
The soybean cyst nematode (SCN), Heterodera glycines Ichinohe, is the most economically significant pathogen in U.S. soybean production (Bandara et al., 2020; Bradley et al., 2021), with the latest estimated yield reductions exceeding $1.5 billion per year (Andres and Grabau, 2025). The nematodes can be found at damaging population levels, causing 30% yield reductions without exhibiting obvious aboveground symptoms (Wang et al., 2003; Allen et al., 2017). Growers are increasingly relying on a few resistant sources, mainly PI 88788 and, more recently, Peking (Tylka and Marett, 2025). However, the overuse of cultivars with PI 88788 as the resistance source has shown diminishing efficacy in protecting against H. glycines (McCarville et al., 2017; Tylka, 2021). Disease management with soil-applied nematicides is not cost-effective for the millions of acres of soybeans planted across the United States. Nematicide seed treatments offer an alternative to soil-applied nematicides (Tylka and Marett, 2025). Such treatments often contain a potpourri of synthetic pesticides, fungicides, insecticides, and a nematicide. The latter is sometimes replaced with microbiologically derived options such as abamectin or live bacteria, which may provide some protection against H. glycines yield reduction. The efficacy of microbial seed protectants against SCNs has been variable in field trials (Bissonnette et al., 2018, 2024; do Nascimento et al., 2022; Jensen et al., 2018; Kessler and Koehler, 2023; Tylka and Marett, 2025).
Mitigating cyst nematode-caused diseases and yield reductions through microbial parasitism was first considered by Julius Kühn, who observed parasitized H. schachtii females during his research on beet weariness (Rübenmüdigkeit) (Kühn, 1877). Numerous publications have subsequently focused on potential biocontrol organisms for cyst nematodes (Stirling, 2014), particularly concerning soybeans (Kim and Riggs, 1991, 1995; Chen and Dickson, 1996; Meyer et al., 1997; Timper et al., 1999; Chen and Liu, 2005; Silva et al., 2022; Zhou et al., 2024). For the past three decades, our group has investigated biologically suppressive soils in which virulent endoparasitic nematodes do not thrive despite conducive environmental conditions and susceptible host crops. The ascomycete Hyalorbilia oviparasitica (Baral et al., 2018, 2020) was previously known only by its anamorphic stage Dactylella oviparasitica (Stirling and Mankau, 1978). This fungus and closely related Hyalorbilia species have been found to parasitize economically significant endoparasitic nematodes, such as the Southern root-knot nematode (Meloidogyne incognita) (Stirling and Mankau, 1978), the guava root-knot nematode (M. enterolobii) (Liang et al., 2020), sugarbeet cyst nematode (Heterodera schachtii) (Westphal and Becker, 2001; Olatinwo et al., 2006), and the SCN (H. glycines) (Kim and Riggs, 1991; Yang et al., 2012). Fungi belonging to the H. oviparasitica clade were demonstrated to be the primary cause of the long-term suppression of the sugarbeet cyst nematode at field 9E of the University of California, Riverside, Agricultural Operations (Westphal and Becker, 2001; Olatinwo et al., 2006). An isolated strain, H. sp. aff. multiguttulata DoUCR50, was shown to parasitize sedentary late-stage juveniles, young females, and undifferentiated eggs of H. schachtii (Smith Becker et al., 2013, 2020). Two additional sugarbeet cyst nematode-suppressive Hyalorbilia strains, HsImV25 and HsImV27, were isolated from sugar beet (Beta vulgaris) growing areas in the Imperial Valley of California (Smith Becker et al., 2024).
In California, where the SCN has not been found, the California Department of Food and Agriculture classifies H. glycines as an invasive A-rated pest with a high risk of introduction and rapid spread among 19 plant families, especially among many agriculturally important Fabaceae (CDFA, 2021; Nemaplex, 2023). This project was initiated to evaluate whether Hyalorbilia strains suppressive to the sugarbeet cyst nematode could also parasitize and suppress population development of the closely related SCN.
Hyalorbilia spp DoUCR50 and HsImV27 were isolated from parasitized sugarbeet cyst nematodes grown in soils from Agricultural Operations field 9E at the University of California, Riverside, CA, and from sugar beet fields in the Imperial Valley of California, respectively (Olatinwo et al., 2006; Smith Becker et al., 2024). The ARF18-L strain used in this study was isolated from parasitized SCN eggs collected in Tennessee (Kim et al., 1998) and was provided by Patricia Timper, USDA-ARS, Tifton, GA. Cultures of DoUCR50, HsImV27, and ARF18-L were maintained at 22°C on potato dextrose agar (PDA) with monthly transfers. Water agar cultures of fungi were prepared by transferring a 1 cm2 plug from a PDA-grown culture to the center of a water agar plate. For the peat inoculum cultures, fungi were grown in autoclaved powdered peat (BioAPT, American Peat Technology, Aitkin, MN) for 2 months as described previously (Smith Becker et al., 2024).
SCNs were collected in Michigan and provided by Marisol Quintanilla, Department of Entomology, Michigan State University, East Lansing, MI. SCN-susceptible soybean seeds (Glycine max cv. Williams 82) were provided by Thomas Maier, Department of Plant Pathology and Microbiology, Iowa State University, Ames, IA, and the USDA National Germplasm System, Beltsville, MD. Nematodes were kept in continuous culture on soybeans in the UCR Nematology Quarantine Facility greenhouse. Cysts and females of H. glycines were dislodged from the roots by a high-pressure stream of water. They were collected by flotation sieving, then placed on Baermann funnels containing 3 mM ZnCl2 and incubated at 26°C ± 3°C in the greenhouse. Hatched J2 were collected at 48 h intervals over 6 days by decanting 5–10 ml aliquots from the basal tubing of the Baermann funnels. The collected J2 were stored at 4°C before use.
Fungal peat cultures were incorporated into autoclaved sandy-loam soil (70% sand, 18% silt, 12% clay, 0.1% organic matter, pH 7.3) at a final concentration of 5 × 102 CFU/cm3 of soil. The controls consisted of autoclaved fungal peat cultures added to the soil. Some soils were amended with additional uninoculated autoclaved peat to ensure that each fungal treatment contained the same proportion of added peat. Test soils were distributed into six cones (200 cm3 per cone, Ray Leach Cone-tainer, Stuewe & Sons, Tangent, OR). Each cone was plugged at the bottom with a small fiber material (Polyester Filter Floss Media, Blue Ribbon Pet Products, Bohemia, NY). Cones were planted with two soybean seeds, arranged in a randomized complete block design in plastic racks, and incubated in a greenhouse at an average temperature of 26°C ± 4°C with ambient light. A HOBO temperature logger (Onset Computer Corporation, Bourne, MA) was used to calculate degree days (DD) with a 5°C basal temperature (Alston and Schmitt, 1988). Seedlings were thinned to one per cone after 7 days and hand-watered as needed for the first 2 weeks. Afterward, a low-pressure drip irrigation system was installed. Ungerminated seeds were removed from cones. Seedlings were fertilized weekly with liquid Miracle-Gro 20-20-20 (Scotts Company, Marysville, OH). Three weeks after planting, a glass rod was used to create two 3 cm deep holes on either side of each seedling. Each cone was infested with 250 H. glycines J2 by pipetting 500 μl of a 250 J2/ml suspension into each hole. After 1,260 DD, approximately 2 months after J2 infestation, roots and soil were carefully removed from four cones of each treatment. SCN females and cysts were dislodged from the roots by a high-pressure stream of water, extracted with those in the soil through flotation-sieving, and enumerated using a stereomicroscope. The experiment was repeated, with each treatment having four replicates per experiment. The nematode data from the two replicate nematode suppression experiments were compared using a Wilcoxon signed-rank test in Prism (GraphPad, La Jolla, CA). As there was no statistically significant difference between the medians of the two datasets (P ≤ 0.05), the values were combined for all subsequent analyses. Since some treatments did not follow a normal distribution (Shapiro–Wilk test, P ≤ 0.05), the treatments were analyzed using the non-parametric Kruskal–Wallis and Dunn’s multiple comparisons tests (P ≤ 0.05).
Water agar (1.2%) was used for the aseptic culture of soybean in 100 × 20 mm culture plates (Fisherbrand, Fisher Scientific, Pittsburgh, PA), as soybean seedling roots did not develop normally in Gamborg’s media. An 8 cm diameter circular cellophane sheet (Gel Company, San Francisco, CA) was placed on the agar surface of each plate to keep roots and nematodes at the surface of the agar and facilitate fungal inoculation of females. Prior to use, cellophane sheets were soaked in deionized water for 30 min, followed by several rinses. Ten sheets were placed in a 15 cm diameter glass Petri dish, separated by moistened sheets of 5.5 cm diameter Whatman Grade 1 filter paper. Deionized water was added to the Petri dish to cover the cellophane and filter paper, and aluminum foil was placed around the covered dish before autoclaving on the liquid cycle for 20 min. Surface sterilization of soybean seeds and H. glycines J2 was performed using the same method as for cabbage seeds and H. schachtii, respectively (Smith Becker et al., 2013). Five surface-sterilized soybean seeds were evenly spaced on each plate, and plates were sealed with Parafilm and placed on a shelf 30 cm under grow lights (Sylvania Gro-Lux 40 W, 10 h light) at 24°C. After 2 weeks, each plate was inoculated with approximately 300 surface-sterilized H. glycines J2. Once aseptic cultures of H. glycines were established, subsequent inoculations of fresh plates were achieved by transferring 5–10 aseptic cysts to new seedling plates. Fungal parasitism of developing nematode females was observed by placing a 0.5 cm2 piece of water agar-cultured DoUCR50 or HsImV27 adjacent to a female when its posterior end broke through the root epidermis. The agar block was removed after 3 days, and fungal parasitism of the female was observed under a stereomicroscope. Images of parasitized nematode females were captured using either a Zeiss stereomicroscope (SteREO Discovery.V8) equipped with a camera (Zeiss Axiocam 305) and controlled by Zeiss ZEN software (3.11) or with a Leica stereomicroscope equipped with a Canon EOS Rebel T1i/500D digital camera controlled by Canon EOS Utility version 2.6.0.
The culture of H. schachtii on cabbage (Brassica oleracea var. capitata cv. Copenhagen Market Early) in Gamborg media agar plates has been described previously (Smith Becker, 2013). An 8 cm diameter circular cellophane sheet was placed on the agar surface of each Gamborg media plate, as described above for soybean tissue culture. Seven surface-sterilized cabbage seeds were placed near the center of each plate. The plates were sealed with Parafilm and placed on a shelf 30 cm below the grow lights (Sylvania Gro-Lux 40 W, 10 h light) at 24°C. After 2 weeks, 5 mature cysts from tissue culture were transferred onto the cellophane on the agar surface of plates. Inoculation with fungi and observation of parasitism were performed as described above for H. glycines.
4-week-old cabbage (Brassica oleracea var. capitata cv. Copenhagen Market Early) and 3-week-old soybean seedlings (Glycine max cv. Williams 82) grown in cones were inoculated with 1,000 J2 of H. schachtii or H. glycines, respectively. The cones were arranged in plastic racks and incubated in the greenhouse at 26°C ± 4°C with ambient light. Young females were collected at the developmental stage when they contained exclusively undifferentiated eggs, approximately 2.5 weeks after J2 infestation for H. glycines and up to 3 weeks for H. schachtii. For each experiment, females of each nematode species were dislodged from the roots with a high-pressure stream of water and then collected by flotation sieving of the soil from two cones. Approximately 300 females were picked from sieved soil debris and transferred with forceps to a 70 μm mesh cell strainer (BD Falcon, Becton Dickinson, Franklin Lakes, NJ). The strainer was placed in 30 ml of sterile deionized water inside a 100 × 20 mm Petri plate (Fisher Scientific) containing 1.4% water agar supplemented with 100 µg/ml rifampicin and 100 µg/ml ampicillin. Females were incubated in the cell strainers on antibiotic-containing agar plates overnight at 4°C, then rinsed with 20 ml sterile deionized water by pouring the water through the strainer. Females were surface-sterilized by placing strainers in 30 ml of 0.5% sodium hypochlorite for 90 s in a Petri plate with gentle agitation. The strainers were transferred to 30 ml sterile deionized water in a Petri plate for rinsing, then moved to 20 ml of sterile deionized water in a Petri dish containing 1.4% water agar. Females were ruptured by gently crushing them with a flame-sterilized glass rod against the mesh of the cell strainer, releasing the eggs into the water. Excess water was pipetted off, and the plate was tilted to concentrate the eggs on one side. A sterile Pasteur pipette was used to transfer eggs onto the agar surface of 1-month-old water agar cultures of fungi. Egg parasitism in vitro was assessed by placing eggs individually or in groups of two or four on water agar cultures of the fungi. Eggs were moved into the groups on the agar using a flame-sterilized insect pin fastened to a Pasteur pipette. On each plate, eggs were arranged into three groups of 40 around the center of the water agar fungal culture, with groups spaced between 1.0 and 3.0 cm from the PDA agar plug. Single eggs were arranged in a 20 × 2 matrix, each spaced 0.5 cm apart. Pairs of eggs (double) were arranged in a 10 × 2 matrix, and groups of four eggs (quad) were organized in a 5 × 2 matrix, with the individual eggs in the groups spaced 0.5 cm apart. The controls consisted of eggs arranged in the same groupings on water agar without the presence of the fungi. Plates containing eggs were sealed with Parafilm and incubated at 26°C for 6 days. Parasitized eggs exhibited extensive mycelial growth, with hyphae spreading throughout the entire egg and halting further development. The total number of parasitized eggs was counted for each egg grouping on the fungal plates, and the number of eggs that developed into J2 was counted on control plates without fungi. Egg development was monitored in the control group to determine whether grouping affected development to the J2 stage, as it has been observed that undifferentiated eggs are more susceptible to fungal parasitism by Hyalorbilia spp. (Smith Becker et al., 2013). The experiments were repeated four times. The normality of data and homogeneity of variance were confirmed using Shapiro–Wilk and Levene’s tests, respectively, before applying parametric One-Way ANOVA. Fisher’s least significant difference (LSD) test (P ≤ 0.05) was used to compare the mean values.
Both DoUCR50 and HsImV27 reduced H. glycines female and cyst numbers, while ARF18-L had no effect (Fig. 1). The extent of suppression was similar to what had previously been observed for H. schachtii (Smith Becker et al., 2024), with DoUCR50 reducing the number of H. glycines females and cysts per plant by 73% and HsImV27 suppressing nematode numbers by 87%. The combination of the two fungi did not provide a higher level of suppression than HsImV27 alone (Fig. 1). The ability of Hyalorbilia spp. to parasitize both H. schachtii and H. glycines may be due to the close phylogenetic relatedness of these two cyst nematodes. An analysis of cytochrome c oxidase 1 gene sequences from cyst nematodes in the Schachtii group estimates that H. schachtii, H. glycines, and several other species diverged from one another approximately 330,000 years ago on the Iberian Peninsula (Subbotin et al., 2023). The phylogeny of the H. oviparasitica clade also indicates that these fungi are closely related (Yang et al., 2012; Smith Becker et al., 2024). The ancestral range of the hyperparasitic Hyalorbilia spp. has not been elucidated; however, the fungi have been found globally (Baral et al., 2020).
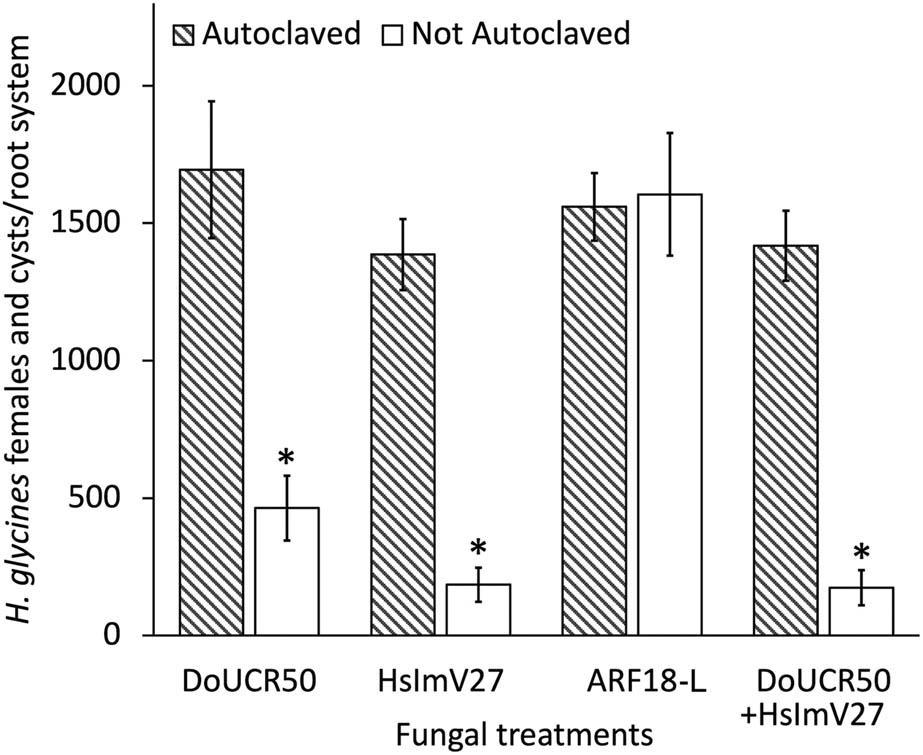
Nematode suppressive activity of Hyalorbilia spp. DoUCR50, HsImV27, and ARF-L were evaluated against Heterodera glycines on soybean (Glycine max cv. Williams 82) in soil-based greenhouse experiments. Autoclaved and not autoclaved fungal peat cultures were incorporated into autoclaved sandy-loam soil at a final concentration of 500 CFU/cm3 of soil. 3-week-old seedlings growing in cones with 200 cm3 of fungus-amended soil were inoculated with 250 H. glycines J2. Cysts and females were enumerated after approximately 2 months (1,260 DD). There was no statistically significant difference between the medians of the two data sets (Wilcoxon signed-rank test, P ≤ 0.05). The data were combined and analyzed using Kruskal–Wallis and Dunn’s multiple comparisons tests (P ≤ 0.05). Bars indicate standard error (n = 8). For each strain, * denotes a significant difference in nematode numbers at P ≤ 0.05.
The failure of ARF18-L to suppress the reproduction of the nematode species from which it was originally isolated may be due to its prolonged culture on artificial media. Wang (2003) reported that parasitism of H. glycines eggs by a strain of ARF18-L decreased from 73 to 9% after several years of axenic culture of the fungus. Parasitism could gradually be restored by culturing the fungus in pots with H. glycines and a susceptible soybean host. After 12 months of continuous culture, the level of ARF18-L egg parasitism rose from 9 to 91%. The mechanism underlying the loss and recovery of parasitism remains unknown, but this observation underscores the importance of routine virulence testing for cultured biocontrol organisms. DoUCR50 has been maintained in axenic culture for more than 20 years without loss of efficacy against H. schachtii. The periodic passage of the fungus through aseptic H. schachtii females may have helped preserve parasitic activity; however, other factors may also contribute to the stability of fungal virulence against nematodes. Current research is underway to analyze and compare whole-genome sequences of Hyalorbilia spp. to understand the molecular mechanisms behind variations in metabolic activities among strains, including differences in nematode parasitism. ARF18-L was not used in subsequent studies on nematode egg and female parasitism due to its lack of activity in the soil nematode suppression experiment.
Both DoUCR50 and HsImV27 were observed to parasitize developing females of H. schachtii and H. glycines in tissue culture (Fig. 2a–d) (Smith Becker et al., 2013). Mycelia were visible on the surface of females within 3 days of exposure to the water agar blocks of fungi, and nematode development appeared to be halted within 5–7 days. Females of both nematode species started turning brown within 10 days of exposure to the fungi.
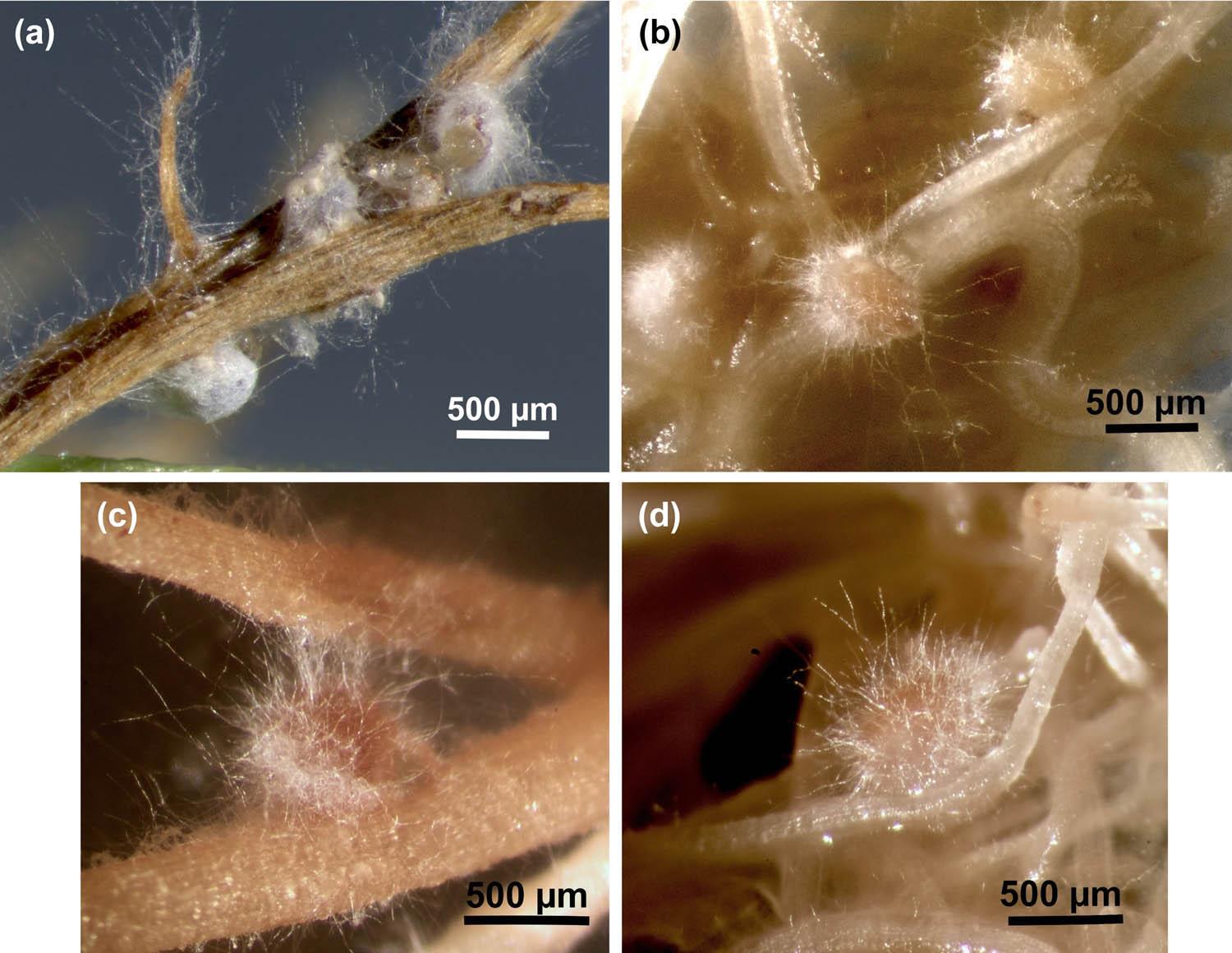
Parasitism of H. glycines and H. schachtii by HsImV27 and DoUCR50 in tissue culture. Females of H. glycines on soybean (a and c) and H. schachtii on cabbage (b and d) were inoculated with HsImV27 (a and b) or DoUCR50 (c and d) by placing a block of water agar culture of the fungus adjacent to the females as their posterior end broke through the epidermis of the roots. The blocks were removed after 3 days, and fungal parasitism was observed with a stereomicroscope. Photos were taken 10 days after fungal inoculation. Images a and b were captured using a Zeiss stereomicroscope (SteREO Discovery.V8) equipped with a camera (Zeiss Axiocam 305) and controlled by Zeiss ZEN software (3.11). The images are an extended depth of focus projection using the First/Last feature in ZEN. Images c and d were captured using a Leica stereomicroscope equipped with a Canon EOS Rebel T1i/500D digital camera controlled by Canon EOS Utility version 2.6.0. Scale bars represent 500 µm.
DoUCR50 exhibited higher egg parasitism than HsImV27 in vitro, and the susceptibility of eggs to DoUCR50 depended on the number of eggs grouped together (Fig. 3a and b). This phenomenon was initially observed fortuitously in experiments designed to quantify egg parasitism in vitro. Healthy eggs, evenly spaced on water agar cultures of DoUCR50, were not attacked by the fungus, but were parasitized when presumably non-viable. In contrast, eggs placed together in clusters on the fungal plates were invaded and killed (unpublished result). In this study, the number of parasitized nematode eggs increased significantly with increasing egg group density (P ≤ 0.05). When 40 eggs were arranged singly on fungal plates, DoUCR50 parasitized an average of 5 ± 0.9 H. glycines eggs and 6 ± 0.7 H. schachtii eggs. However, when the eggs were placed in pairs, the number of parasitized eggs increased to 17 ± 2.3 and 15 ± 2.4, respectively. This trend continued with groups of four, with parasitism reaching 25 ± 4.4 for H. glycines and 29 ± 1.1 for H. schachtii. For both nematode species, the number of eggs parasitized by DoUCR50 increased significantly with each doubling of the egg group size. In contrast, HslmV27 parasitized three or fewer eggs of either species, regardless of the group size (Fig. 3a and b). For the analyses without the fungi, an average of 36 out of 40 eggs (90%) from both nematode species successfully developed, and the grouping of eggs did not affect the number of eggs that completed development to the J2 stage (P ≤ 0.05) (data not shown).
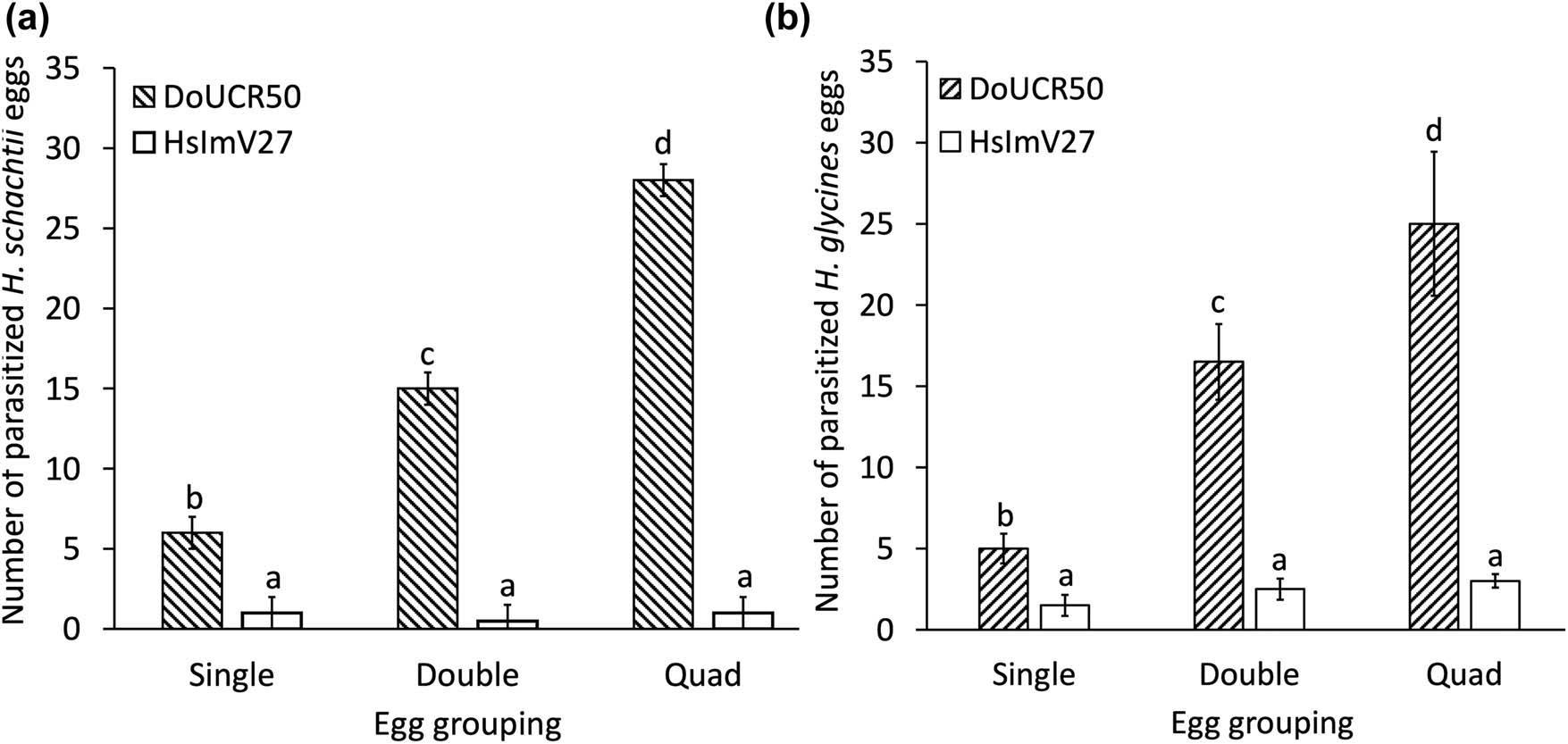
Effect of Heterodera schachtii and H. glycines egg group size on susceptibility to parasitism by Hyalorbilia sp. DoUCR50 and HsImV27. Forty undifferentiated eggs of H. schachtii (a) or H. glycines (b) were arranged on water agar cultures of the fungi in groups of 1 (Single), 2 (Double), or 4 (Quad). The total number of parasitized eggs in each group of 40 was determined after 6 days of incubation. The results of four experiments were combined, analyzed with ANOVA, and mean values compared using Fisher’s LSD (P ≤ 0.05). For each figure, a and b, different letters indicate a significant difference in the number of parasitized eggs among treatments (P ≤ 0.05). Bars represent standard error.
It was hypothesized that the ability of DoUCR50 to parasitize all eggs within H. schachtii females up to 3 weeks old was due to the induction of virulence factors such as toxins or hydrolytic enzymes (Yang et al., 2007; Smith Becker et al., 2020). The positive correlation between egg group size and the rate of parasitism by DoUCR50 suggests that the eggs may produce chemical signals or nutrients that trigger the induction of fungal virulence factors, leading to parasitism. Extracellular hydrolytic enzyme production by DoUCR50, ARF18-L, and StM, a nematophagous Hyalorbilia isolated from Meloidogyne incognita, was previously evaluated using API ZYM (bioMérieux) test strips (Smith Becker et al., 2020). Trypsin-like protease activity was uniquely detected in DoUCR50 grown on PDA and H. schachtii females, with the highest activity associated with the fungus grown on parasitized females. HsImV27 and a closely related strain from the same agricultural area, HsImV25, were also tested using the API ZYM kit, and neither fungus was positive for trypsin activity (data not shown). It remains to be determined whether the protease activity detected in DoUCR50 is an adaptation for egg parasitism.
The ability of HsImV27 to suppress H. glycines by 87% in greenhouse experiments, despite the observed lack of egg parasitism in vitro, suggests that the parasitism of developing females is the primary mode of action for this strain. DoUCR50 parasitizes both developing females and exhibits high egg parasitic activity in vitro, but it is less effective at suppressing H. schachtii and H. glycines populations in soil (Figs 1 and 3). Combining DoUCR50 and HsImV27 in greenhouse tests did not enhance suppression beyond that provided by HsImV27 alone (Fig. 1); however, longer-term studies will be necessary to determine whether any synergism exists. Factors such as fungal growth rates and persistence in soil may contribute to the variability of suppressive activity between the two strains.
ARF18 was the only Hyalorbilia spp. isolated from H. glycines cysts collected from soybean fields in ten states along the Mississippi and Missouri Rivers (Kim et al., 1998), suggesting that other members of the H. oviparasitica clade might not be present in the major soybean-growing regions of the United States. Although all 17 fungi collected in the surveys were designated as ARF18, the fungi fell into two categories with respect to colony morphology on cornmeal agar (CMA) and mitochondrial DNA restriction fragment length polymorphism. Strains designated as ARF18-C showed more compact sclerotia-like structures on CMA and higher rates of egg parasitism than strains designated as ARF18-L, and the groups had different mitochondrial DNA restriction fragment length polymorphism. Despite extensive sampling of SCN from soybean-growing regions throughout the Midwest, ARF18 was only found south of 37°N latitude. The authors speculated that the soil temperatures and the longer history of H. glycines in the South, or differences in soybean cultivars grown, might explain this geographic boundary. López et al. (2024) used amplicon sequencing of the 18S-ITS rDNA regions of DNA extracted from soil to evaluate the effect of H. glycines population density on fungal communities from 26 counties in Ohio. Although they observed enhanced activity in two fungi with potentially nematophagous activity, they did not report the occurrence of Hyalorbilia spp. in the samples. The apparent absence of Hyalorbilia spp. in major soybean-growing regions north of 37° N latitude raises the possibility that introducing these fungi might help reduce SCN populations and protect soybeans from nematode damage, provided that state and federal regulations permit such introduction. The ability of highly effective Hyalorbilia spp., isolated from H. schachtii in California agricultural areas, to parasitize the more commercially damaging H. glycines could provide an opportunity to expand the range of these effective biocontrol agents.
This work is supported by the Pests and Beneficial Species in Agricultural Production Systems Program, project award nos. 2018-67014-28066 and 2023-67014-39161, and by Hatch and Multistate Hatch Programs, project nos. 7007404, 7007401, and 7006441 from the U.S. Department of Agriculture’s National Institute of Food and Agriculture. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
The authors contributed equally.
Authors state no conflict of interest.