
Globodera pallida (Stone) Behrens, the pale cyst nematode, is a major pest in potato worldwide. It is estimated to cause over 80% yield loss in highly infested fields (Contina et al., 2019). G. pallida originated from the Andes region of South America, but has spread to most major potato growing regions of the world. G. pallida was first found in fields in southeastern Idaho in 2006 and is regulated by USDA-APHIS (Hafez et al., 2007; Skantar et al., 2007; USDA-APHIS, 2009). The regulated area of infested and associated fields encompasses 2,611 ha in Bonneville and Bingham counties, consisting of about 1% of Idaho potato production acreage (USDA-APHIS, 2024). Idaho is the leading U.S. producer of potato, producing about 6,870,255 MT worth US$1.26 billion in 2024 (USDA-NASS, 2025a, 2025b). Potato cannot be planted in G. pallida infested acreage until regulatory steps have been satisfied (USDA-APHIS, 2024). Even with a low percentage of potato acreage out of production from G. pallida infestation, the Idaho potato industry loses US$30 million annually due to G. pallida, offset by only US$5 million from production of other crops (Koirala et al., 2020).
The primary control measures for G. pallida eradication in Idaho are stringent phytosanitary measures and fumigation with 1,3-dichloropropene (USDA-APHIS, 2024). Other control strategies such as trap crops and crop rotation may be of use to support G. pallida eradication efforts. Crop rotation is a common strategy for controlling pests and diseases in agricultural crops because growers can rotate to non-hosts, fallow, or resistant varieties to disrupt the pest lifecycle in an effort to reduce the pest population below an economically damaging level. Effective crop rotation is determined by several factors, including the target pest's lifecycle and host range, length of rotation, availability of resistant varieties and non-hosts, and the production feasibility and market for the rotational crop. Because G. pallida is highly specialized with a narrow host range, rotation to a non-host could be expected to be effective in reducing population densities. However, G. pallida requires specific hatching factors to hatch and can survive in a dormant state in the absence of a host for decades (Turner, 1996; Masler and Perry, 2018). Thus, rotations to a non-host or fallow are ineffective in controlling this nematode.
Globodera pallida persists in soil as protective cysts containing hundreds of eggs. The eggs contain second-stage juveniles (J2) which require a chemical hatching factor, typically produced by its primary host, potato, or other closely related solanaceous species (Masler and Perry, 2018; Ngala et al., 2021). Rotation to fallow or a non-host does not stimulate hatch of G. pallida and only results in about 20%–40% annual decline in the population due to natural attrition resulting from environmental conditions, microbes, and some spontaneous hatch (Storey, 1984; Brodie, 1996; Turner, 1996; Atkinson et al., 2001; López-Lima et al., 2013). Dormant, unhatched juveniles from field populations also begin to lose their lipid reserves, which reduces their ability to infect, but a 50% loss in lipid reserves can take at least 7.5 years (Storey, 1984; Atkinson et al., 2001). Efficient crop rotation to control G. pallida would incorporate resistant potato varieties or trap crops that induce hatch but prevent or limit reproduction of the nematode to decrease overall population densities.
Various crop rotation strategies have been investigated for Globodera spp. in different regions of the world. The tobacco cyst nematode, Globodera tabacum tabacum (Lownsbery & Lownsbery) is only reduced by 30%–40% after a non-host crop, but a resistant tobacco variety or tomato trap crop was found to reduce nematode population densities by as much as 96% (LaMondia, 1996). In the U.K., Whitehead et al. (1980) saw the golden nematode, Globodera rostochiensis (Wollenweber) Behrens controlled in rotation with nematicide before susceptible or resistant potato followed by 2 years of non-host. Tiilikkala (1991) found that in Finland, resistant potato crops reduced G. rostochiensis populations by 70%–80% each year, while 3 years of susceptible potato monoculture increased the population from 0.1 larvae/g soil to 265 larvae/g soil. In Canada, Bélair et al. (2016) observed that 3 years of a resistant potato reduced G. rostochiensis viable eggs to zero in some plots with an overall population density reduction of 95%. Additionally, 1 year of resistant potato reduced G. rostochiensis populations to 73%–86%, while Solanum sisymbriifolium Lam. as a trap crop reduced populations to 59%–66%, compared with only 28%–35% reduction by natural attrition during a non-host crop (Bélair et al., 2016).
Reduction of G. pallida and G. rostochiensis populations is limited with short rotations to non-host or fallow (Storey, 1984; Brodie, 1996; Turner, 1996; Atkinson et al., 2001; López-Lima et al., 2013). This was also demonstrated by López-Lima et al. (2013), who found that the G. rostochiensis infestation level only decreased by 30% in a rotation to either fallow or non-host crops, pea (Pisum sativum L.) and faba bean (Vicia faba L.). Long rotations to non-host or fallow are required to reduce the population densities below damaging levels. In the U.K., Whitehead (1995) observed that after 4 years of non-host barley, G. pallida and G. rostochiensis egg densities had decreased by as much as 86% and 71%, respectively. Long rotations of non-host barley for 4–6 years were found to reduce G. rostochiensis populations from 20 eggs/g soil to about 0.6–1.6 eggs/g soil, which prevented yield loss in subsequent susceptible potato (Whitehead et al., 1991). However, despite these years of a non-host crop, planting susceptible potato caused up to a 50-fold increase in the population (Whitehead et al., 1991). Regardless of natural attrition during several seasons of fallow or non-hosts, there can be a rapid resurgence of the population when susceptible potato is planted again, because some viable eggs survive season to season (Brodie, 1996; Turner, 1996; Devine et al., 1999; Ryan and Devine, 2005). Increase of G. pallida on susceptible potato depends on the potato variety and initial population density, but can be dramatic even in just one season (Whitehead et al., 1987, 1991). Ultimately, long rotations of non-hosts would not be practical for Idaho where rotations are typically 3 years and the goal is to eradicate G. pallida.
Susceptible potato has been investigated as a trap crop in rotation, but is not practical as it must be terminated before the nematode can reproduce, at the expense of the potato yield (Whitehead, 1977, 1992; Scholte, 2000a; Bélair et al., 2016). Susceptible potato was found to reduce G. pallida populations up to 93% when terminated after 8 weeks of growth (Whitehead, 1992). In long-term field studies, Scholte (2000a) saw that a susceptible or resistant potato trap crop grown for 8 weeks, then rotated to resistant potato or fallow, caused 92%–97% reduction in G. pallida populations. Compared with a season of susceptible potato, they also saw a 95%–97% reduction in G. pallida population density when a potato trap crop was planted before rotating to a susceptible potato (Scholte, 2000a). However, with a resistant potato alone, Halford et al. (1999) saw a 95% reduction in G. pallida populations while achieving a profitable yield.
In New York, G. rostochiensis is controlled with a crop rotation that incorporates resistant potato. Potato varieties with the H1 resistance gene provide resistance to pathotype Ro1, the primary pathotype of the New York population (Brodie and Mai, 1989; Limantseva et al., 2014; Whitworth et al., 2018). Numerous studies were performed to develop the most effective rotation to control G. rostochiensis in New York. LaMondia and Brodie (1986b) saw that a resistant potato grown for 4 weeks reduced G. rostochiensis to 50%–60% versus a 15%–30% reduction when infested fields remained fallow. Depending on the initial population density, rotation to a season of a completely resistant potato variety can reduce population densities to 75%–95% (LaMondia and Brodie, 1986a; Brodie, 1996). In comparison, a susceptible potato can increase G. rostochiensis densities by as much as 25- to 50-fold (Jones, 1970; LaMondia and Brodie, 1986a). However, when G. rostochiensis population densities were very low at an initial population of 0.01 eggs/cm3 soil or less, Brodie (1996) reported no detectible population increase on a susceptible potato variety (Jones, 1970; Brodie, 1996). Studies also showed that the spread of G. rostochiensis is limited at populations of below 0.2 eggs/cm3 soil or less (Brodie, 1996). G. rostochiensis was therefore found to be suppressed below 0.2 eggs/cm3 soil by a rotation of 2 years of resistant potato, then 1 year of a non-host, followed by 1 year of susceptible potato (Brodie, 1996). Based on these research findings, this 4 years crop rotation is employed to successfully control and contain G. rostochiensis in New York (Brodie and Mai, 1989; Brodie, 1996; Dandurand et al., 2019b).
While there are European potato varieties with resistance to G. pallida, a resistant variety for commercial use in Idaho is not yet available (Whitworth et al., 2018). Most Idaho potatoes are grown for processing into french-fries, for which white-flesh oblong russet-type potatoes are preferred. Efforts to breed a G. pallida-resistant russet potato for Idaho are underway, so for the purposes of this study, a European variety was used to determine the effect of a resistant potato in an Idaho crop rotation to reduce G. pallida. This study uses ‘Innovator’, a yellow-flesh russet potato developed in the Netherlands with high resistance to G. pallida pathotype Pa2/3 (Buckley, 2015; Varypatakis et al., 2019), which is the same pathotype as the Idaho population (Blok and Phillips, 2012).
Another valuable tool for use in rotation to reduce G. pallida may be the trap crop S. sisymbriifolium. S. sisymbriifolium is a non-host of G. pallida that induces significant egg hatch, thereby dramatically reducing the population of G. pallida (Scholte, 2000b; Scholte and Vos, 2000; Timmermans et al., 2006). Multiple studies have found that S. sisymbriifolium in rotation with susceptible potato varieties reduces G. pallida by over 90% (Dandurand and Knudsen, 2016; Dandurand et al., 2019a; Mhatre et al., 2021). While S. sisymbriifolium has proven to be a successful trap crop for G. pallida, its use has not been widely adopted because of limited access to seed. Nevertheless, S. sisymbriifolium remains an effective trap crop for G. pallida and its efficacy in rotation with a resistant potato should be determined.
The goal of this study is to determine the potential for a resistant potato variety and the trap crop S. sisymbriifolium to reduce G. pallida populations when in rotation with susceptible potato. The impact of 3-years crop rotations on G. pallida were evaluated under Idaho field conditions. These rotations incorporated the resistant potato variety ‘Innovator’ and the trap crop S. sisymbriifolium. The rotation sequences were evaluated by assessing G. pallida initial population reduction and reproduction on potato after each growing season. To better understand how these crops may reduce G. pallida egg densities when in rotation, hatching assays were conducted in vitro to compare hatch rates caused by ‘Innovator’ and S. sisymbriifolium. The results from this study may contribute to development of a crop rotation plan for controlling G. pallida in Idaho.
Globodera pallida was originally obtained from infested fields near Shelley, ID and identified with morphological and molecular methods (Skantar et al., 2007). G. pallida used in this study was reared on susceptible potato ‘Désirée’ or ‘Russet Burbank’ for 16 wk in the greenhouse at 18°C with 16-hr light/8-hr dark photoperiod and 60% relative humidity. Soil and roots were dried for 2 wk, then extracted with an elutriator to recover cysts (USDA-APHIS, 2009). Following extraction, cysts were dried, separated from debris, and stored at 4°C until use.
The hatching effect of ‘Innovator’ and S. sisymbriifolium was compared in vitro with that of susceptible potatoes ‘Désirée’ and ‘Russet Burbank’, and to a bare soil-negative control using diffusate applied to G. pallida eggs. For diffusate production, four replicate pots per treatment were grown in randomized complete block design in the greenhouse in 15-cm-diam terracotta pots containing 1,200 g soil. Soil was a 2:1 ratio of Lane Mountain 20/30 industrial silica sand (Valley, WA) to Mission-series silt loam soil (UI-SOAC, Sandpoint, ID) that had been dried and sieved through a 5-mm mesh (Dandurand and Knudsen, 2016). Mixed sieved soil composition was 76% sand, 14% silt, and 10% clay (1% organic matter; pH 5.5). Prior to use, soil was autoclaved twice at 121°C for 90 min with 48 hr between cycles. Pots were autoclaved once at 121°C for 90 min. Greenhouse conditions were 18°C with 16-hr light to 8-hr dark photoperiod and 60% relative humidity. The potato varieties ‘Désirée’, ‘Russet Burbank’, and ‘Innovator’ were grown in tissue culture using standard media (Murashige and Skoog, 1962) for 4 wk before transplanting into the experiments. S. sisymbriifolium was germinated and grown for 4 wk in pro-mix potting soil (Premier Horticulture, Quakertown PA) before transplanting into pots. Two plants were planted per pot. Unplanted bare soil pots were also included as a treatment. Diffusate was collected from each pot at 4-wk and at 6-wk of growth following transplanting into soil. Diffusate from replicate pots was maintained in distinct replicates. Diffusate was collected by the soil leaching method modified from Widdowson and Wiltshire (1958) and then filtered, as described by Hickman and Dandurand (2023). The resulting diffusate samples were then frozen at −20° until use.
Hatching assays were performed within 1 mon of collecting the diffusate. The assays utilized cysts of the Idaho G. pallida population as described above. Cysts were first surface sterilized in 0.3% hypochlorous bleach for 5 min before being thoroughly rinsed five times in sterile deionized water (Nour et al., 2003). Cysts were then placed into 96- well plates with one cyst and 100 μl of 100 μg/ml gentamicin sulfate (VWR®, Sanborn, NY) solution per well. Cysts were hydrated for 48 hr at 18°C. Following hydration, cysts were crushed to release all eggs. Eggs, as well as second stage juveniles (J2) that were released from the egg, were counted using an inverted microscope (Leica DMi1). Diffusates were thawed to room temperature. Each diffusate replicate was applied to four cysts and 100 μl of diffusate was applied. The plates were incubated at 18°C for 2 wk and then the final hatched second stage juveniles (J2) were counted. The hatch percentage was calculated as (Final hatched J2 − Initial J2)/Initial eggs × 100. The root diffusate experiment and hatching assays were independently repeated.
Two field rotation experiments were each conducted over 3 yr in microplots designed for G. pallida containment (Fig. 1) at a field site in a G. pallida-infested field in Shelley, ID. Microplot setup has been previously described by Dandurand et al. (2019a) with the following modifications. Microplots were made up of three 19-l buckets (ULINE, Pleasant Prairie, WI), consisting of an upper bucket, a middle half bucket, and a lower bucket (Fig. 1A). The upper bucket had six 1-cm-diam holes drilled into the bottom for drainage (Fig. 1B). To prevent escape of cysts, the drainage holes were covered by a nylon mesh with 250-μm pores (McMaster Carr, Elmhurst, IL) caulked into the bottom of the bucket with silicone. The lower bucket was left intact to function as containment for water that drained from the upper bucket. A third bucket was cut into half and the upper portion was used to create space between the upper bucket and the lower bucket for drainage water collection. Water or soil collected in the lower bucket was pumped out through a 10-μm pore filter bag (Pentek, Golden Valley, MN) to remove any escaped G. pallida. Microplots were spaced 1-m apart and placed in the ground so that the lower bucket was level with the soil surface. The upper bucket of the microplot was filled with a layer of river rock approximately 7.6 cm in depth (Fig. 1C) followed by Bock-series loam field soil (40% sand, 40% silt, 20% clay; 3.7% organic matter; pH 8.2) 25 cm in depth (Fig. 1E). As an additional G. pallida containment precaution, the soil surface of the upper bucket was covered with a layer of landscape fabric (Vigoro, Portland, TX) on top of a layer of nylon mesh with a 250-μm pore size (Fig. 1F). Mesh and landscape fabric were secured over the soil surface with 15-cm galvanized steel yard staples.

Field microplot design for G. pallida containment. (A) Microplot design of three 19-l buckets consisting of an upper bucket, a middle half bucket, and a lower bucket. (B) The upper bucket had six 1-cm-diam holes drilled into the bottom for drainage, covered by 250-μm pore nylon mesh caulked into the bottom of the bucket with silicone. (C) The upper bucket of the microplot was filled with a layer of river rock approximately 7.6 cm in depth. (D) G. pallida cyst bags were attached to stakes placed in the microplots so that cyst bags would be in the anticipated root zone at a depth of approximately 15 cm below the soil surface. (E) Field soil was added to the upper bucket, approximately 25 cm in total depth. (F) The soil surface of the upper bucket covered with a layer of landscape fabric on top of a layer of 250-μm pore nylon mesh and 4-wk-old S. sisymbriifolium seedlings planted into the microplot.
Microplots were inoculated with G. pallida cysts sealed within a nylon mesh bag with 250-μm pores approximately 5 cm × 3.8 cm and double-sealed with a table top impulse sealer (ULINE, Pleasant Prairie, WI). For easy recovery, cyst bags were tied to 0.6 m of a 22.7 kg monofilament fishing line (Zebco Holdings Inc., Tulsa, OK) with a Palomar fishing knot. The fishing line was then secured to 30-cm stakes with duct tape. The stakes were placed in the microplots so that the cyst bags would be in the anticipated root zone at a depth of approximately 15 cm below the soil surface (Fig. 1D). In the first year of rotation, microplots were inoculated at 7.5 eggs/g soil. To achieve 7.5 eggs/g soil, 300 cysts per microplot were divided into 15 cyst bags with 20 cysts per cyst bag. Cyst bags were distributed throughout the plot on five stakes with a single cyst bag and two stakes with five cyst bags. Holes were cut in the layers of mesh and landscape fabric on the microplot soil surface to allow for plant emergence and cyst bag stakes.
Solanum sisymbriifolium seeds were produced at the USDA Agricultural Research Service in Prosser, WA. Seeds were germinated in pro-mix potting soil and grown for 3 wk in the greenhouse. Seedlings were acclimated outside in the shade 1 wk prior to transplantation in the field microplots. Four seedlings were transplanted per microplot. Potatoes ‘Innovator’ and ‘Russet Burbank’ were planted as mini tubers received from the University of Idaho Seed Potato Germplasm lab in Moscow, ID. Four mini tubers were planted per microplot. Barley variety ‘GemCraft’ (Hu et al., 2024) was directly seeded into the microplots at 2.5-cm planting depth. Before planting each season, the soil was tilled using a hand trowel. Osmocote Smart-Release Plant Food Plus Outdoor and Indoor slow-release fertilizer (15-9-12 NPK) (The Scotts Company, Marysville, OH) was applied on the plants at a rate of 50 g/plot. Microplots were watered daily to maintain soil moisture. Jack's Classic All Purpose 20–20–20 water soluble fertilizer (JR Peters Inc., Allentown, PA) was applied weekly at a rate of 0.5 g/l of water. Microplots were grown for 12 wk each season.
Six different rotation sequences with five replicates for each sequence arranged in randomized complete block design were grown in Idaho field conditions (Table 1). In the “Results” and “Discussion” sections, rotation crops are abbreviated and the sequences are hyphenated (Table 1). The susceptible potato ‘Russet Burbank’ is abbreviated as ‘RB’, the resistant potato ‘Innovator’ is abbreviated as ‘INN’, and Solanum sisymbriifolium is abbreviated as ‘S. sisym.’ Two trials of this experiment were performed. Rotation trial 1 was started in 2020 and was completed in 2022. Rotation trial 2 was started in 2021 and was completed in 2023. Planting took place in the beginning of June each year. Plants were grown for 12 wk following tuber sprout emergence. The growing season ended in mid-September each year. Year 1 of the rotation was the resistant potato ‘Innovator’ or S. sisymbriifolium. Year 2 of the rotation was ‘Innovator’, S. sisymbriifolium, or the non-host barley. In year 3 of the rotation, all microplots were planted with susceptible potato ‘Russet Burbank’. At the end of each growing season, plants were terminated by being cut to the soil surface and the roots were removed. Tubers from each microplot with potato were also removed. Soil and cyst bags were sampled. Microplots were dried out for 2 wk, then sealed and transported to a storage unit where they were stored until the next growing season.
Rotation sequences with corresponding abbreviations.
| Year 1 | Year 2 | Year 3 | Rotation sequence abbreviationa |
|---|---|---|---|
| ‘Innovator’ resistant potato | ‘Innovator’ resistant potato | ‘Russet Burbank’ susceptible potato | INN-INN-RB |
| ‘Innovator’ resistant potato | S. sisymbriifolium | ‘Russet Burbank’ susceptible potato | INN-S. sisym.-RB |
| ‘Innovator’ resistant potato | Barley (non-host) | ‘Russet Burbank’ susceptible potato | INN-Barley-RB |
| S. sisymbriifolium | ‘Innovator’ resistant potato | ‘Russet Burbank’ susceptible potato | S. sisym.-INN-RB |
| S. sisymbriifolium | S. sisymbriifolium | ‘Russet Burbank’ susceptible potato | S. sisym.-S. sisym.-RB |
| S. sisymbriifolium | Barley (non-host) | ‘Russet Burbank’ susceptible potato | S. sisym.-Barley-RB |
In the rotation sequences, susceptible potato ‘Russet Burbank’ is abbreviated as ‘RB’, resistant potato ‘Innovator’ is abbreviated as ‘INN’, and Solanum sisymbriifolium is abbreviated as ‘S. sisym’.
At the beginning and at the end of each growing season, one stake with a single cyst bag was removed from each microplot to sample the original G. pallida population used to infest soil. These cysts were used in viability and hatch assays to assess the remaining encysted eggs of the initial population used to infest the microplots. Cysts were first surface sterilized in 0.3% hypochlorous bleach for 5 min followed by five thorough rinses with sterile deionized water (Nour et al., 2003). Five cysts per replicate were used for the viability assay. Eight cysts per replicate were used for the hatching assay.
For the viability assay, cysts were placed into a 96-well plate (VWR® tissue culture plates, Radnor, PA) with one cyst and 135 μl sterile deionized water per well. Cysts were gently crushed with forceps to release the eggs. The eggs were counted using an inverted microscope (Leica DMi1). The acridine orange method was used to determine egg viability (Pillai and Dandurand, 2019). An amount of 15 μl of 100 μl/ml acridine orange stain (Life Technologies Corporation, Eugene, OR) was applied per well. Plates were incubated at 18°C in the dark for 4 hr. The stain was washed and fluorescing non-viable eggs were counted using an inverted fluorescent microscope (Leica DMi8, Leica Microsystems, Wetzlar, Germany). The percentage of viability was calculated as (viable eggs)/total eggs × 100.
Hatching assays were conducted by applying root diffusate to the remaining encysted eggs; G. pallida eggs for the cyst samples were collected at the beginning and end of each growing season. Four cysts per replicate were incubated with potato root diffusate (PRD) and four cysts per replicate were incubated with bare soil diffusate. Diffusate was collected from 4-wk-old ‘Désirée’ potato or bare soil by a modified soil leaching method adapted from Widdowson and Wiltshire (1958), as described above. Diffusates were kept frozen at −20°C and were less than 2 mon old at the time of use. Hatching assays were performed as previously described.
At the end of the first- and second-year growing seasons, 500-g soil samples were taken from each microplot. Soil sampling was done by taking five soil cores uniformly distributed throughout each plot. Soil cores were combined and homogenized, then 500 g soil was weighed out and placed into a labeled paper sample bag. The remaining soil was returned to the microplot. Soil samples were dried for 2 wk, then cysts were extracted from soil samples using the elutriator system (USDA-APHIS, 2009). After year 3 in rotation, soil and roots from each microplot were homogenized and mixed, then 2 kg of soil was sampled. The soil samples were dried for 2 wk in labeled paper sample bags. Cysts were extracted from the soil samples by elutriator (USDA-APHIS, 2009).
Following extraction, cysts were further separated from debris by acetone floatation (Brodie et al., 1976) and then counted using a dissecting microscope (Leica M80). To determine the average eggs per cyst, the eggs of five cysts per replicate were counted when sufficient cysts were available. In samples with fewer than five progeny cysts, eggs of all cysts were counted. Cysts were then crushed and the eggs were counted with an inverted microscope (Leica DMi1). The reproduction factor (RF) was calculated as Pf/Pi, where Pi was the starting egg population density (7.5 eggs/g soil) and Pf was the final egg population density (eggs/g soil).
Data were analyzed using the SAS statistical package (SAS Institute Inc., Cary, NC). To determine whether data from repeated trials could be combined, analysis of variance was used to check for significant interactions between trial and treatment. If there were no interactions, data from trials were combined for further analysis. Rotation data were analyzed independently for each year. Egg count data were analyzed with a generalized linear mixed model (PROC GLIMMIX) assuming the Poisson distribution. The hatch and viability percentages were transformed by arcsin(sqrt(×/100)) before analysis to meet the assumptions of a normal distribution. To normalize the progeny cysts, Pf, and RF data, data were transformed by log10(x + 1) before analysis. The transformed data were then analyzed using a generalized linear mixed model (PROC GLIMMIX) assuming the normal distribution. Non-transformed means are shown in the tables. Means separation by least squares means was used to determine statistically significant differences between treatment means. Data were considered significantly different at P ≤ 0.05. Additionally, for egg hatch in PRD data to be considered significant, hatch in PRD for the treatments was compared to hatch in bare soil diffusate in a split-plot analysis of variance.
For the root diffusate collected at 4 wk of growth, G. pallida hatch was lowest in bare soil diffusate (1% hatch) and highest in susceptible potato ‘Désirée’ and ‘Russet Burbank’ diffusate (54% and 61% hatch) (Fig. 2A). The ‘Innovator’ hatching effect (41% hatch) was statistically the same as the S. sisymbriifolium hatching effect (36% hatch), and these were significantly lower than that of susceptible potato but significantly higher than that of bare soil (Fig. 2A). For the root diffusate collected at 6 wk of growth, hatch was again lowest in bare soil diffusate (3% hatch) (Fig. 2B). Hatch caused by root diffusate of ‘Désirée’ (54% hatch), ‘Russet Burbank’ (53% hatch), ‘Innovator’ (44% hatch), and S. sisymbriifolium (50% hatch) was statistically the same (Fig. 2B).
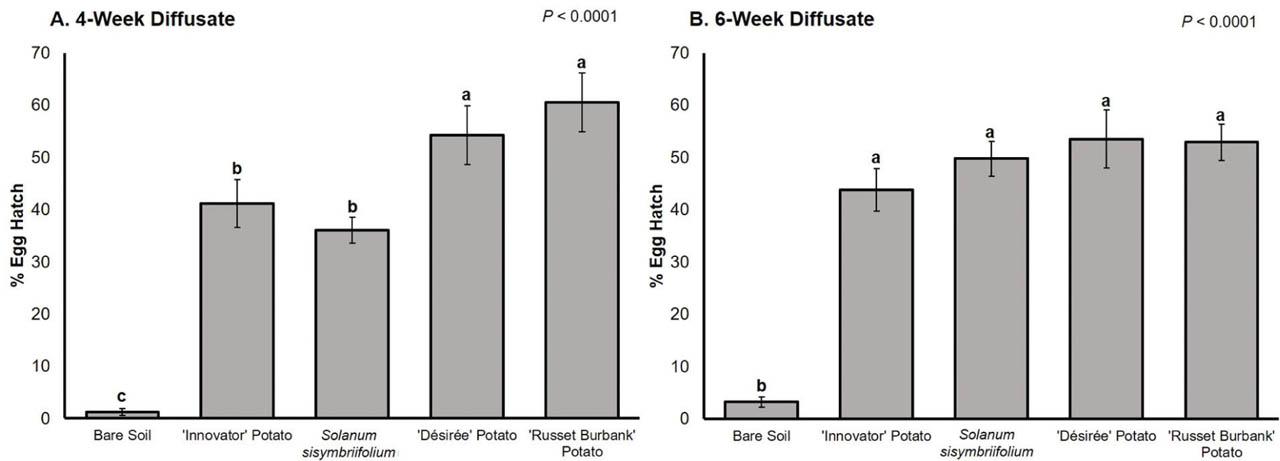
Mean percentage G. pallida egg hatch 2 wk after root exudate application. (A) Percentage egg hatch for root exudates was collected at 4 wk of growth. (B) Percentage of egg hatch for root exudates were collected at 6 wk of growth. Standard error of the means is indicated by the bars. Different letters indicate significantly different means based on least squares means at α = 0.05.
Three-year rotation sequences for the six rotation plans will be referred to henceforth using hyphenated sequence abbreviations as described in the methods (Table 1). When the remaining encysted eggs of the original population were assessed at the end of the first year of rotation, the remaining G. pallida encysted eggs were statistically the same when ‘Innovator’ or S. sisymbriifolium was planted (Table 2). In the first year, ‘Innovator’ and S. sisymbriifolium both reduced the average remaining encysted eggs per cyst by about 70% (Fig. 3). At the end of the first season, viable eggs were 39% lower for S. sisymbriifolium compared with ‘Innovator’ and hatch of remaining encysted eggs was not significantly different between the two treatments (P < 0.001) (Table 2). The year 1 Pf of S. sisymbriifolium (Pf = 0.79 eggs/g soil) was significantly lower than that of ‘Innovator’ (Pf = 1.28 eggs/g soil) (P < 0.0001) (Table 3).
Remaining G. pallida encysted egg densities, viability, and hatch at the end of the first year of rotation.a
| Year 1 treatment | Remaining encysted eggs per cyst | Remaining viable encysted eggs per cystb | Egg viability (%)b | Egg hatch in PRD (%)c |
|---|---|---|---|---|
| ‘Innovator’ resistant potato | 76.4 ± 5.5 | 63.8 ± 3.7 a | 88.5 ± 2.0 a | 4.2 ± 0.6 |
| S. sisymbriifolium | 73.0 ± 5.1 | 39.3 ± 2.5 b | 58.4 ± 3.6 b | 5.4 ± 0.7 |
Data presented are means of two trials with 15 replicates per trial. Numbers following means are standard error. Significant differences are denoted by different letters in the columns based on least squares means at α = 0.05.
Viable encysted eggs are determined through viability staining of remaining encysted eggs to reveal non-viable eggs.
Percentage hatch in PRD is the percentage hatch of the remaining encysted eggs in PRD.
PRD, potato root diffusate.
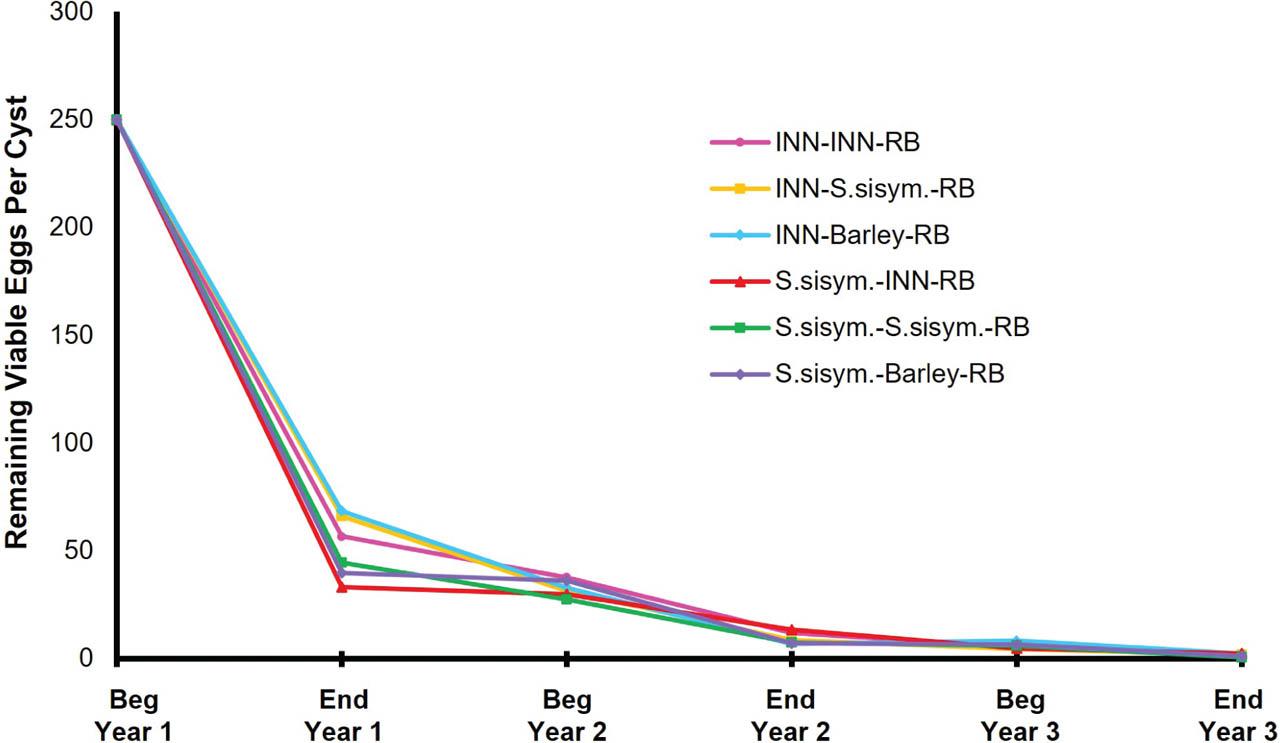
Remaining viable G. pallida encysted eggs per cyst of the initial G. pallida population over the 3-yr rotation. The initial G. pallida population at the beginning of year 1 in rotation started with an average of 250 eggs/cyst and an initial infestation rate of 7.5 eggs/g soil. INN, Innovator; RB, russet Burbank; S. sisym., S. sisymbriifolium.
Globodera pallida progeny cysts, final population (Pf), and RF over a 3-yr rotation.a
| Rotation sequenceb | End year 1 | End year 2 | End year 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Progeny cystsc | Pf (eggs/g soil)d | RF (Pf/Pi)e | Progeny cysts | Pf (eggs/g soil) | RF (Pf/Pi) | Progeny cysts | Pf (eggs/g soil) | RF (Pf/Pi) | |
| INN-INN-RB | 0.1 ± 0.1 | 1.28 ± 0.07 a | 0.26 ± 0.02 a | 1.0 ± 0.2 a | 0.36 ± 0.08 a | 0.07 ± 0.02 a | 0.5 ± 0.1 a | 0.06 ± 0.01 ab | 0.011 ± 0.002 ab |
| INN-S. sisym.-RB | 0.1 ± 0.1 | 1.28 ± 0.07 a | 0.26 ± 0.02 a | 0.1 ± 0.1 b | 0.17 ± 0.06 b | 0.04 ± 0.01 b | 0.2 ± 0.1 bc | 0.04 ± 0.01 abc | 0.008 ± 0.002 abc |
| INN-Barley-RB | 0.1 ± 0.1 | 1.28 ± 0.07 a | 0.26 ± 0.02 a | 0.3 ± 0.2 b | 0.16 ± 0 b | 0.03 ± 0.01 b | 0.4 ± 0.1 b | 0.07 ± 0.02 a | 0.013 ± 0.004 a |
| S. sisym.-INN-RB | 0 | 0.79 ± 0.05 b | 0.16 ± 0.01 b | 0 b | 0.27 ± 0.07 ab | 0.05 ± 0.01 ab | 0.1 ± 0 bc | 0.06 ± 0.01 ab | 0.010 ± 0.002 ab |
| S. sisym.-S. sisym.-RB | 0 | 0.79 ± 0.05 b | 0.16 ± 0.01 b | 0 b | 0.15 ± 0.04 b | 0.03 ± 0.01 b | 0 c | 0.02 ± 0.01 c | 0.003 ± 0.001 c |
| S. sisym.-Barley-RB | 0 | 0.79 ± 0.05 b | 0.16 ± 0.01 b | 0 b | 0.14 ± 0.04 b | 0.03 ± 0.01 b | 0.2 ± 0.1 b | 0.04 ± 0.01 bc | 0.007 ± 0.003 bc |
Data presented are means of two trials with five replicates per trial. Numbers following means are standard error. Significant differences are denoted by different letters in the columns based on least squares means at α = 0.05.
In the rotation sequences, the susceptible potato ‘Russet Burbank’ is abbreviated as ‘RB’, the resistant potato ‘Innovator’ is abbreviated as ‘INN’, and Solanum sisymbriifolium is abbreviated as ‘S. sisym.’. The sequence is given as Year 1-Year 2-Year 3.
Progeny cysts are cysts in 500-g soil sampled at the end of each growing season.
Pf in eggs/g soil is the final egg population at the end of the growing season calculated by adding the remaining viable eggs of the initial population and the eggs of the progeny cysts.
RF is reproduction factor calculated as final egg population (Pf) divided by initial egg population (Pi).
In years 2 and 3, there were no significant differences in the remaining encysted eggs, remaining viable encysted eggs, hatch, and viability of the initial G. pallida population for the different rotation sequences. Although there are no significant differences between the rotation sequences in years 2 and 3, the remaining viable encysted eggs of the initial population continue to decrease by about 50% season to season for all rotations (Fig. 3).
In general, there was low G. pallida reproduction for all rotation sequences. At the end of year 1, there were very low progeny cysts in ‘Innovator’ plots with 0–0.1 progeny cysts per 500-g soil (Table 3). Year 1 progeny cysts from ‘Innovator’ were not significantly different from that of S. sisymbriifolium (Table 3). The year 1 RF of ‘Innovator’ (RF = 0.26) was significantly greater than that of S. sisymbriifolium (RF = 0.16) (P < 0.0001), but both were <1, meaning the overall population was reduced (Table 3). In the second year, progeny cysts (P < 0.001) were significantly greater for the rotation sequence receiving 2 yr of ‘Innovator’ (INN-INN) compared with INN-S. sisym., INN-Barley, S. sisym.-INN, S. sisym.-S. sisym., and S. sisym.-Barley (Table 3). Also, at the end of year 2, Pf (P = 0.039) and RF (P = 0.039) were significantly greater for the rotation sequence receiving 2 yr of ‘Innovator’ (INN-INN) compared with INN-S. sisym., INN-Barley, S. sisym.-S. sisym., and S. sisym.-Barley (Table 3). At the end of year 3, the INN-INN-RB rotation produced the greatest progeny cysts (P < 0.0001) (Table 3). There were no detectible progeny cysts in the S. sisym.-S. sisym.-RB rotation sequence in year 3 (Table 3). By the end of year 3, the RF values indicated that the initial G. pallida population of 7.5 eggs/g soil was reduced by 99% to nearly 100% for all rotations (Table 3).
In vitro hatch assays showed that ‘Innovator’ and S. sisymbriifolium were alike in their effect on G. pallida egg hatch with significant egg hatch compared with a bare soil negative control whether 4- or 6-wk-old diffusate was used. However, the 4-wk-old diffusate hatch results showed ‘Innovator’ and S. sisymbriifolium had a hatching effect less than that of susceptible potato, while 6-wk-old diffusate from these two plants induced similar hatch as that of susceptible potato. This discrepancy may be explained by a variety of factors that can affect the root diffusate such as differences in plant growth, age, and health (Ochola et al., 2021). The chemical composition of root diffusate can change with the age of the plant and affect hatch (Byrne et al., 2001; Masler and Perry, 2018; Ochola et al., 2021). The hatching effect can also be impacted by concentration of the hatching factor in the diffusate, as higher concentrations can inhibit hatch (Ochola et al., 2021). Resistance genes are not known to affect root diffusate efficacy, so ‘Innovator’ resistant potato hatching effect being comparable to that of susceptible potato is an expected result (Ochola et al., 2021). In essence, the in vitro assays demonstrated that ‘Innovator’ and S. sisymbriifolium have the potential to cause a similar reduction in G. pallida egg densities through stimulating egg hatch when in rotation.
In the rotation experiment after year 1, the average remaining encysted egg densities and hatch of remaining eggs were the same for ‘Innovator’ and S. sisymbriifolium plots. This is likely because they caused a hatching event resulting in similar hatch percentage of the G. pallida population, as was shown in the in vitro hatching assays. However, viable eggs and viability percentage after year 1 were lower for S. sisymbriifolium than for ‘Innovator’. Non-viable eggs will not hatch during subsequent exposure to a host because their egg membrane has been disrupted (Pillai and Dandurand, 2019). S. sisymbriifolium may have an additional effect on egg viability. This effect may be the result of steroidal glycoalkaloids that S. sisymbriifolium produces (Pillai and Dandurand, 2021; Schulz et al., 2024). Pure glycoalkaloids have been found to decrease G. pallida hatch (Pillai and Dandurand, 2021). Schulz et al. (2024) fractionated chemicals from S. sisymbriifolium to try to isolate the toxic compounds and found some of the fractions to significantly reduce G. pallida hatch.
Most of the decline in the initial G. pallida population occurred in year 1 due to ‘Innovator’ and S. sisymbriifolium. As previous research has also shown, both of these crops are powerful tools against G. pallida in a crop rotation (Buckley, 2015; Dandurand and Knudsen, 2016; Dandurand et al., 2019a; Varypatakis et al., 2019; Mhatre et al., 2021). Throughout the rest of the rotation, viable encysted eggs of the initial G. pallida population continued to decline. This decline occurred even in rotation to the non-host barley, which should not simulate hatch of the nematode. While the barley itself may not have caused decline in G. pallida eggs, other factors likely contributed to natural attrition of the nematode such as degradation by microbes or spontaneous hatch (Storey, 1984; Brodie, 1996; Turner, 1996; Devine et al., 1999; Atkinson et al., 2001; López-Lima et al., 2013). Brodie (1996) found a 30%–40% decline in G. rostochiensis during rotation to fallow, independent of initial nematode density. Ryan and Devine (2005) reported 37% spontaneous hatch of G. pallida over a season of fallow. Microbial activity and spontaneous hatch have also been found to be positively correlated with soil temperature in the field (Devine et al., 1999). It is plausible that the high temperatures of southeastern Idaho, exceeding 26°C during much of the growing season, may have been a contributing factor in the decline in viable eggs in the 3-yr rotations. G. pallida is best adapted for temperatures between 10°C and 20°C, and temperature extremes can cause nematode mortality (Franco, 1979; Masler and Perry, 2018). Nonetheless, at the end of year 3, all rotation sequences had some remaining viable encysted eggs in the initial G. pallida population, which may not hatch until exposure to a host in subsequent seasons (Jones and Perry, 1978).
Reproduction of G. pallida was also evaluated over the 3-yr rotations. In years 1 and 2, there were a few progeny cysts recovered from ‘Innovator’ plots. ‘Innovator’ plots in rotation for 2 yr (INN-INN) had significantly higher progeny cysts and RF than the other rotations (INN-S. sisym., INN-Barley, S. sisym.-INN, S. sisym.-S. sisym., S. sisym.-Barley). However, the RF was much <1, indicating that the overall population of G. pallida was reduced when rotation included a resistant potato variety. The initial G. pallida population densities were reduced by all the 3-yr rotation sequences even after a susceptible potato in year 3 as evidenced by RF all being <1. Based on these results, it can be concluded that in Idaho conditions, incorporating resistant potato or S. sisymbriifolium in rotation before a susceptible potato is effective at reducing populations of G. pallida. Previous studies also support this finding as resistant potato can reduce G. pallida populations by 95% (Halford et al., 1999) and one crop of S. sisymbriifolium can reduce G. pallida by 90% on subsequently planted susceptible potato (Dandurand and Knudsen, 2016; Dandurand et al., 2019a; Mhatre et al., 2021). It is also important to note that no progeny cysts were recovered at the end of year 3, at 2-kg soil samples for the S. sisym.-S. sisym.-RB rotation sequence, indicating that cysts were non-detectable after a rotation sequence of 2 yr of S. sisymbriifolium prior to planting the susceptible variety ‘Russet Burbank’.
In conclusion, a highly resistant potato like ‘Innovator’ and the trap crop S. sisymbriifolium were both valuable in a crop rotation to reduce G. pallida population densities in Idaho. Both ‘Innovator’ and S. sisymbriifolium caused a hatching effect comparable to that of susceptible potato and similarly reduced G. pallida populations with a 70% reduction of encysted eggs in the first season. Although a highly resistant potato like ‘Innovator’ still allowed some reproduction of G. pallida, it resulted in an overall drastic decline in the population densities in rotation. Development of a resistant russet potato for Idaho is imperative because it could be used in rotation to control G. pallida and keep it below detectable levels while providing a profitable yield, as is done with G. rostochiensis in New York (Brodie and Mai, 1989; Brodie, 1996; Dandurand et al., 2019b). It is important to recognize that even with the use of a resistant potato or trap crop to reduce G. pallida, such crops must be continuously used in a rotation with susceptible potato to avoid resurgence of the population, as some viable eggs still survive season to season (Brodie, 1996; Turner, 1996; Devine et al., 1999; Ryan and Devine, 2005). Although all rotations incorporating ‘Innovator’ and S. sisymbriifolium before susceptible potato ‘Russet Burbank’ were shown to reduce the initial G. pallida population by about 99% to nearly 100%, few viable encysted eggs were still found at the end of year 3 in all plots. Ultimately, this study demonstrated that crop rotation with a highly resistant potato or the trap crop S. sisymbriifolium has great potential to control G. pallida in Idaho.