
Papaya (Carica papaya L.), a tropical fruit crop of the Caricaceae family, is cultivated extensively in tropical and subtropical regions worldwide. In 2020, papaya was the third most produced tropical fruit crop in the world (Burns et al., 2022). It is the fourth in terms of global exports after mango, pineapple, and avocado (FAO, 2023) and serves as an important source of vitamins and minerals. In recent years, global papaya production has risen significantly, making it an increasingly important agricultural export commodity – especially for developing countries in Asia and Latin America – where it supports the livelihoods of thousands of smallholder farmers (Evans and Ballen, 2012).
Despite its growing economic importance, papaya production is constrained by several biotic stresses, among which plant-parasitic nematodes (PPN) are particularly damaging. Several nematode genera have been reported in association with papaya worldwide (Khan et al., 2007; Perera et al., 2008; Cornejo-Condori et al., 2021; Indarti and Taryono, 2023). The most widespread and economically damaging nematodes in papaya fields are species of the genera Meloidogyne and Rotylenchulus (El-Borai and Duncan, 2005; Dias-Arieira et al., 2008; Plena et al., 2022), with yield losses estimated between 15% and 20% (Koenning et al., 1999). Indeed, Meloidogyne javanica may develop specific feeding sites in galls and efficiently reproduce in papaya roots (Coulibaly et al., 2025). Nowadays, molecular identification techniques can assist field surveys to identify the most important PPN species in productive areas (Plena et al., 2022; Brenes-Campos et al., 2025; Wanjala et al., 2025). Molecular identification of PPNs typically relies on polymerase chain reaction (PCR) methods for amplification and sequencing of diagnostic loci, such as the internal transcribed spacer (ITS) region or the 18S small subunit rRNA gene (Floyd et al., 2002). In addition, species-specific primers have been developed to support species discrimination, particularly for Meloidogyne and Rotylenchulus species (Zijlstra et al., 2000; Randig et al., 2002; Van den Berg et al., 2016).
In Burkina Faso, papaya cultivation is expanding, with production reaching 18,939 tons in the 2020/2021 season (DGESS, 2019). The crop is grown under various agroecological conditions and farming systems. However, despite the increasing relevance of papaya in the country, little research is known about the parasitic nematode communities associated with its cultivation.
This study aims to address this knowledge gap by (i) identifying the nematode genera associated with papaya in major production areas of Burkina Faso, (ii) characterizing species within the Meloidogyne and Rotylenchulus genera, and (iii) assessing the cropping systems and agroecological factors influencing nematode population densities. In this study, we present the results of a comprehensive field survey conducted in 61 papaya orchards, along with the characterization of nematode genera and species using both morphological and molecular tools.
The study was conducted in papaya orchards across Burkina Faso during two sampling periods: October 2018 and July 2021. Nine administrative regions representing the main papaya production areas (Figure 1) and comprising 29 localities were surveyed. A total of 61 papaya orchards were surveyed, comprising 26 in 2018 and 45 in 2021. With the exception of seven sites, all orchards had been recently established. The age of papaya trees varied among orchards, ranging from 2 months to 36 months. These regions span the Sudanian and Sudanian-Sahelian agro-climatic zones, which show spatial and temporal variation in rainfall, decreasing along a south-to-north gradient. The Sudanian-Sahelian zone receives < 900 mm of annual rainfall, whereas the Sudanian zone receives between 900 mm and 1,200 mm.
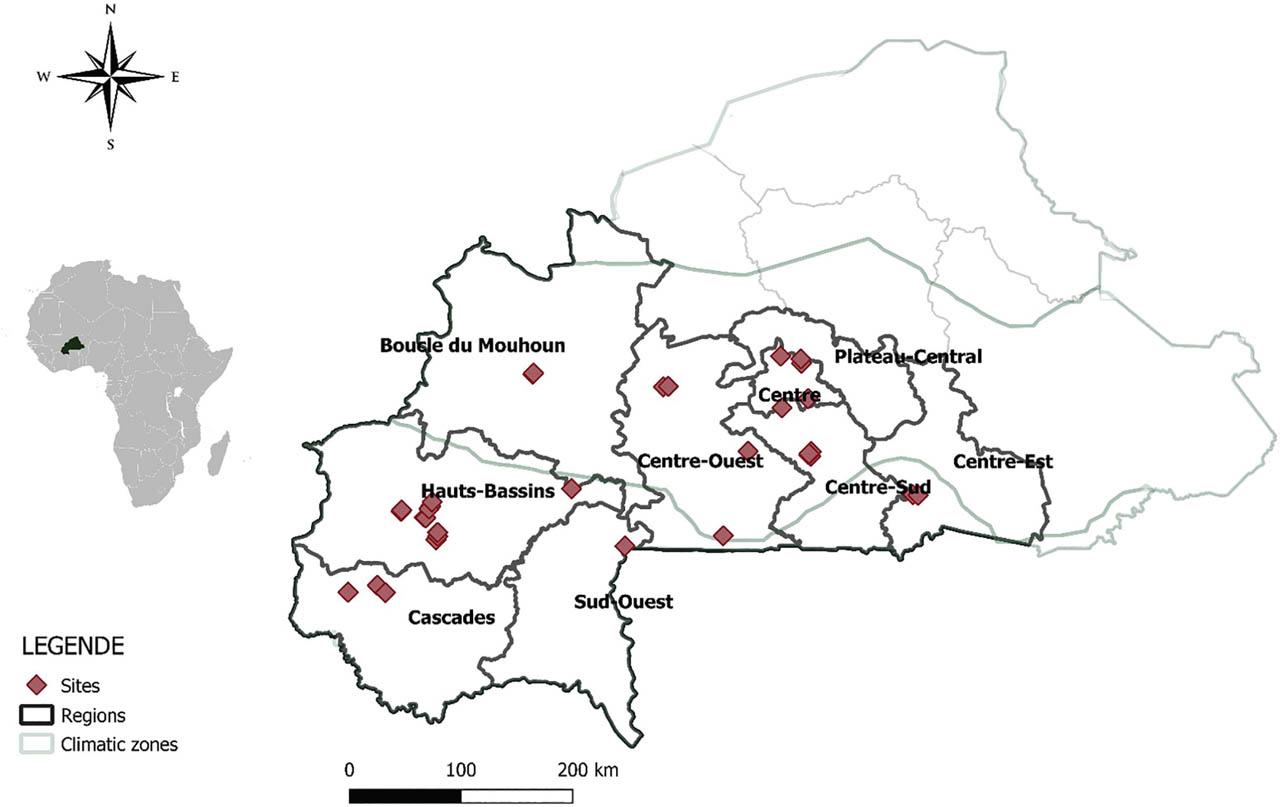
Map of Burkina Faso (administrative map) indicating the sampling sites across nine regions where nematological samples were collected.
A total of 207 composite soil and root samples were collected from farmers' papaya orchards. Samples were taken from cultivars including ‘Solo’, ‘FBPA1’, and other locally grown but unidentified varieties. A minimum of three samples per orchard were collected. For each sampling site, metadata such as date, GPS coordinates, region, province, locality, cultivar type, cropping history, and intercropping practices were recorded. Samples were transported to the nematology laboratory of Institut de ľEnvironnement et de Recherches Agricoles (INERA) in Farako-Ba for morphological analysis of soil and root nematodes.
Soil nematodes were extracted using the Seinhorst elutriator method (Seinhorst, 1962), and root nematodes were extracted using the Seinhorst sprinkler method (Seinhorst, 1950). After extraction, nematodes were observed under an inverted microscope (Hund Wilovert, Serial# 202139 (Wetzlar, DE, Germany) and morphologically identified using the taxonomic key of Mai and Lyon (1975). Nematode populations were quantified as number per cubic decimeter (n/dm3) of soil or per gram (n/g) of roots.
Population density was calculated as the mean number of nematodes in all samples in which a particular genus was present, expressed as n/dm3 of soil or n/g of roots.
The frequency of each nematode genus was computed as the proportion of samples in which the genus was present relative to the total number of samples (Boag, 1993). A bipartite diagram representing nematode genera across the regions was generated to visualize the distribution of nematode genera and cultivars in the surveyed areas.
Monthly climatic data of papaya orchards were retrieved for the 2018–2022 period (covering the survey interval) from NASA's POWER Project using the Nasapower R package (version 4.0.9; Sparks, 2018). Variables included relative humidity at 2 m above ground (RH2M), dew point temperature (T2MDEW), maximum air temperature (T2M_MAX), mean air temperature (T2M), and minimum air temperature (T2M_MIN). These data were extracted based on the geographic coordinates of each sampled orchards (61) across the nine papaya-producing administrative regions. Monthly averages of each climatic variable were correlated with the frequency and abundance of Meloidogyne and Rotylenchulus to determine the strength of association between climatic variables (averaged >5 years) and nematode infestation severity. All statistical analyses were conducted using R software (version 4.3) (R Core Team, 2021).
To assess the effect of cropping systems on nematode population densities, intercropping patterns (papaya monoculture, papaya–banana, papaya–eggplant) were observed in papaya orchards and documented through a pre-established survey form. Twenty-seven orchards were sampled per cropping system. Biplots and hierarchical clustering analyses were performed. To identify the significant differences of the mean number of nematodes among cropping systems, the Kruskal-Wallis test was followed by post-hoc Dunn's test at a significant level of P < 0.05. All statistical analyses were performed using R version 4.3 (R Core Team, 2021).
To enable species-level identification of the main nematodes associated with papaya in Burkina Faso, molecular characterization of 138 root or soil samples with Meloidogyne and Rotylenchulus was conducted at the Institut de Recherche pour le Développement (IRD) in Montpellier, France. Two DNA extraction methods were employed: a modified protocol from Holterman et al. (2006) for root nematodes, where β-mercaptoethanol was replaced by dithiothreitol (DTT), and the Qiagen DNeasy PowerSoil Pro Kit (Qiagen S.A.S., Courtaboeuf, France) for soil nematodes to overcome soil-derived PCR inhibitors (Roose-Amsaleg et al., 2001). Aliquots of 300 nematodes per sample were prepared in 2-ml Eppendorf tubes. One aliquot per sample was used to extract DNA. When necessary (no DNA or weak concentrations), extraction was repeated on additional aliquots. DNA extracted using the Holterman protocol was diluted 1:10, 1:20, and 1:50 in sterile distilled water before PCR amplification to avoid inhibition of PCR assays by soil or root chemicals and to permit repeating multiple PCR assays with different primer pairs. In general, PCR reactions were tested with a dilution of 1:50. Extracted DNA was stored at −20°C for future use.
The DNA was then amplified by PCR using universal primers for nematodes such as Nem-18S (Forward: CGCGAATRGCTCATTACAACAGC, Reverse: GGGCGGTATCTGATCGCC; Floyd et al., 2002) to check for nematode DNA presence. For Meloidogyne spp. detection, specific primers such as MIG (Forward: ACACAGGGGAAAGTTTGCCA, Reverse: GAGTAAGGCGAAGCATATCC; Zijlstra et al., 2000) and Mjav (Forward: 5GGTGCGCGATTGAACTGAGC, Reverse: CAGGCCCTTCAGTGGAACTATAC; Randig et al., 2002) were used. To test for Rotylenchulus reniformis type A and B presence, the D2A-F primer (ACAAGTACCGTGAGGGAAAGTTG) was tested as a forward primer with the type A reverse primers R1A-R (GAAAAGGCCTACCCAATGTG) or R2A-R (CCCGATACCATTTCCATACAA G) or the type B primer R1B-RCACAGARCCCRAGCAGCCA) as described in Van den Berg et al. (2016). All PCR reactions were performed in a thermocycler (MyCycler™ Thermal Cycler, Bio-Rad, Marnes-la-Coquette, France) in a reaction volume of 25 μl with different characteristics depending on the type of primers. In addition, the sequences of a subset of amplicons from PCR reactions were checked after Sanger sequencing at Genewiz (Steiβlingen, Germany). Amplicons were sequenced that were obtained with Nem primers (five soil samples) Rotylenchulus type A (three soil samples), MIG (eight root samples), and Mjav (three root samples). Sequencing was performed bidirectionally. Raw sequences were trimmed, aligned, and manually adjusted, and consensus sequences were generated in Geneious (version 8.1.8 created by Biomatters, available from http://www.geneious.com) (Kearse et al., 2012). The genetic relatedness of sequences obtained with other nematode sequences in the GenBank database was determined using the nucleotide basic local alignment search tool (BLASTN) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) with default parameters (Altschul et al., 1990).
Following the survey and sampling across the 9 papaya-producing regions, 10 nematode genera from 7 families were identified based on morphological criteria (Table 1). The most frequently encountered soil nematodes belonged to the genera Helicotylenchus (84.5%), Rotylenchulus (84.1%), Meloidogyne (68.6%), Scutellonema (52.7%), and Pratylenchus (33.3%). In terms of abundance in soil samples, the highest population densities were recorded for the genera Rotylenchulus (93,800 n/dm3 of soil), followed by Helicotylenchus (9,800 n/dm3 of soil), Meloidogyne (7,880 n/dm3 of soil), Scutellonema (3,100 n/dm3 of soil), and Pratylenchus (2,600 n/dm3 of soil). In root samples, Meloidogyne was observed with a frequency of 42%, reaching a maximum density of 1,573 n/g of roots.
Maximum density (n/dm3 of soil or n/g of roots) and frequency (%) of parasitic soil and root nematodes associated with papaya in Burkina Faso.
| Family | Genus | Density | Frequency |
|---|---|---|---|
| Soil nematodes | |||
| Hoplolaimidae | Rotylenchulus | 93,800 | 84.1 |
| Hoplolaimidae | Helicotylenchus | 9,800 | 84.5 |
| Heteroderidae | Meloidogyne | 7,880 | 68.6 |
| Hoplolaimidae | Scutellonema | 3,100 | 52.7 |
| Pratylenchidae | Pratylenchus | 2,600 | 33.3 |
| Telotylenchidae | Tylenchorhynchus | 400 | 14.0 |
| Criconematidae | Criconemoïdes | 60 | 12.1 |
| Longidoridae | Xiphinema | 40 | 4.3 |
| Trichodoridae | Trichodorus | 240 | 2.9 |
| Telotylenchidae | Telotylenchus | 40 | 0.48 |
| Root nematodes | |||
| Heteroderidae | Meloidogyne | 1,573 | 42.0 |
| Pratylenchidae | Pratylenchus | 219 | 13.0 |
| Hoplolaimidae | Rotylenchulus | 89 | 10.6 |
| Hoplolaimidae | Helicotylenchus | 25 | 8.2 |
| Hoplolaimidae | Scutellonema | 2 | 1.0 |
During the survey, two main papaya varieties were identified in the orchards: Solo, FBPA1, and a third group consisting of unknown varieties. A distribution map of the papaya varieties produced was developed based on the nine surveyed regions, to identify the type and percentage of varieties grown in papaya orchards across Burkina Faso (Table 2).
Distribution (%) of papaya varieties across nine production regions in Burkina Faso.
| Region | Papaya variety | ||
|---|---|---|---|
| Solo | FBPA1 | nd. | |
| Boucle du Mouhoun | 67 | 0 | 33 |
| Centre-South | 85 | 0 | 17 |
| Cascades | 25 | 62 | 12 |
| Centre | 67 | 17 | 17 |
| Plateau Central | 67 | 0 | 33 |
| Centre-East | 75 | 0 | 25 |
| South-West | 100 | 0 | 0 |
| Centre-West | 62 | 0 | 38 |
| High-Basins | 68 | 26 | 6 |
nd, not determined.
The Solo variety was thus produced in all nine surveyed regions, unlike the FBPA1 variety, which was only cultivated in the Centre, High-Basins, and Cascades regions. In addition to the identified varieties, several producers were unaware of the specific varieties grown in their orchards, a trend observed in all surveyed regions except the South-West, where only the Solo variety was produced.
The bipartite diagram (Figure 2) reveals that four nematode genera – Meloidogyne, Rotylenchulus, Helicotylenchus, Scutellonema – were common to all regions. In contrast, Telotylenchus and Trichodorus were specific to the High-Basins region. The greatest genus diversity was recorded in the High-Basins region, where all 10 identified genera were present. This was followed by the Cascades region, which harbored eight genera, differing from the High-Basins by the absence of Telotylenchus and Trichodorus. The lowest genus diversity was observed in the Centre (four genera) and Boucle du Mouhoun (five genera).
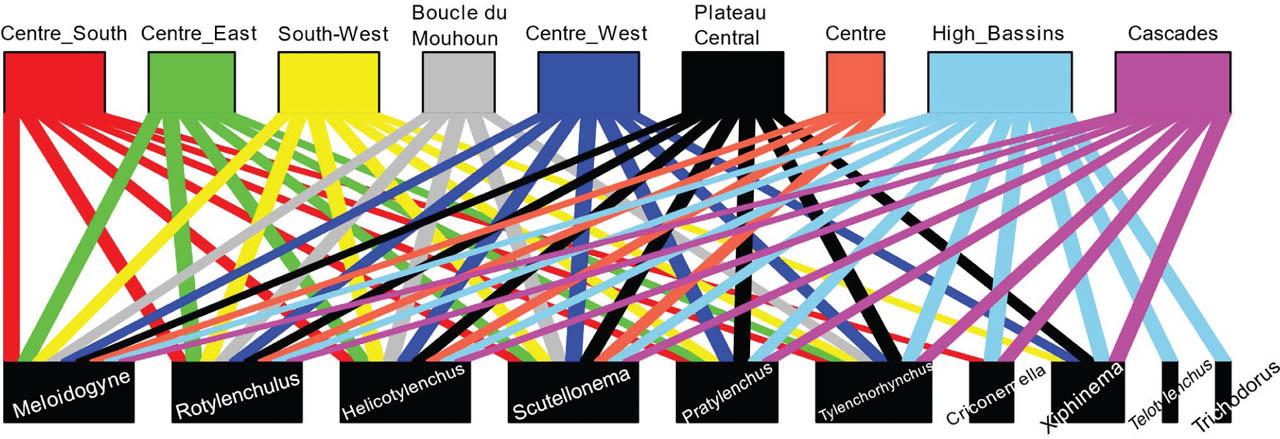
Bipartite diagram illustrating the distribution of nematode genera (down) by region (up) in Burkina Faso. A connecting line indicates the presence of a specific genus in a region. Boxes are sized according to the number of connections established.
Three papaya-based cropping systems were found to influence the population densities of nematodes in papaya orchards: monoculture of papaya, papaya–banana intercropping, and papaya–eggplant intercropping (Table 3; Figure 3). The study demonstrated that the papaya-based cropping systems significantly influence the population densities of parasitic nematodes associated with papaya.
Parasitic nematodes associated with papaya according to the cropping system (pure or associated to banana or eggplant).
| Nematode | Papaya | Papaya + Banana | Papaya + Eggplant | Global Average | P-value |
|---|---|---|---|---|---|
| Meloidogyne | 1,247 ± 1,205 | 525 ± 1,226 | 952 ± 1,798 | 908 | Papaya vs Banana (P = 7.17e−06) |
| Papaya vs Eggplant (P = 0.000758) | |||||
| Rotylenchulus | 11,767 ± 17,355 | 4,176 ± 8,470 | 5,250 ± 1,2217 | 7,064 | Papaya vs Banana (P = 3.81e−05) |
| Papaya vs Eggplant (P = 2.07e−05) | |||||
| Helicotylenchus | 2,391 ± 1,974 | 546 ± 2,126 | 1,552 ± 671 | 1,496 | Papaya vs Banana (P = 1.22e−06) |
| Papaya vs Eggplant (P = 0.02) | |||||
| Scutellonema | 161 ± 153 | 24 ± 51 | 239 ± 659 | 141 | Papaya vs Banana (P = 5.26e−08) |
| Papaya vs Eggplant (P = 0.005) | |||||
| Pratylenchus | 184 ± 487 | 10 ± 33 | 49 ± 132 | 81 | Papaya vs Banana (P = 2.85e−09) |
| Papaya vs Eggplant (P = 9.61e−07) | |||||
| Tylenchorhynchus | 33 ± 64 | 3 ± 11 | 30 ± 81 | 22 | Papaya vs Banana (P = 0.002) |
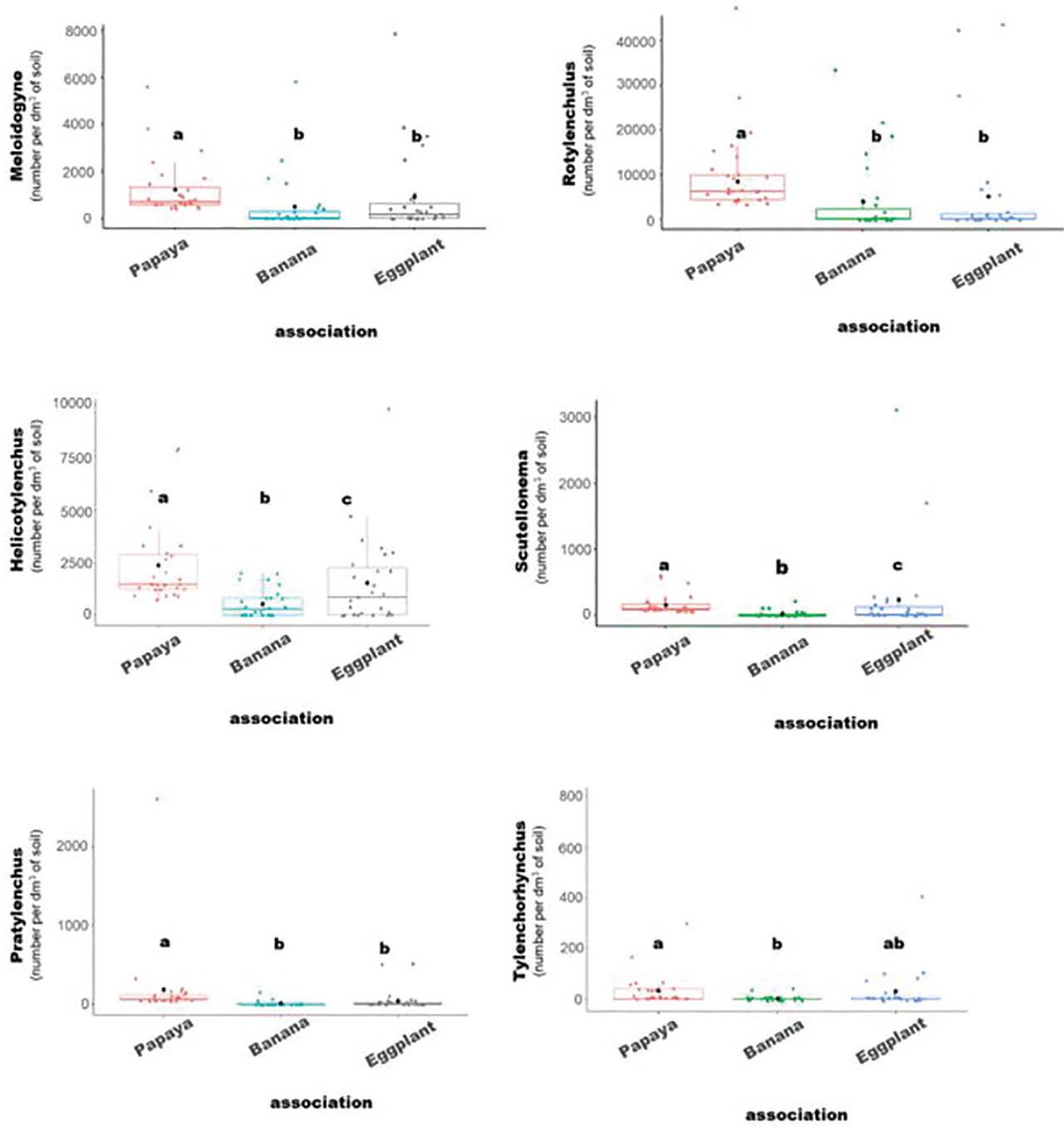
Box plots illustrating nematode population densities (Meloidogyne spp., Rotylenchulus spp., Helicotylenchus spp., Scutellonema spp., Pratylenchus spp., and Tylenchorhynchus spp.) according to the pure papaya cultivation system, papaya–banana associations, and papaya–eggplant associations. The study was conducted during a survey/collection of soil samples from 27 plots in papaya-based cropping systems to compare the impact of each system on nematode population densities. Different letters indicate statistically significant differences between groups when tested with the Kruskal–Wallis test, followed by post-hoc Dunn's test (P < 0.05).
Meloidogyne spp. exhibited an average density of 908 n/dm3 of soil, peaking in papaya monocultures (1,247 n/dm3 of soil). Papaya–eggplant and papaya–banana intercropping systems recorded lower densities, with highly significant differences between cropping systems. Rotylenchulus spp. showed the highest average density among all genera, with 7,064 n/dm3 of soil, and a marked predominance in papaya monocultures (11,767 n/dm3 of soil). This genus was significantly less abundant in the banana and eggplant intercropping systems. For Helicotylenchus spp., the overall average density was 1,496 n/dm3 of soil, again peaking in pure papaya orchards (2,391 n/dm3 of soil). Intercropped systems showed significantly lower densities. Although less abundant overall, Scutellonema spp. showed a clear preference for eggplant (238 n/dm3 of soil), with statistically significant differences among all cropping systems. Pratylenchus spp. also favored papaya, with an average density of 184 n/dm3 of soil, compared to only 49 n/dm3 in eggplant and 10 n/dm3 in banana systems, differences that were highly significant. Finally, Tylenchorhynchus spp. was generally less abundant (22 n/dm3 of soil on average), but mainly detected in papaya and eggplant fields, with significantly lower densities in banana associations (3 n/dm3 of soil).
Overall, papaya monoculture promoted the highest nematode densities across most genera, highlighting its key role in nematode proliferation. In contrast, intercropping, particularly with banana, appeared to exert a moderating effect on nematode populations.
The biplot (Figure 4A) illustrating the distribution of the main papaya-associated nematode genera in relation to papaya-based cropping systems revealed that the first two axes accounted for 83.4% and 16.6% of the total variance, respectively. This plot showed a strong positive correlation between the papaya–eggplant intercropping system and the genera Scutellonema spp. and Tylenchorhynchus spp. Conversely, papaya monoculture was associated with high population densities of Rotylenchulus spp., Pratylenchus spp., Helicotylenchus spp., and Meloidogyne spp.
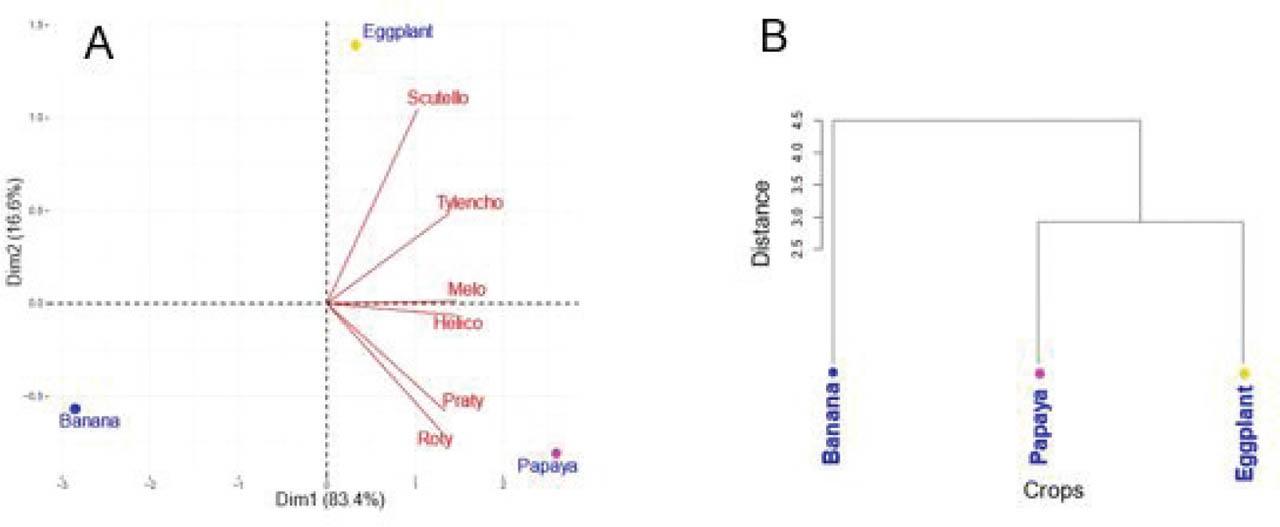
Distribution of parasitic nematodes associated with papaya according to cropping systems (pure or associated with banana or eggplant): (A) Biplot of PPN in interaction with cropping systems. (B) Hierarchical cluster analysis showing similarities between cropping systems based on nematode population densities. PPN, plant-parasitic nematode.
According to the hierarchical cluster analysis (Figure 4B), the cropping systems involving papaya and eggplant shared similar nematode population structures, while banana intercropping formed a distinct cluster, indicating a different nematode community profile.
An overview of the distribution of Meloidogyne spp. in the 61 orchards surveyed is presented in Figure 5. Nematode prevalence (Figures 5A,C) ranged from 0% to 100%, while abundances (Figures 5B,D) ranged from 0 nematodes/dm3 to 3,940 nematodes/dm3 of soil (Table S1 in Supplementary Materials). The correlation between frequency and abundance of Meloidogyne was weak (r = 0.474; P < 0,001), indicating that the abundance of Meloidogyne does not depend on its frequency.
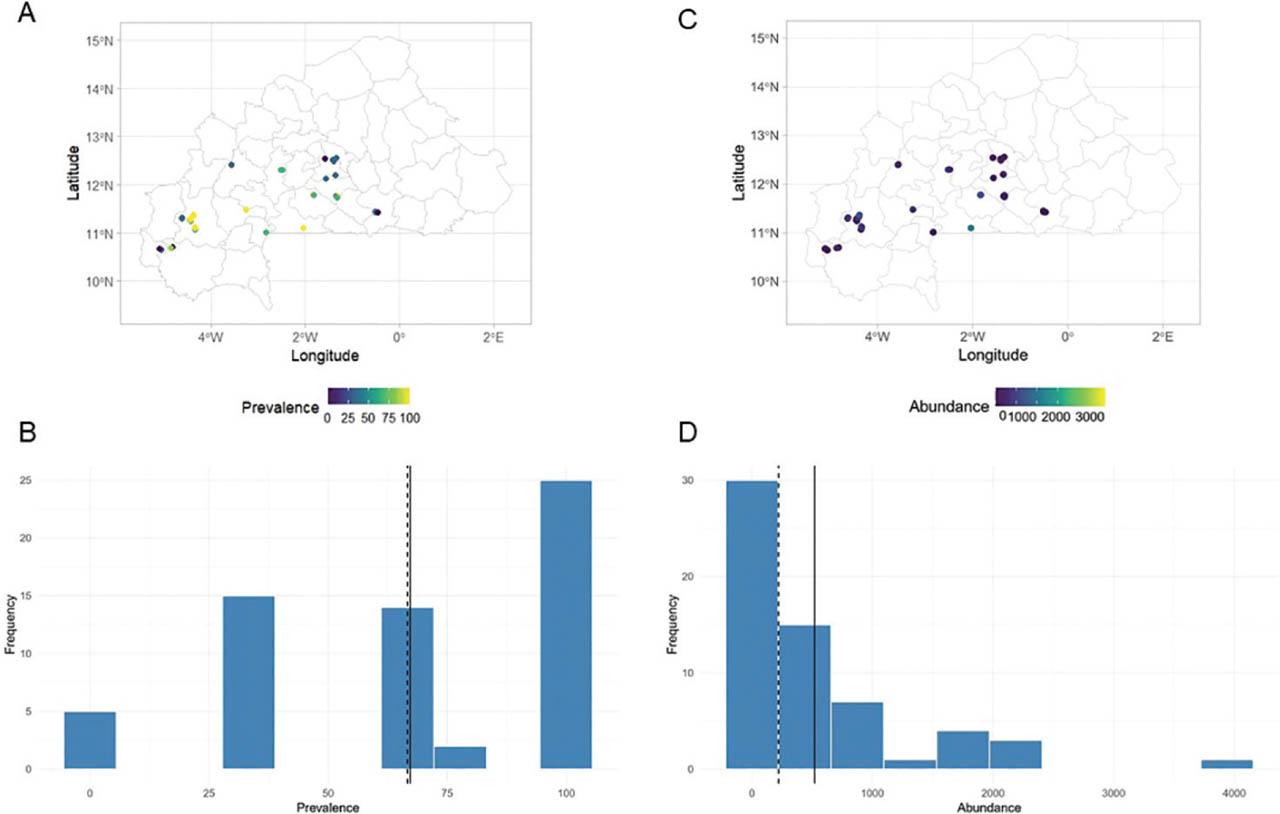
Distribution maps of the prevalence (A,B) and abundance (C,D) of Meloidogyne spp. root samples of 61 papaya orchards across Burkina Faso.
High positive correlations were observed between the average air temperature at 2 m above ground level (T2M), the maximum air temperature at 2 m above ground level (T2M_MAX) and the frequency of Meloidogyne spp. with coefficients of 0.75. Conversely, significant negative correlations were observed between precipitation (PREC), relative humidity at 2 meters above ground level (RH2M) and dew point temperature at 2 m above ground level (T2MDEW) with respective coefficients of −0.74, −0.81 and −0.77 (Figure S1A in Supplementary Materials).
The abundance of Meloidogyne sp. was strongly positively correlated with RH2M and T2MDEW, with coefficients of 0.63 and 0.70. In contrast, T2M_MIN, T2M_MAX and T2M were negatively correlated with the abundance of Meloidogyne spp. in papaya orchards with respective coefficients of −0.84, −0.55 and −0.49 (Figure S1B in Supplementary Materials).
Figure 6 presents the distribution of Rotylenchulus spp. across the 61 surveyed orchards. Nematode prevalence (Figures 6A,C) ranged from 0% to 100% while abundances (Figures 6B,D) varied between 0 n/dm3 and 35,100 n/dm3 of soil (Table S2 in Supplementary Materials).
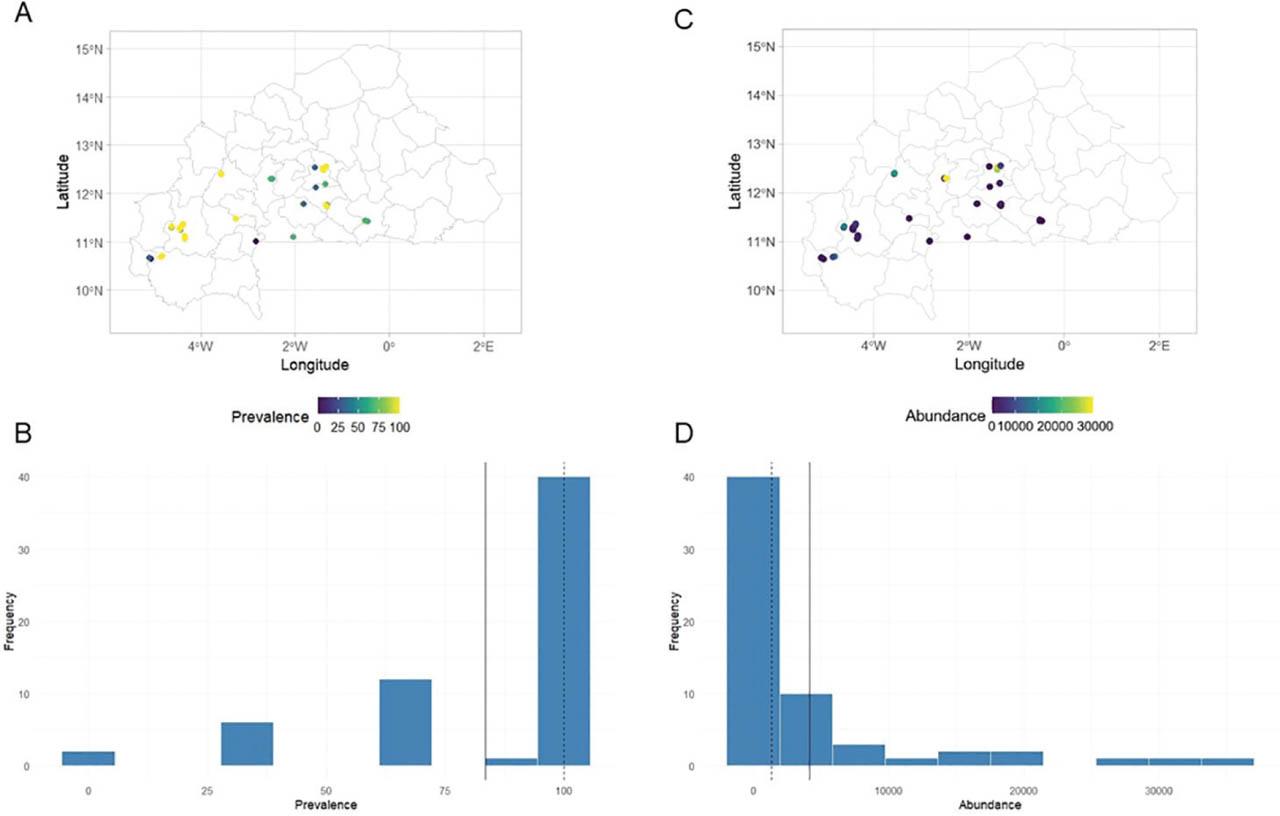
Distribution maps of the frequency (A,B) and abundance (C,D) of Rotylenchulus spp. in soil samples of 61 papaya orchards across Burkina Faso.
Strong positive correlations were found between the average air temperature at 2 m above ground level (T2M), the maximum air temperature at 2 meters above ground level (T2M_MAX), and the frequency of Rotylenchulus spp., with respective coefficients of 0.63 and 0.60. In contrast, strong negative correlations were observed between PREC, relative humidity at 2 meters above ground level (RH2M), and the frequency of Rotylenchulus spp. with respective coefficients of −0.81 and −0.53 (Figure S2A in Supplementary Materials).
The average air temperature at 2 m above ground level (T2M) and the maximum air temperature at 2 meters above ground level (T2M_MAX) were positively correlated with the abundance of Rotylenchulus spp. The coefficients were 0.78 and 0.86, respectively. However, negative correlations were observed between abundance and PREC, relative humidity 2 meters above ground level (RH2M), dew point temperature at 2 m above ground level (T2MDEW), and minimum air temperature at 2 m above ground level (T2M_MIN), with coefficients of −0.80, −0.93, −0.91, and −0.64 (Figure S2B in Supplementary Materials).
Molecular characterization of nematodes was performed on composite samples collected in 2021 from papaya roots or soil associated with papaya to identify the main species of Meloidogyne and Rotylenchulus. DNA was successfully extracted from 138 samples containing one or both genera, and further amplified by PCR using several primer pairs targeting different nematode genomic regions. Amplified bands were sequenced for a subset (3–8) of randomly chosen isolates to verify their homology with the corresponding nematode species targeted by primers (data not shown). Some amplicons obtained with Nem primers contained mixed sequences from several nematodes.
Using the universal nematode primers Nem-18S, 132 out of the 138 samples produced an amplified fragment of 900 bp, indicating the presence of nematode DNA. Among these 132 samples, 129 tested positive with the Mjav-F/Mjav-R primer pair, specific to M. javanica (670 bp amplicon). Additionally, primers specific to the MIG group (M. incognita, M. javanica, M. arenaria) amplified DNA in 114 samples, yielding a 500 bp fragment. Some discrepancies between results of amplification with Mja or MIG primers also occurred, suggesting that other Meloidogyne species may be present (for samples MIG+; Mjav−) and that MIG primers do not amplify all M. javanica isolates (for samples MIG−; Mjav+).
From 80 soil samples where Rotylenchulus had been observed, DNA from R. reniformis was amplified in 77 samples using the type A-specific primers (D2AF/R1A-R or D2AF/R2A-R), resulting in amplicons of 142 bp and 320 bp, respectively. No amplification could be observed with the type B primers (D2AF/R1B-R/179 pb), suggesting that only R. reniformis type A may be present in papaya orchards surveyed.
This study assessed the diversity and distribution of the major PPNs associated with papaya in nine major production regions of Burkina Faso. Using combined morphological and molecular approaches, 10 nematode genera, belonging to 7 different families were identified. In addition, molecular markers allowed characterization of M. javanica as in a large number of samples, and R. reniformis (type A) as the Rotylenchulus species present in papaya root and soil samples. Other Meloidogyne species may be present in some MIG-positive samples since MIG primers can also detect M. incognita and M. arenaria (Zijlstra et al., 2000). The most frequently encountered genera were Helicotylenchus, Rotylenchulus, Meloidogyne, Scutellonema, and Pratylenchus. These findings align with reports from other papaya-growing regions such as Latin America and Southeast Asia (Da Silva et al., 2007; Martínez Gallardo et al., 2014; Indarti and Taryono, 2023), suggesting a shared nematode fauna across tropical climates.
Rotylenchulus reniformis was the most abundant and widely distributed species, with densities reaching 93,800 n/dm3 of soil. Its prevalence confirms its adaptability to diverse environments and its status as a major constraint in tropical horticulture (El-Borai and Duncan, 2005; Martínez Gallardo et al., 2014). M. javanica, the second most dominant species, reinforces earlier findings on papaya susceptibility to root-knot nematodes, which are known to cause significant yield losses (Peraza Padilla, 2021). Several other authors have reported the presence of these two species in papaya orchards around the world (Bridge et al., 1996; El-Borai and Duncan, 2005; Dias-Arieira et al., 2008; Perera et al., 2008; Plena et al., 2022; Baldan et al., 2024; Wanjala et al., 2025). According to Peraza Padilla (2021), M. javanica is one of the four most commonly identified root-knot nematode species on papaya in Brazil and Colombia, and it was recently reported in Kenya (Wanjala et al., 2025). M. javanica is the predominant species of root-knot nematode in regions with well-defined dry seasons (Eisenback et al., 1981), as is the case in Burkina Faso. Molecular confirmation of M. javanica emphasizes the importance of integrating molecular diagnostics for accurate species-level identification in nematode surveys.
Differences in nematode community composition across regions are likely influenced by multiple interacting factors, including climate, soil texture, cropping history, and management practices. Regional differences in rainfall (600–1,200 mm), temperature (27–28°C), and humidity may shape nematode population dynamics and spatial patterns (MECV/PANA, 2007). This agrees with previous studies demonstrating the role of edaphic and climatic factors in structuring nematode communities (Norton, 1979; Yeates et al., 1993; Mateille et al., 2014).
Cropping systems significantly affected nematode presence and abundance. Papaya monocultures tended to harbor higher densities of Meloidogyne and Rotylenchulus compared to intercropped systems, suggesting that continuous cropping of a single host may amplify nematode pressures. Intercropping with banana or eggplant, both known hosts of several PPNs, did not mitigate nematode infestation, possibly due to overlapping host ranges (Netscher and Sikora, 1990; Daneel and De Waele, 2017). Moreover, the predominance of Scutellonema, Pratylenchus, and Helicotylenchus in monocultures suggests that papaya supports a broad spectrum of PPNs, underlining the need for crop rotation or soil health management practices to suppress nematode buildup.
Climatic variables were also key drivers of nematode population dynamics. Relative humidity and dew point were positively correlated with Meloidogyne abundance, reflecting its preference for moist conditions conducive to hatching and root penetration. Rotylenchulus populations were more responsive to average air temperature and maximum air temperature, consistent with their ability to thrive in warmer climates. These findings support earlier work on climate-nematode interactions (Ankrom et al., 2020; Dutta and Phani, 2023) and suggest that climate change may exacerbate PPN problems in the region.
Overall, the results reveal a rich and diverse PPN community associated with papaya in Burkina Faso, with important implications for disease management. Integrated nematode control strategies – including resistant cultivars, improved cropping systems, and soil health restoration – will be essential for sustainable papaya production in the face of nematode pressure and climatic variability.