
The welfare of ruminant farming is greatly affected by infections caused by gastrointestinal nematodes (GIN). Economic losses due to these parasites primarily manifest as reduced production rather than mortality, with estimates indicating that GIN infections contribute to an annual production cost of approximately €1.8 billion across the EU, including both production losses and treatment expenses (Charlier et al., 2020; Charlier et al., 2024). Therefore, infections caused by GINs are usually classified as a production disease (Marshall et al., 2012).
The most common GINs responsible for economic losses in small ruminants belong to the family Trichostrongylidae (trichostrongylids), particularly Haemonchus contortus, Trichostrongylus spp., and Teladorsagia circumcincta (O’Connor et al., 2006; Gilleard, 2013). The severity of infection depends on various factors, with the composition of the parasite species being one of the most crucial. Certain GIN species are more pathogenic to their hosts than others. In natural infections, hosts are usually infected by multiple GIN species simultaneously, whereas mono-infections are rare (Leignel & Cabaret 2001; Demeler et al. 2012).
Sheep and goats harbour identical GIN species (Hoste et al., 2008; Domke et al., 2013); however, some data suggest the possibility of different nematode strains between sheep and goats (Sutherland & Scott, 2010). Most existing data on host-parasite interactions have been collected from studies focusing solely on sheep, and their results have been applied to goats. It is assumed that, due to different evolutionary processes, sheep and goats have developed distinct strategies for controlling GIN infections – sheep rely primarily on their immune response. In contrast, goats are more dependent on grazing behavior (Houdijk et al., 2012). These two strategies rely on a balance between developing an immune response in sheep or a behavioural response that limits contact with infective larvae present in the vegetation in goats. In goats, avoidance of infective larvae associated with grazing is suspected to be high due to their grazing behaviour (Hoste et al., 2010). Observations conducted under natural grazing conditions in Australia and Scotland indicate that goats exhibit higher levels of parasitic infection than sheep when co-grazing, supporting the hypothesis of a weaker immune response in goats to GIN infections (Torres-Acosta & Hoste, 2008). Specifically, goats may take approximately 12 months to develop a complete immune reaction, whereas sheep typically develop immunity within six months (Vlassoff et al., 1999). Moreover, in sheep, the manifestations of nematode infection in adult females are much less severe compared to young animals, unlike goats, for which the response to parasites is not well described (Hoste et al., 2008). Furthermore, adult goats excrete significantly larger quantities of parasite eggs in their faeces compared to sheep. This difference is likely due to physiological and immunological variations between the species. Although extensive research has been conducted on GIN infections in sheep and goats worldwide, data on co-grazing systems under Central European climatic conditions remain scarce. Most available studies have focused on Southern or Northern European regions, where environmental factors significantly differ from those in Central Europe (Kyriánová et al., 2019; Ruiz-Huidobro et al., 2019). The present study aims to fill this knowledge gap by examining the GIN load and species richness of co-grazing dairy sheep and goats in the Czech Republic. Understanding these dynamics is crucial for optimizing parasite management strategies in mixed-species grazing systems in temperate climates.
This study tests the hypothesis that quantitative descriptors of GIN populations differ significantly between sheep and goats in co-grazing systems. The objectives of our study were: (i) to compare the intensity of GIN infections in sheep and goats; (ii) to identify the nematode species richness in sympatric sheep and goats.
Our study was conducted over a continuous 12-month period, from February to January of the following year, on a conventional dairy farm in the northern Bohemia region, Czech Republic. The farm is situated at an altitude of 320 meters above sea level in a slightly warm, drier area with an average annual temperature of 9.1°C and an average annual rainfall of 670 mm. Basic meteorological data for the study period were obtained from the nearby meteorological station (see Fig. 1).

Average monthly temperature (°C) and precipitation (mm) recorded during the 12-month monitoring period. Data were obtained from a nearby meteorological station and reflect typical temperate climate conditions of the study area. Seasonal variation in temperature and rainfall may influence parasite development and transmission.
The base stock consisted of more than 200 animals, all of which were kept under identical farm conditions. About half of the flock comprised dairy goats (White Shorthaired and the Brown Shorthaired goat, and their crossbreds), while the other half consisted of dairy sheep (Laucane, East Friserian sheep, and their crossbreds). All animals had year-round unrestricted access to 45-hectare pastures, divided into smaller fenced areas with rotational grazing across four pastures. The rotation schedule was not fixed but depended on pasture condition and growth, with animals typically spending 2 – 3 weeks in each pasture.
In the barn, the animals had access to hay, water, and a partial mixed ration, while concentrates were provided during milking. The complete feeding regimen, including the botanical composition of pastures and other dietary components, was characterized in detail by Ptáček et al. (2021).
The farm processes milk into goat and sheep cheese, yoghurts, and other products, which it sells at local farmers’ markets. Animals were dried off based on milk production and udder status, but always at least eight weeks before the expected parturition. The mating period occurred from August to September, with the lambing, or kidding, period taking place from January to March. Lambs and goat kids were separated from their mothers immediately after birth and reared artificially on milk replacements. Granulates and hay were offered in the creeping pen from the 2nd week of age. Lambs and kids were weaned after achieving 18 kg of their live weight.
Average milk yield during the lactation period was 3.04 kg/day in goats and 1.55 kg/day in sheep. Detailed monthly values for milk yield and basic composition parameters are provided in Table 1. These data were recorded as background herd characteristics; the present study did not assess milk composition or relate it to gastrointestinal nematode infection.
Monthly milk production and composition in monitored goats and sheep during the lactation period. Values include milk yield (kg/day) and percentages of fat, protein, and lactose. Data were collected as part of the official milk recording system in the Czech Republic. Results represent the average performance of the sampled animals over five (goats) or four (sheep) monthly recordings.
| Goats - monitoring performance check results | ||||
|---|---|---|---|---|
| milk (kg) | fat (%) | protein (%) | lactose (%) | |
| III. | 2.9 | 4.00 | 3.15 | 4.56 |
| IV. | 3.0 | 2.69 | 2.72 | 4.18 |
| V. | 3.1 | 2.80 | 2.95 | 4.32 |
| VI. | 3.3 | 2.70 | 2.96 | 4.52 |
| VII. | 2.9 | 2.93 | 3.01 | 4.32 |
| mean | mean | mean | mean | |
| Sheep - monitoring performance check results | ||||
|---|---|---|---|---|
| milk (kg) | fat (%) | protein (%) | lactose (%) | |
| III. | 1.5 | 7.61 | 5.39 | 4.83 |
| IV. | 2.0 | 5.62 | 5.64 | 4.70 |
| V. | 1.3 | 7.03 | 6.03 | 4.72 |
| VI. | 1.4 | 7.27 | 6.42 | 4.68 |
| mean | mean | mean | mean | |
Weaned animals and adult males were kept separately on pasture, with or without shelters, and with or without rotational grazing. The herd operated under a closed turnover policy, except for the introduction of new sires to renew the bloodline. Newly purchased males were not quarantined; they were treated with anthelmintics before being integrated with the rest of the herd. Specifically, the anthelmintic product “First Drench” (containing abamectin and praziquantel) was used at dosages recommended by the attending veterinarian for both sheep and goats, typically administered orally at a dosage of 2.5 ml per 10 kg body weight. Weaned lambs and goat kids received anthelmintic treatment individually and only when necessary, based on a combination of clinical signs (e.g., altered faecal consistency, reduced body condition score) and, in some cases, faecal egg count results provided by the farm veterinarian. Dairy animals did not receive any anthelmintic or anticoccidial treatment during the study period or within at least six months preceding its onset.
Twenty dairy goats and twenty dairy sheep were randomly selected for monthly rectal sampling over 12 months. The same individually numbered animals were followed throughout the study. Sampling was performed before the afternoon milking, when the animals were housed indoors; sheep and goats were separated for faecal collection and subsequent milking. Samples were stored in labelled plastic bags at 4 °C and examined the following day.
Over the course of the study, a total of 210 goat and 196 sheep faecal samples were collected. The difference from the expected maximum (240 samples per species) reflects occasional instances where faecal collection was not possible, for example, due to an empty rectum or animal handling constraints. This resulted in a small number of missing monthly samples per individual.
Nematode eggs were quantified using the Concentration McMaster method (Roepstorff & Nansen, 1998), with a sensitivity of 20 nematode eggs per gram (EPG). This method was selected due to its simplicity and cost-effectiveness, making it suitable for routine faecal egg count monitoring under farm conditions. Both the prevalence and infection intensity were evaluated as described by Bush et al. (1997).
Because morphological differentiation of nematode eggs to genus level is generally unreliable, all strongyle-type eggs were merged into strongylid nematodes. The remaining portions of individual faecal samples, which were not used for quantitative egg counts, were pooled and subjected to coproculture for larval development and incubated for seven days at 27 °C to obtain infective larvae (L3) for reliable strongylid identification. Recovered L3 were identified to the genus or species level according to van Wyk and Mayhew (2013). Trichostrongylus spp. and Teladorsagia spp. L3 were merged into one group (Trichostrongylus/Teladorsagia) due to their similar morphology, which precluded reliable differentiation.
A power analysis was conducted before the survey to determine the effect of sample size. Differences in GIN infection intensities between goats and sheep were analysed using the Mann-Whitney test, with statistical significance set at α = 0.05. Statistical calculations were performed using Statistica 12 (StatSoft ČR, 2012). Violin plots comparing infection intensities in sheep and goats were generated using R statistical software, version 3.1.2 (R Development Core Team, 2014). In addition to the primary analyses, supplementary correlation analyses were performed separately for sheep and goats to explore potential relationships between milk production parameters and the intensity of gastrointestinal nematode infection, measured as faecal egg counts (EPG).
No animals were harmed during the course of the study. Faecal samples were collected non-invasively during routine handling, and all applicable national and institutional guidelines for animal welfare were strictly adhered to.
Over the 12-month monitoring period, a total of 210 samples from goats and 196 samples from sheep were analysed (see Table 2). The overall prevalence of trichostrongylid nematodes was 81 % (158/196) in sheep and 96 % (202/210) in goats. Trichuris eggs were also detected in goats, with an overall prevalence of 6.1 % and a maximum EPG of 180 in June. In sheep, Trichuris eggs were found only once, at a level of 20 EPG, also in June.
Summary of faecal egg counts (EPG), prevalence, and number of samples analysed in sheep and goats throughout the 12-month monitoring period. EPG values reflect the intensity of trichostrongylid infection, while prevalence indicates the proportion of positive samples per species. The table also includes the total number of samples obtained, accounting for minor occasional sampling losses due to technical limitations.
| Trichostrongylids - sheep | ||||||||
|---|---|---|---|---|---|---|---|---|
| prevalence (%) | median | mean | geomean | CI (%) | SD | pooled samp. (EPG) | min. EPG | max. EPG |
| 100 | 1780 | 1747 | 1648 | 1477 – 2017 | 543 | 1340 | 720 | 2520 |
| 75 | 1120 | 990 | 1254 | 669 – 1311 | 687 | 300 | 0 | 1980 |
| 40 | 0 | 457 | 1103 | 175 – 739 | 602 | 180 | 0 | 1520 |
| 86 | 140 | 1087 | 326 | 96 – 2078 | 1717 | 460 | 0 | 5040 |
| 60 | 100 | 627 | 501 | 60 – 1193 | 1023 | 0 | 0 | 3640 |
| 80 | 500 | 856 | 472 | 305 – 1407 | 996 | 0 | 0 | 2600 |
| 93 | 280 | 692 | 314 | 170 – 1214 | 943 | 120 | 0 | 3540 |
| 94 | 530 | 607 | 1533 | 385 – 829 | 446 | 0 | 0 | 1520 |
| 80 | 20 | 375 | 107 | 0 – 872 | 899 | 140 | 0 | 3520 |
| 79 | 40 | 189 | 85 | 0 – 396 | 360 | 0 | 0 | 1260 |
| 93 | 700 | 804 | 643 | 459 – 1149 | 623 | 520 | 0 | 2140 |
| 94 | 1760 | 1639 | 1202 | 1007 – 2270 | 1228 | 580 | 0 | 3900 |
| Trichostrongylids - goats | ||||||||
|---|---|---|---|---|---|---|---|---|
| prevalence (%) | median | mean | geomean | CI (%) | SD | pooled samp. (EPG) | min. EPG | max. EPG |
| 90 | 180 | 316 | 214 | 166 – 466 | 319 | 680 | 0 | 1140 |
| 100 | 510 | 910 | 478 | 341 – 1479 | 1145 | 400 | 20 | 4380 |
| 100 | 900 | 1222 | 721 | 692 – 1752 | 1100 | 2720 | 80 | 3500 |
| 100 | 3500 | 4254 | 2813 | 2526 – 5981 | 3584 | 520 | 120 | 13740 |
| 100 | 5090 | 6555 | 4144 | 3775 – 9336 | 5591 | 540 | 340 | 20980 |
| 89 | 2160 | 2788 | 1992 | 1427 – 4148 | 2736 | 160 | 0 | 10220 |
| 100 | 940 | 1760 | 933 | 803 – 2717 | 1924 | 120 | 40 | 7380 |
| 100 | 730 | 1292 | 716 | 581 – 2003 | 1430 | 100 | 20 | 5500 |
| 100 | 560 | 734 | 428 | 336 – 1132 | 774 | 240 | 80 | 2840 |
| 94 | 160 | 531 | 325 | 228 – 834 | 589 | 480 | 0 | 1880 |
| 92 | 100 | 138 | 111 | 63 – 214 | 119 | 320 | 0 | 380 |
| 88 | 260 | 239 | 194 | 143 – 335 | 180 | 340 | 0 | 540 |
The seasonal dynamics of egg shedding, as depicted in Table 2, show that both species experienced fluctuations in nematode transmission throughout the year. However, these seasonal changes did not result in consistent differences in infection intensity between goats and sheep.
When data from the entire 12-month period were analysed, the overall infection intensity, expressed as eggs per gram of faeces (EPG), was significantly higher in goats compared to sheep (Mann-Whitney U test, U = 24,697.5, p < 0.001). Despite comparable prevalence, goats showed greater individual variability and higher maximum EPG values (max. 20,980 EPG in June), whereas the maximum in sheep was 5,040 EPG (in May). The distribution of infection intensity is visualized in Fig. 2. Violin plots reveal that while both groups share overlapping ranges of EPG values, goats tend to exhibit higher median values and more pronounced tails, reflecting a broader distribution and a greater number of extreme cases. This confirms greater heterogeneity of infection among goats and supports the statistical findings. Differences in infection intensity between sheep and goats were statistically significant (Mann–Whitney U = 24 697.5, p < 0.001).

Comparison of strongylid nematode infection in sheep and goats. Data are expressed as eggs per gram of faeces (EPG) and were log-transformed for visualization. Violin plot displays the distribution, median (white dot), interquartile range (thick bar), and density. Higher median and variability were observed in goats. Differences between species were statistically evaluated using the Mann-Whitney U test (U = 24,697.5, p < 0.001).
A more detailed monthly analysis (see Table 2) revealed significant differences in infection intensity across several months. Sheep had significantly higher EPG values in February (U = 5.0, p < 0.001), whereas goats had significantly higher EPG values in April, May, June, July, August, October, and November (all p < 0.05). No significant differences were found in March and September. These results indicate seasonal shifts in the parasite burden, with goats showing peak infection during the summer months and sheep experiencing a transient peak in late winter. Spearman’s rank correlation analysis was conducted to evaluate the relationship between the intensity of strongylid nematode infection in goats and sheep. There was a weak but statistically significant negative correlation between the two variables (R = –0.20, p = 0.0028, n = 218), suggesting a slight tendency for higher infection levels in goats to coincide with lower infection levels in sheep. However, the strength of this association was low.
Haemonchus contortus was the most frequently detected species and remained dominant in the goat population throughout the year. In sheep, the dominant species shifted seasonally, with Trichostrongylus/Teladorsagia spp. becoming more prominent during certain months. Oesophagostomum columbianum was detected at lower levels in both host species.
Clear seasonal patterns were also observed in the larval composition. In sheep, H. contortus declined markedly in October (Fig. 3), followed by an increase in winter months, possibly reflecting microclimatic influences, larval hypobiosis, or delayed development. In goats, Trichostrongylus/Teladorsagia spp. peaked in July (Fig. 4) with a subsequent decline, while sheep showed a peak in October. O. columbianum (Fig. 5) was reduced in summer (June – July)in both species and showed variable increases during autumn and winter.
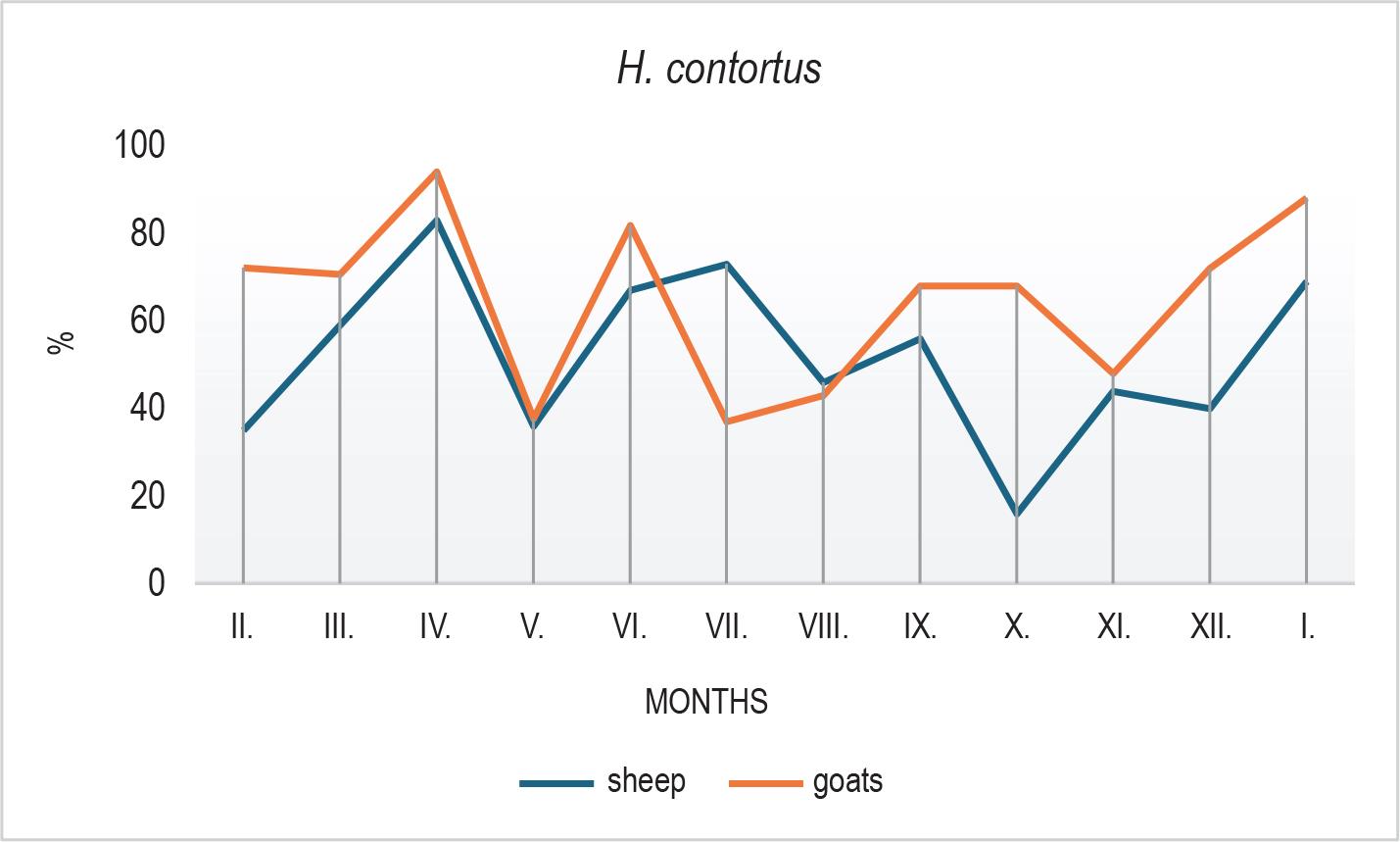
Seasonal composition of Haemonchus contortus larvae isolated from faecal cultures of goats and sheep over the 12-month monitoring period. H. contortus predominated in goats throughout the year, while in sheep, its prevalence fluctuated seasonally, with a decline in autumn and resurgence in winter months.
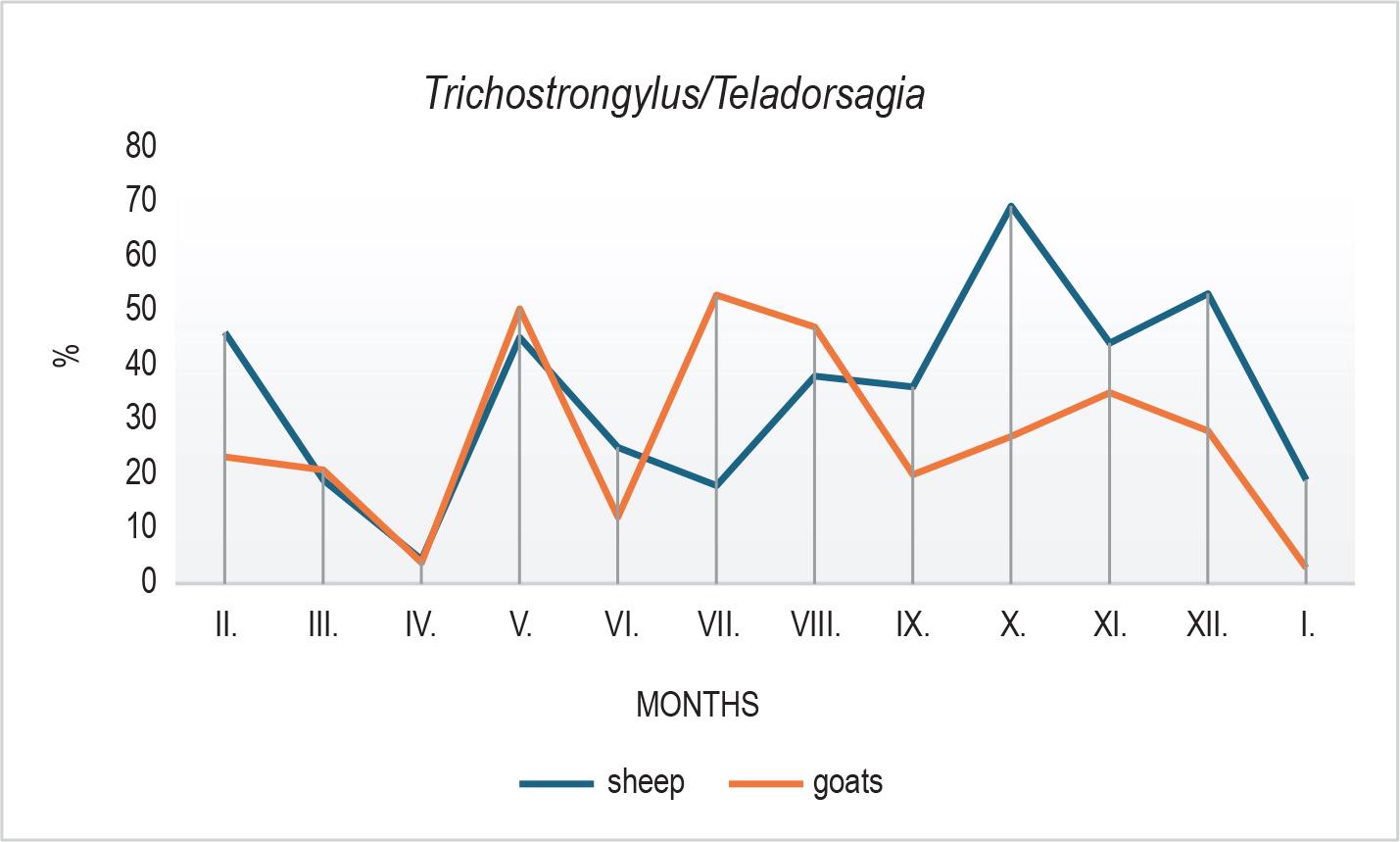
Proportion of Trichostrongylus/Teladorsagia spp. larvae recovered from faecal samples during the study. In goats, their abundance increased during the summer, while in sheep a marked peak was observed in October. The grouping reflects the indistinguishable morphology of L3 larvae from these two genera.

Seasonal dynamics of Oesophagostomum columbianum larvae identified in coprocultures from goats and sheep. A general decline was observed during the summer (June–July) in both host species, followed by fluctuating increases in the autumn and winter months.
Co-grazing of sheep and goats is practiced in many temperate and Mediterranean regions, but its effect on GIN epidemiology remains debated. Goats and sheep share most GIN species, with goats often producing higher egg counts during the grazing season, which may potentially increase the infection risk for sheep (Hoste et al., 2008; Hoste et al., 2010). In this study, we compared GIN species richness and infection intensity in sheep and goats co-grazed on a conventional dairy farm.
We found no significant difference in GIN species richness between sheep and goats. In both species, Haemonchus contortus was the most prevalent nematode, followed by Trichostrongylus/Teladorsagia and Oesophagostomum columbianum. However, H. contortus predominated in goats throughout the year, while in sheep, Trichostrongylus/Teladorsagia spp. showed periodic dominance. This seasonal variation aligns with previous studies (Torina et al., 2004b; Hosté et al., 2010; Domke et al., 2012; Kelly et al., 2012; Domke et al., 2013; Kyriánová et al., 2019; Ruiz-Huidobro et al., 2019). Torina et al. (2004) highlight the complexity of parasite dynamics in mixed-species grazing systems. In addition to the dominant species of the genus Trichostrongylus, less abundant species were also found, of which H. contortus was the most common. In contrast, in goats, the authors reported only T. circumcincta and T. axei as the dominant nematodes in the abomasum; all other nematode species were less represented.
While numerous studies have examined GIN infections in sheep and goats, most of them focused on systems where the species were reared separately. Studies specifically addressing co-grazing systems remain limited. However, recent research using metabarcoding approaches has provided new insights into the dynamics of GIN infections under mixed-species grazing. Notably, Costa-Junior et al. (2021) and Mohammed et al. (2024) observed that Haemonchus contortus was consistently dominant in goats, whereas sheep harbored more diverse nematode communities, including Teladorsagia and Trichostrongylus species. Both studies also reported seasonal shifts in species composition, underscoring the influence of host species and seasonality on parasite epidemiology. Our results align with these findings. In goats, Haemonchus contortus predominated throughout the year, whereas in sheep, Trichostrongylus and Teladorsagia spp. showed periodic dominance. These patterns suggest that parasite dynamics may differ between co-grazed species, underlining the potential benefit of considering host-specific factors in parasite control. Further comparative research is needed to evaluate whether control strategies developed for single-species systems are equally effective in co-grazing contexts. This is particularly relevant considering our findings that, despite distinct seasonal peaks—such as the higher mean EPG in goats in June and in sheep in February—the overall infection pressure in both groups remained stable and of similar magnitude throughout the study period. Although individual months revealed significant differences in EPG between host species, these differences were inconsistent and likely reflect short-term environmental or physiological fluctuations. These results support the conclusion that co-grazing, as practiced in this study under a rotational pasture system and without recent anthelmintic treatment, does not increase the risk of gastrointestinal nematode infection in either sheep or goats. While species-specific responses to GIN exposure were observed, the comparable prevalence and long-term infection intensity across hosts suggest that mixed-species grazing per se is not a primary driver of elevated parasite transmission under the given conditions.
The presence of GIN infective larvae on the vegetation and their availability to the host are influenced by numerous external and internal factors. Key external factors include local climatic conditions and microclimatic conditions on the pasture. Temperature and moisture play a dominant role in the development of the trichostrongylid free-living stages (O’Connor et al., 2006). Interestingly, H. contortus was most prevalent during winter in our study, contrary to its typical ecological pattern. Possible explanations include specific microclimatic conditions, pasture management practices that affect larval survival, or the resumption of development from hypobiotic larvae. Further research is needed to clarify these mechanisms.
The large pasture area, relative to the number of animals, likely reduced larval density through host dispersal and natural mortality (Torres-Acosta & Hoste, 2008). Faecal moisture and dense vegetation may also prolong larval survival (van Dijk & Morgan, 2011). Internal factors, such as host immunity, differ between sheep and goats, with goats’ browsing behaviour potentially reducing contact with infective larvae but also limiting immune development (Houdijk et al., 2012). Another key factor influencing nematode prevalence is anthelmintic resistance (AR). Anthelmintic resistance, reportedly higher in goats, may further contribute to the dominance of H. contortus (Hoste et al., 2011).
Furthermore, H. contortus is known for its high fecundity, with females laying up to 10000 eggs daily, compared to Trichostrongylus/Teladorsagia, which produce only around 100 eggs per day. This reproductive advantage means that even at similar infection levels, H. contortus has a disproportionately higher impact on pasture contamination. Finally, differences in host specificity between ovine and caprine GIN lineages may play a role in infection dynamics. Studies suggest that sheep are less susceptible to caprine GIN lineages, which could further affect nematode transmission dynamics in co-grazing systems.
These findings highlight the complexity of host-parasite interactions in mixed-species grazing systems and underscore the need for further research into environmental, genetic, and management factors influencing GIN epidemiology. The cross-transmission of resistant nematodes between species, as well as between domestic and wild ruminants, has been recently documented, further emphasizing the importance of sustainable parasite control strategies (Beaumelle et al., 2024). Additionally, studies on anthelmintic resistance in small ruminants suggest that goats exhibit higher resistance levels, likely due to more frequent and often suboptimal treatment applications. Hoste et al. (2010) emphasized that improper dosing, combined with the pharmacological differences between goats and sheep, may contribute to selection pressure and subsequent shifts in nematode species composition.
During our study, no significant differences in the intensity of GIN infections were detected between sheep and goats. Similar patterns were observed in India by Singh et al. (2021) and Kumar et al. (2019), who reported comparable EPG values across both species, with infection levels strongly influenced by seasonal conditions. In contrast, Domke et al. (2013) observed higher EPG levels in sheep than in goats. Our study confirmed that goats shed increased numbers of eggs during the season; however, this shedding pattern was not consistent and did not appear to impact infection intensity in sheep significantly. The observed seasonal variations and differences in infection intensities between sheep and goats suggest that multiple factors, including environmental conditions, anthelmintic resistance, and host-specific immune responses, contribute to shaping the dynamics of GIN infections in mixed-grazing systems. The dominance of H. contortus in goats may be linked to its higher fecundity and ability to develop resistance to anthelmintics more rapidly than other nematode species. Additionally, differences in host grazing behaviour and susceptibility to specific nematode lineages could further explain the infection trends seen in this study. Future research should focus on integrating detailed environmental monitoring, genetic analysis of nematode populations, and long-term studies on anthelmintic resistance to develop effective parasite management strategies for mixed-species grazing systems.
Overall, our results showed that goats consistently exhibited higher GIN infection intensities than sheep, with mean peak faecal egg counts of 1 240 EPG in June compared to 620 EPG in February in sheep. Haemonchus contortus predominated in goats year-round, while Trichostrongylus/Teladorsagia spp. in sheep displayed pronounced seasonal peaks, with coproculture of larvae showing a predominance of up to 60 % Trichostrongylus/Teladorsagia larvae in sheep in the autumn. These findings fulfill the study aim of characterizing species composition and infection intensity in co-grazed small ruminants, supporting the hypothesis that co-grazing does not homogenize parasite burdens between host species. The observed species-specific and seasonal patterns highlight the need for targeted, host- and season-specific parasite control strategies, as well as further research into environmental and management factors that influence GIN epidemiology in mixed-species grazing systems.