Chile, Briceño et al. (5) reported C. meleagridis in invasive monk parakeets from Santiago, and their finding points to the need for ongoing surveillance in widely distributed synanthropic species. Cryptosporidium meleagridis is among the three species in the genus most frequently linked to human cryptosporidiosis (8, 42). In our study, it was found in an African grey parrot with chronic respiratory infection caused by Mycobacterium sp. Similarly, Huh et al. (15) reported chronic diarrhoea in an AIDS patient coinfected with Mycobacterium ulcerans and Cryptosporidium spp. In Poland, human cryptosporidiosis caused by C. meleagridis was reported in four patients: two immunocompromised children (3, 50), an adult HIV-positive woman suffering from chronic diarrhoea (49) and an immunocompetent patient with colon adenocarcinoma (19). All these cases highlight the importance of comprehensive diagnostic workups, particularly in immunocompromised patients suffering from chronic respiratory infections or persistent diarrhoea.
Detection of Cryptosporidium spp. by PCR offers greater sensitivity than traditional microscopy or immunoassays, enables high-throughput analysis and identifies the parasite precisely to species level (6, 27, 39, 47, 51). Key genetic targets for species differentiation include small-subunit rRNA, Cryptosporidium oocyst-wall protein, 70-kDa heatshock protein, thrombospondin-related anonymous protein C2 and the actin gene (14, 30, 36, 51). The most widely used method for diagnosing cryptosporidiosis and conducting phylogenetic analysis is amplification of the 18S rRNA gene fragment (13, 26, 29, 37, 51). Our study utilised two PCR-based approaches which amplified this fragment: nested PCR (51) and one-tube nested real-time PCR (17, 39). However, for initial screening for Cryptosporidium spp., we applied a rapid immunochromatographic test targeting protozoan antigens. Antigen presence was detected in 30.2% (19/63) of faecal samples. Using molecular methods, Cryptosporidium spp. DNA was identified in 25.4% (16/63) of samples with nested PCR and in 33.3% (21/63) in the one-tube nested real-time PCR. As all faecal samples were tested using the same diagnostic methods, we validated the rapid test against the one-tube nested real-time PCR, the chain reaction being considered the reference based on our findings and those of Santana et al. (39). The rapid immunochromatographic test showed a sensitivity of 66.7% and specificity of 88.1%, indicating moderate sensitivity and high specificity for detecting Cryptosporidium antigen in avian faeces. Although the test was designed for human samples, similar products from the same manufacturer have been used in dogs and cats and have achieved sensitivity of 46.1% and specificity of 99.0% (4). Abdou et al. (1) also used a rapid immunochromatographic test in cattle and detected Cryptosporidium in 23% (92/400) of samples, establishing the test’s sensitivity at 74.07%. These tests have nevertheless been associated with high false-positive rates. In our study, five samples tested positive for antigen in the rapid IC assay but negative in both PCR tests. Conversely, seven PCR-positive samples lacked detectable antigen in rapid IC. These discrepancies and the modest test performance in birds may result from interference by substances in avian faeces, such as uric acid, which is absent from human stool. The current literature offers limited data on rapid immunochromatographic test performance in birds. One study assessing occult blood tests in cockatiels suggested that false positives could stem from diet, peroxidase activity or minor gastrointestinal bleeding (10). These findings indicate that there is a need for reliable diagnostic tools for Cryptosporidium spp. detection in avian veterinary practice, particularly since no effective therapy is available for avian cryptosporidiosis. Early detection enables timely isolation and biosecurity measures to prevent spread. Therapy with paromomycin has demonstrated partial efficacy in chickens, reducing oocyst shedding, although it may predispose birds to secondary fungal infections (44). Consequently, control strategies rely primarily on prophylaxis, afforded jointly through strict biosecurity and hygiene to negate the advantage provided by the marked environmental resistance of oocysts, and through immune support to at-risk birds (40).
Cryptosporidium spp. infections are widespread in pet and wild birds, with clinical involvement often extending beyond the gastrointestinal tract and frequently complicated by bacterial co-infections. Among psittacines, C. proventriculi was the dominant species, while C. meleagridis, a zoonotic pathogen of major public health concern, was also identified in pigeons and parrots. Given the absence of effective therapies and the risk of secondary fungal infections associated with paromomycin, preventive measures like biosecurity, hygiene, nutritional balance and immune support remain essential. Importantly, zoonotic species such as C. meleagridis warrant molecular confirmation in birds, particularly those owned by immunocompromised individuals. Detection of Cryptosporidium in avian patients should prompt comprehensive clinical evaluation, as infection may signal underlying systemic disease.