
Root-knot nematodes (Meloidogyne spp.) are among the most devastating plant-parasitic nematodes worldwide, causing substantial economic losses in cucurbit cultivation (Jones et al., 2017; Sikora et al., 2022). These nematodes infect plant roots, inducing gall formation that disrupts water and nutrient uptake, leading to stunted growth, diminished yields, and poor fruit quality (Khan and Tanaka, 2023). Meloidogyne incognita is notably prevalent globally and highly polyphagous, posing a significant challenge to sustainable cucurbit production (Bui and Desaeger, 2022).
Watermelon (Citrullus lanatus) and sweet melon (Cucumis melo) are economically important cucurbit crops highly susceptible to M. incognita infestations. Even low nematode population densities can markedly impair plant growth and productivity (Chahar et al., 2021; Patel et al., 2023). Notably, inoculum levels as low as 10 second-stage juveniles (J 2) per plant reduce vine length and biomass in cucumbers, underlining the critical need for effective nematode management strategies.
Grafting susceptible scions onto resistant rootstocks represents a promising and eco-friendly approach to mitigate root-knot nematode damage. Resistant rootstocks confer enhanced tolerance by restricting nematode penetration and reproduction through both physical barriers and induced biochemical defenses (García-Mendívil and Sorribas, 2021; Mukhtar and Hussain, 2023; McKenry and Anwar, 2006). However, grafting success depends on rootstock-scion compatibility and the rootstock’s capacity to withstand biotic and abiotic stresses (Jayswal and Lal, 2020).
Biochemical defense responses, mediated by key enzymes, such as peroxidase (POD), polyphenol oxidase (PPO), and phenylalanine ammonia-lyase, are integral to plant resistance against nematodes. These enzymes facilitate cell wall reinforcement via lignin and phenolic synthesis, generate reactive oxygen species, and produce antimicrobial compounds, thereby limiting pathogen progression (Ngadze et al., 2012). Increased activities of POD and PPO have been correlated with reduced M. incognita reproduction and enhanced host tolerance in cucurbits (Chandrawat et al., 2020; Khalifa et al., 2024).
Despite these advances, a comprehensive understanding of how defense enzyme activities respond to varying M. incognita initial population densities in grafted cucurbit hybrids remains incomplete. Investigating these biochemical responses in conjunction with nematode reproduction and plant growth under differing inoculum pressures is essential to optimize grafting strategies and refine integrated pest management practices.
While the relationship between inoculum level density and disease severity is well established, the integrated physiological, biochemical, and nematological responses of grafted cucurbit systems under varying inoculum pressure remain less explored. This study uniquely evaluates how grafting onto resistant root stocks influences not only M. incognita reproduction and plant growth but also key defense enzyme activities across a gradient of nematode densities, providing a mechanistic insight into rootstock-mediated resistance.
A pure culture of M. incognita was maintained on tomato plants (Solanum lycopersicum Mill.) cv. Super Strain B, following the method of Abdel-Rahman et al. (2023). The culture was sustained under greenhouse conditions at the Department of Zoology and Agricultural Nematology (Nematology Division), Faculty of Agriculture, Cairo University. Species identification was confirmed by morphological examination of perineal patterns from adult females (Khalifa et al., 2024), and race determination was conducted according to the differential host test of Taylor and Sasser (1978). Tomato cultivar susceptibility profiles confirmed the population as M. incognita race 1.
For inoculum preparation, heavily galled roots were collected from infested tomato plants. Egg masses were isolated using the sodium hypochlorite method as described by Khalifa et al. (2024), a modification of the Hussey and Barker (1973) protocol using 1% NaOCl with gentle shaking for 4 min, and incubated in water at room temperature to obtain freshly hatched second-stage juveniles (J 2). Juvenile counts were made under a stereomicroscope to standardize inoculum density.
Certified seeds of watermelon (Citrullus lanatus) cv. Aswan F1, sweet melon (Cucumis melo) cv. Faransawy, two commercial rootstocks (6001 F1 and BS F1), and their grafted combinations (Aswan F1/6001 F1, Aswan F1/BS F1) were obtained from Roots Nurseries, km 75 Cairo–Alexandria Desert Road, Egypt. Resistance categories highly resistant (HR), resistant (R), or moderately susceptible (MS) were assigned according to preliminary screening data (Hassnin et al., 2023).
Seedlings were transplanted into 20 cm diameter clay pots containing approximately 2 kg sterilized sandy-loam soil mixture (1:1, v/v). Four inoculum levels of freshly hatched M. incognita J 2 1,000, 2,000, 4,000, and 8,000 juveniles/plant were applied in a 25 mL water suspension. The suspension was pipetted into four evenly spaced holes around the root zone. Control plants received no nematode inoculation. Treatments were arranged in a completely randomized design with four replicate pots per treatment. Plants were grown on sterilized benches in a greenhouse (27 ± 2°C; 60–70% relative humidity (RH)) under uniform horticultural care. The experiment was conducted twice under identical greenhouse conditions with consistent results; data presented are from one representative trial with four replicate pots per treatment.
Forty-five days after inoculation, shoot and root lengths and fresh weights were recorded. Infected root systems were washed free of soil and fixed in formalin–acetic acid–alcohol. Nematode parameters were evaluated as follows:
-
Number of galls, females, and egg masses per root (Hussey and Barker, 1973)
-
Number of J 2 in 250 g soil (extracted by the modified Baermann funnel method; Hooper et al., 2005)
-
Final population (Pf) = nematodes in roots + soil. Note: Pf included only females/ egg masses in roots + hatched J 2 from soil. Unhatched eggs within egg masses were not counted separately as they were considered part of the reproductive potential already accounted for in root assessments.
-
Reproduction factor (Rf) = Pf/Pi.
-
Penetration rate (%R.P.) = (No. of developmental stages and females in root/Pi) × 100. Roots were stained with 0.1% acid fuchsin to visualize and count developmental stages and females prior to assessment.
-
Reproduction rate (%R.R.) = (egg masses/root population) × 100.
Defense-related enzyme activities were assayed in fresh shoot and root tissues (1 g/sample). Samples were homogenized in 10 mL of ice-cold 0.1 M phosphate buffer and centrifuged at 10,000 ×g for 15 min at 4°C. The resulting supernatants were used for spectrophotometric assays.
POD activity was determined following the method by Hammerschmidt et al. (1982) in a reaction mixture containing phosphate buffer, 0.3% H2O2, and 0.1% guaiacol; absorbance was measured at 470 nm after 5 min incubation at room temperature.
PPO activity was measured according to Mayer and Harel (1965) using catechol (0.1%) as a substrate; absorbance was read at 420 nm after 5 min incubation.
Enzyme activities were expressed as µg substrate converted min−1 g−1 fresh weight (µg min−1 g−1 FW).
Data were analyzed using SPSS version 12 (SPSS Inc., 2003). A two-way ANOVA tested the main effects of genotype and inoculum level, and their interaction (genotype × inoculum). Mean comparisons were performed using Duncan’s multiple range test (Duncan, 1955) at P ≤ 0.05.
Based on previous screening results, watermelon cv. Aswan F1 (MS), and the rootstocks 6001 F1 (HR) and BS F1 (R), together with their grafted combinations, were evaluated under five M. incognita inoculum levels (0, 1,000, 2,000, 4,000, and 8,000 J 2/plant).
Across all host genotypes, nematode reproduction parameters increased significantly (P < 0.001) with rising inoculum density (Table 1). The susceptible watermelon cultivar Aswan F1 showed the highest Pf/Pi ratios at 2,000 J 2/plant (7.51) and 4,000 J 2/plant (4.73). In contrast, the resistant rootstocks 6001 F1, BS F1, along with their grafted combinations maintained significantly lower Pf/Pi values. For instance, grafting Aswan F1 onto BS F1 reduced Pf/Pi by 68% compared to non-grafted Aswan F1 at 8,000 J 2/plant (P < 0.001). Similar trends were observed for gall, female, and egg mass counts.
Meloidogyne incognita reproduction parameters on watermelon, rootstock hybrids, and grafted combinations at different inoculum levels
| Hybrids | Inoculum level | Galls/root | Females/root | Egg-masses/root | Pf | Pf/Pi | %R.P. | %R.R. |
|---|---|---|---|---|---|---|---|---|
| 6001 F1 (HR) | 1,000 | 20 K | 12 O | 18 N | 252 Q | 0.25 M | 3.0 | 60.0 |
| 2,000 | 91 J | 71 N | 81 L | 1,360 NO | 0.68 JK | 7.6 | 53.3 | |
| 4,000 | 171 H | 128 KL | 150 I | 2,520 LM | 0.63 K | 7.0 | 54.0 | |
| 8,000 | 272 G | 154 JK | 163 I | 3,261 K | 0.41 L | 4.0 | 51.4 | |
| BS F1 (R) | 1,000 | 51 K | 16 O | 39 M | 706 P | 0.71 JK | 5.5 | 70.9 |
| 2,000 | 181 H | 164 JK | 73 L | 2,743 L | 1.37 H | 11.9 | 30.8 | |
| 4,000 | 261 G | 258 H | 185 H | 4,662 I | 1.17 I | 11.1 | 41.8 | |
| 8,000 | 392 F | 319 F | 181 H | 5,850 H | 0.73 J | 6.3 | 36.2 | |
| ASWAN F1 (MS) | 1,000 | 709 E | 405 E | 210 G | 1,550 N | 1.55 G | 61.5 | 34.1 |
| 2,000 | 827 D | 685 D | 469 D | 15,012 D | 7.51 A | 57.7 | 40.6 | |
| 4,000 | 983 C | 815 C | 688 C | 18,903 C | 4.73 B | 37.6 | 45.8 | |
| 8,000 | 1325 A | 994 A | 1,147 A | 21,421 A | 2.68 C | 26.8 | 53.6 | |
| ASWAN F1/6001 F1 (HR) | 1,000 | 107 J | 79 MN | 41 M | 382 Q | 0.38 L | 12.0 | 34.2 |
| 2,000 | 127 IJ | 95 LMN | 111 K | 2,402 M | 1.20 I | 10.3 | 53.9 | |
| 4,000 | 152 HI | 114 LM | 134 J | 6,362 G | 1.59 G | 6.2 | 54.0 | |
| 8,000 | 395 F | 297 FG | 346 E | 11,248 E | 1.41 H | 8.0 | 53.8 | |
| ASWAN F1/BS F1 (MS) | 1,000 | 391 F | 223 I | 79 L | 1,142 O | 1.14 I | 30.2 | 26.2 |
| 2,000 | 246 G | 185 J | 216 J | 3,870 J | 1.94 E | 20.1 | 53.9 | |
| 4,000 | 359 F | 269 GH | 314 F | 7,287 F | 1.82 F | 14.6 | 53.9 | |
| 8,000 | 1,039 B | 933 B | 836 B | 19,466 B | 2.43 D | 22.1 | 47.3 | |
| SE± | 40.867 | 33.245 | 32.653 | 754.085 | 0.186 |
Mean values followed by the same letter(s) within a column in each block are not significantly different (P ≤ 0.05) according to Duncans׳ multiple range tests. Pf = Final population = Embedded stages + Soil population. %R.P. = Rate of Penetration = (Root population/Initial population) × 100. %R.R. = Rate of Reproduction = (Number of egg masses/Root population) × 100. SE± = Standard error.
Across all genotypes, female and egg-mass counts rose significantly with increasing inoculum pressure, with the largest values occurring at 8,000 J 2/plant in Aswan F1. Penetration rate (%R.P.) generally increased with inoculum level but was lowest in resistant hosts and highest in Aswan F1 at 8,000 J 2/plant. Reproduction rate (%R.R.) gradually declined with higher inoculum loads in rootstocks, but in Aswan F1 and its grafts, %R.R. tended to remain higher or decline less sharply.
Regression analysis indicated no consistent relationship between inoculum level and Pf/Pi in the rootstocks alone but a positive association in the grafted accessions (Aswan F1/6001 F1 and Aswan F1/BS F1) (Fig. 1).
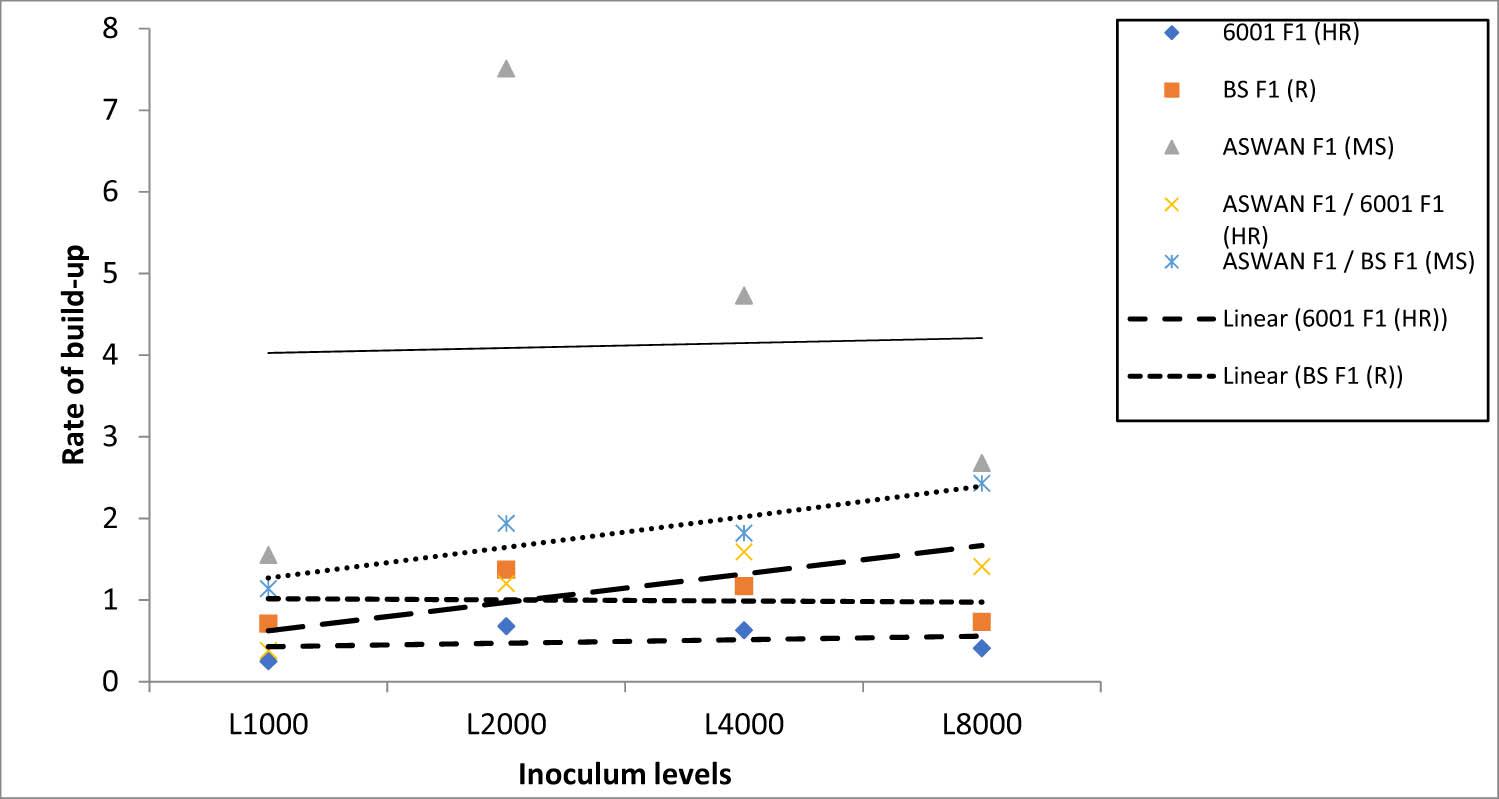
Correlation between initial inoculum levels and population build-up of Meloidogyne incognita on watermelon, rootstock hybrids, and grafted combinations.
All inoculum levels significantly suppressed growth parameters, with maximum inhibition observed at 8,000 J 2/plant (Figs 2 and 3). Shoot and root FWs decreased progressively with increasing inoculum in all genotypes except Aswan F1/BS F1, which maintained relatively higher shoot weights, even at higher nematode pressures. Root weight reductions were generally more pronounced than shoot weight reductions.

Shoot FW reduction (%) in watermelon and rootstock hybrids at different inoculum levels of Meloidogyne incognita.

Root FW reduction (%) in watermelon and rootstock hybrids at different inoculum levels of Meloidogyne incognita.
At each inoculum level, non-grafted 6001 F1, BS F1, and Aswan F1 suffered greater percentage losses in root weight compared with grafted combinations. Aswan F1/BS F1 exhibited the smallest reductions in both shoot and root biomass, followed by Aswan F1/6001 F1. Across hybrids, the greatest shoot-weight loss was at 8,000 J 2/plant, while the least occurred at 1,000 J 2/plant. The partial resistance observed in grafted combinations is characterized by reduced nematode penetration, impaired development, and enhanced biochemical defenses, consistent with rootstock-mediated resistance mechanisms. Even resistant rootstocks showed mild growth reduction under high inoculum pressure, indicating that resistance does not confer complete immunity but rather reduces damage through physiological and biochemical tolerance mechanisms.
For sweet melon cv. Faransawy (MS), rootstocks 6001 F1 (HR) and BS F1 (R), and their grafted accessions, reproductive criteria also increased with inoculum density (Table 2). Faransawy supported the highest nematode reproduction and Pf/Pi, followed by Faransawy/BS F1 and Faransawy/6001 F1. At inoculum levels ≥2,000 J 2/plant, Faransawy exhibited markedly elevated nematode counts in both roots and soil. The 6001 F1 rootstock maintained the lowest Pf at all inoculum levels.
Meloidogyne incognita reproduction parameters on sweet melon, rootstock hybrids, and grafted combinations at different inoculum levels
| Hybrids | Inoculum level | Galls/root | Females/root | Egg-masses/root | Pf | Pf/Pi | %R.P. | %R.R. |
|---|---|---|---|---|---|---|---|---|
| 6001 F1 (HR) | 1,000 | 20 M | 12 J | 18 O | 252 Q | 0.25 O | 3.0 | 60.0 |
| 2,000 | 91 K | 71 I | 81 M | 1,360 P | 0.68 LM | 7.6 | 53.3 | |
| 4,000 | 171 J | 128 H | 150 L | 2,520 L | 0.63 M | 7.0 | 54.0 | |
| 8,000 | 272 I | 154 H | 163 KL | 3,261 K | 0.41 N | 4.0 | 51.4 | |
| BS F1 (R) | 1,000 | 51 L | 16 J | 39 N | 343 Q | 0.34 NO | 5.5 | 70.9 |
| 2,000 | 181 J | 164 H | 73 M | 2,743 L | 1.37 J | 11.9 | 30.8 | |
| 4,000 | 261 I | 258 G | 185 J | 4,662 I | 1.17 K | 11.1 | 41.8 | |
| 8,000 | 392 G | 319 F | 181 JK | 5,850 H | 0.73 L | 6.3 | 36.2 | |
| Faransawy (MS) | 1,000 | 777 E | 485 D | 213 I | 2,219 M | 2.22 D | 69.8 | 30.5 |
| 2,000 | 942 D | 763 C | 586 D | 17,377 C | 8.69 A | 67.5 | 43.4 | |
| 4,000 | 1,327 B | 937 B | 860 B | 21,165 B | 5.29 B | 44.9 | 47.9 | |
| 8,000 | 1,507 A | 1,072 A | 945 A | 23,686 A | 2.96 C | 25.2 | 46.9 | |
| Faransawy/6001 F1 (R) | 1,000 | 561 F | 447 E | 348 E | 1,642 O | 1.64 I | 79.5 | 43.8 |
| 2,000 | 566 F | 322 F | 305 G | 2,628 L | 1.31 J | 31.4 | 48.6 | |
| 4,000 | 338 H | 237 G | 227 I | 6,578 G | 1.64 I | 11.6 | 48.9 | |
| 8,000 | 328 H | 356 F | 329 F | 11,290 E | 1.41 J | 8.6 | 48.0 | |
| Faransawy/BS F1 (MS) | 1,000 | 260 I | 472 DE | 256 H | 1,890 N | 1.89 G | 72.8 | 35.2 |
| 2,000 | 271 I | 319 F | 228 I | 4,211 J | 2.11 E | 27.4 | 41.7 | |
| 4,000 | 388 G | 236 G | 261 H | 7,202 F | 1.80 H | 12.4 | 52.5 | |
| 8,000 | 1,053 C | 943 B | 837 C | 15,937 D | 1.99 F | 22.3 | 47.0 | |
| SE± | 46.348 | 34.472 | 30.178 | 783.694 | 0.216 |
Mean values followed by the same letter(s) within a column in each block are not significantly different (P ≤ 0.05) according to Duncans’ multiple range tests. Pf = Final population = Embedded stages + Soil population. %R.P. = Rate of Penetration = (Root population/Initial population) × 100. %R.R. = Rate of Reproduction = (Number of egg masses/Root population) × 100. SE± = Standard error.
Faransawy recorded its maximum Pf/Pi values at 2,000 J 2/plant (8.69), 4,000 J 2/plant (5.29), and 8,000 J 2/plant (2.96). Female and egg mass numbers peaked at 4,000 and 8,000 J 2/plant for Faransawy, and at 8,000 J 2/plant for Faransawy/BS F1.
Correlation analyses showed no relationship between inoculum level and Pf/Pi in 6001 F1, BS F1, or Faransawy/BS F1. In contrast, a negative correlation was found for Faransawy and Faransawy/6001 F1, where increasing inoculum levels reduced Pf/Pi (Fig. 4).

Correlation between initial inoculum levels and population build-up of Meloidogyne incognita on sweet melon, rootstock hybrids, and grafted combinations.
All inoculation levels ≥2,000 J 2/plant significantly reduced shoot and root biomass in sweet melon hybrids (Figs 5 and 6). Losses increased with inoculum level but were less severe in grafted plants. Notably, Faransawy/6001 F1 exhibited increases in root FW at the lower inoculum levels (1,000–4,000 J 2/plant), while Faransawy/BS F1 showed similar increases at 1,000–2,000 J 2/plant. In most cases, root weight reductions exceeded shoot reductions, and the magnitude of reduction was genotype-dependent. The observed partial resistance in grafted sweet melon combinations similarly reflects rootstock-mediated defense mechanisms that limit nematode success while maintaining plant growth.

Shoot FW reduction (%) in sweet melon and rootstock hybrids at different inoculum levels of Meloidogyne incognita.

Root FW reduction (%) in sweet melon and rootstock hybrids at different inoculum levels of Meloidogyne incognita.
The largest biomass decreases occurred at 8,000 J 2/plant in all hybrids, whereas 1,000 J 2/plant caused the smallest losses. The Faransawy cultivar had the highest percentage reductions in root biomass, followed by BS F1, while Faransawy/6001 F1 exhibited the smallest losses. As observed in watermelon, even resistant sweet melon rootstocks displayed some growth reduction at the highest inoculum level, reinforcing that resistance represents damage mitigation rather than complete immunity.
ANOVA (Table 3) confirmed significant effects (P < 0.001) of hybrid genotype, inoculum level, and their interaction on nematode reproduction and plant growth parameters. Resistant rootstocks (6001 F1, BS F1) exhibited consistently lower Pf and %R.P., higher %R.R. stability, and reduced growth suppression compared with susceptible cultivars (Aswan F1, Faransawy). Susceptible cultivars showed steep increases in Pf and pronounced growth losses under high nematode pressure.
ANOVA summary for effects of genotype, inoculum level, and interaction on nematode and plant growth traits
| Source of variation | Galls/root | Females/root | Egg-masses/root | Pf | P/Pi | %R.P. | %R.R. |
|---|---|---|---|---|---|---|---|
| Hybrids | 45.23 (<0.001) | 38.76 (<0.001) | 42.15 (<0.001) | 50.12 (<0.001) | 12.34 (<0.001) | 25.67 (<0.001) | 18.45 (<0.001) |
| Inoculum levels | 67.89 (<0.001) | 55.43 (<0.001) | 60.78 (<0.001) | 72.34 (<0.001) | 15.67 (<0.001) | 30.12 (<0.001) | 22.34 (<0.001) |
| Hybrids × inoculum | 12.34 (<0.001) | 10.56 (<0.001) | 11.23 (<0.001) | 14.56 (<0.001) | 5.67 (<0.001) | 8.45 (<0.001) | 7.89 (<0.001) |
The F-values and P-values indicate significant interactions between hybrids and inoculum levels for all measured parameters (P < 0.001).
This suggests that both the hybrid type and inoculum level significantly influence the development and reproduction of M. incognita.
In watermelon cv. Aswan F1, POD and PPO activities increased progressively with inoculum density (Fig. 7). At 8,000 J 2/plant, POD and PPO activities were 10.2 ± 0.7 U g−1 FW and 6.1 ± 0.6 U g−1 FW, respectively (P < 0.01).

POD and PPO activities in watermelon and rootstock hybrids at different inoculum levels of Meloidogyne incognita (mean value ± SD, n = 4).
Grafted combinations (Aswan F1/6001 F1, Aswan F1/BS F1) exhibited significantly higher baseline and induced enzyme activities than non-grafted Aswan F1. At 8,000 J 2/plant, Aswan F1/6001 F1 recorded POD activity of 24.5 ± 2.3 µg H2O2 min−1 g−1 FW and PPO activity of 16.8 ± 1.2 µg catechol min−1 g−1 FW (P < 0.01), with slightly lower but still elevated values in Aswan F1/BS F1.
Similar patterns were observed in sweet melon cv. Faransawy (Fig. 8). POD and PPO activities increased significantly with inoculum level, peaking at 9.0 ± 0.6 U g−1 FW and 5.5 ± 0.5 U g−1 FW, respectively, in non-grafted plants at 8,000 J 2/plant.

POD and PPO activities in sweet melon, and rootstock hybrids at different inoculum levels of Meloidogyne incognita (mean value ± SD, n = 4).
Grafted combinations showed substantially higher constitutive and induced enzyme levels. At 8,000 J 2/plant, Faransawy/6001 F1 reached 21.5 ± 2.1 µg H2O2 min−1 g−1 FW (POD) and 14.5 ± 1.0 µg catechol min−1 g−1 FW (PPO), outperforming their non-grafted counterpart (P < 0.01).
Across both cucurbit crops, increasing M. incognita inoculum levels generally enhanced nematode reproduction and enzyme activity, while suppressing plant growth. Resistant rootstocks and grafted combinations consistently moderated nematode multiplication, limited growth reductions, and boosted defense enzyme activities compared to susceptible cultivars, even under high nematode pressure.
The study clearly demonstrates that resistant rootstocks, such as 6001 F1 (HR) and BS F1 (R), effectively suppress M. incognita multiplication across a range of inoculum levels, maintaining their resistance even at high nematode pressures (up to 8,000 J 2/plant). This suggests strong potential for practical use in heavily infested fields. In contrast, MS cultivars like Aswan F1 and Faransawy, as well as their grafted versions, suffer greater reductions in plant growth, particularly in shoot and root fresh weights, due to extensive root damage from high nematode populations. This damage disrupts key physiological processes, including photosynthesis, carbon metabolism, and nutrient uptake, in line with previous findings (Mateille et al., 1995; Khan et al., 2019; Chahar et al., 2021).
Nematode population dynamics show that susceptible plants support successful juvenile parasitism through the formation of giant cells in vascular tissues, which are essential for nematode development. Conversely, resistant hybrids inhibit giant cell formation, leading to juvenile mortality or root abandonment. Differences in nematode reproduction metrics, including egg mass counts and reproduction factors, between resistant and susceptible types reflect underlying genetic variation in host resistance and nematode adaptability (Griffin, 1982; Castagnone-Sereno, 2006). MS hybrids typically had elevated egg masses and reproduction rates, while resistant cultivars showed limited nematode development.
The resistance mechanisms described including hypersensitive response, and giant cell inhibition, which effectively suppressed nematode multiplication and maintained growth under high inoculum pressure (Dropkin, 1969; Nelson et al., 1990). Notably, nematode multiplication rates were paradoxically higher at lower inoculum densities, probably due to reduced intraspecific competition and more abundant resources in less damaged roots (Haynes and Jones, 1976; Bendezu and Starr, 2003). High nematode populations restrict subsequent juvenile infection sites, limiting reproduction (Ogunfowora, 1977; Oostenbrink, 1966).
This study confirms the complex interplay among host resistance, nematode genetics, and environmental conditions in determining nematode population dynamics and plant health outcomes. Importantly, growth parameters were significantly influenced by final nematode populations and gall severity, corroborating earlier research on nematode pathogenicity thresholds.
On the biochemical defense front, key enzymes POD and PPO showed significant positive correlations with inoculum density in both watermelon cv. Aswan F1 and sweet melon cv. Faransawy. These enzymes are integral to plant defense, contributing to reactive oxygen species generation, cell wall fortification, and antimicrobial compound synthesis (Almagro et al., 2009; Constabel and Barbehenn, 2008). The increased enzyme activities at higher nematode inoculum levels indicate an inducible defense response aligned with previous reports linking elevated POD and PPO to nematode resistance and better plant health (Elsharkawy et al., 2022; Chandra et al., 2010).
Differences in enzyme activity between the two cucurbit crops suggest that watermelon possesses more robust biochemical defense mechanisms compared to sweet melon, consistent with their relative resistance profiles (López-Gómez et al., 2016). The clear statistical distinctions in enzyme responses across inoculum levels underscore their potential as biomarkers for plant health and resistance, supporting their use in integrated pest management strategies that harness natural defense pathways. While this study was conducted under controlled greenhouse conditions, the findings have important implications for field management. In our pot system (2 kg soil), 8,000 J 2/plant corresponded approximately to 4,000 J 2/ 250 g soil. However, field equivalency depends on root volume, soil type, and environmental factors. We recommended that future field trials establish soil-density-based thresholds for these rootstocks. For growers, pre-plant soil sampling is advised to estimate nematode pressure, with grafted plants recommended for fields with known M. incognita history, particularly at densities exceeding 2,000 J 2/250 g soil.
Together, these findings confirm the efficacy of resistant rootstocks and grafting in mitigating root-knot nematode damage, highlight the importance of initial nematode population management, and emphasize the valuable role of defense enzymes in cucurbit resistance. The results also reinforce the need for rootstock specificity when selecting for nematode resistance, echoing studies such as García-Mendívil and Sorribas (2021) while accounting for some contradictory findings (Giné et al., 2017).
Overall, the study advances understanding of nematode-host interactions in grafted cucurbits and provides practical insights for improving nematode management through resistant rootstock deployment and biochemical defense monitoring.
This study demonstrates that resistant cucurbit rootstocks (6001 F1 and BS F1) significantly suppress M. incognita reproduction and mitigate growth damage compared to susceptible cultivars. Grafting MS cultivars onto resistant rootstocks enhances tolerance and reduces nematode population build-up. Increased POD and PPO activities in resistant and grafted plants suggest key roles for biochemical defenses in nematode resistance. These findings support integrating genetic resistance, grafting, and biochemical monitoring in sustainable root-knot nematode management for cucurbit production. Future research should include multi-location field trials with higher replication to establish economic thresholds and validate the practical efficacy of tested rootstocks under varying environmental conditions.
The authors thank the Deanship of Scientific Research, King Faisal University, for funding support (grant KFU260588) and the Faculty of Agriculture, Cairo University, for providing greenhouse and laboratory facilities.
This work was supported by the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia (grant number KFU260588).
Hosny Kesba and Ali Ali conceived and designed the study; Sherif El-Ganainy and Nourhan Hassnin performed data analysis, with Nourhan Hassnin conducting the investigation and data validation; Hosny Kesba provided resources, secured funding, and supervised the project, while Ali Ali managed project administration; Sherif El-Ganainy developed the methodology and handled software tasks. All authors contributed to drafting the manuscript, with Nourhan Hassnin and Sherif El-Ganainy preparing the original draft, and Hosny Kesba and Ali Ali reviewing and editing the final version.
No potential conflict of interest was reported by the author(s).
The data that support the findings of this study are available from the corresponding author upon reasonable request.