
Soil moisture determines some of the physical and chemical properties of soil and affects its biological activity. Among the physical properties of soil modified by water content, soil aeration and soil colloid stability can be distinguished (Furtak, Gałązka, 2019a). Water content primarily affects soil pH, solvent and gas diffusion, and salt solubility. It also plays a role in hydrolysis processes and determines the rate of carbon and nitrogen mineralization (Wolińska, 2010). It has been shown that soil moisture correlates with the abundance of bacteria and fungi (Czaban et al., 2010). Natural fluctuations in moisture content associated with seasonal changes and precipitation are an important environmental factor in the metabolism of microorganisms (Nguyen et al., 2021; Peng et al., 2023). Recently, the frequency of various hydrological phenomena, such as floods and periodic flooding, has increased in Poland. These phenomena cause hydrological stress, which affects the functioning of soil microbiomes (Furtak, Wolińska, 2023; Young, Ritz, 2000).
Excessive soil moisture can be caused by flooding, thawing and heavy rainfall (Basri et al., 2022; Zhu et al., 2023). Improper land use can lead to water accumulating in the soil. After winter, waterlogging is often observed, which is often associated with inefficient or inadequate drainage, overgrown ditches, or excessive soil compaction. Although excess water is much less common than drought, the effects of prolonged and increased moisture are more severe for the soil environment (Chmura et al., 2009; Oleszczak, 2016). Excessive moisture in the soil can decrease its fertility by promoting the growth of pathogenic microorganisms, reducing gas diffusion, leaching nutrients (including phosphorus, potassium, calcium and magnesium) and destroying the soil’s structure. It can also introduce contaminants (Khoshru et al., 2023; Singh et al., 2022).
Flooding affects soil microorganisms, including those responsible for biogeochemical processes and promoting plant growth (Furtak, Wolińska, 2023). The disruption of the soil microbiome caused by flooding can be short- or long-term, but there is no data indicating when soil microbiology returns to equilibrium and ecosystem functions. When flooding occurs, the greatest challenge for soil and the organisms living in it is the drastic decrease in oxygen (Martínez-Arias et al., 2022). Osmotic stress and hypoxia can disrupt plant metabolism and affect microorganisms. Research by Unger et al. (2009) indicated that stagnant water in greenhouses was associated with reduced biomass of microorganisms and decreased levels of markers for aerobic bacteria, Gram-negative bacteria, Gram-positive bacteria, and mycorrhizal fungi. Studies in the model experiment have shown that flooding leads to a significant increase in the abundance of bacterial taxa capable of anaerobic respiration, such as members of the phyla Firmicutes and Desulfobacterota. At the same time, there is a decrease in the abundance of Actinobacteria and Proteobacteria, as well as the depletion of several taxa that are presumably beneficial to plants (Francioli et al., 2021). Soil biodiversity and microbiological activity directly translate into plant growth and yield, which is extremely important in areas affected by natural disasters such as floods (Baldi, 2021; Barros-Rodríguez et al., 2021). Currently, national and international literature lacks research on the impact of excessive moisture on agricultural soils and the impact of periodic flooding on soil quality in floodplains (Furtak, Wolińska, 2023). The most significant unknown factor is the impact of flooding on the soil microbiome, as well as the mechanisms that regulate changes in microbial communities and their resilience in flooded soils (Das et al., 2025).
The study aimed to conduct a preliminary assessment of changes in the soil seven months after the flood, comparing them with soil samples taken seven days after the floodwaters receded. Two different types of soil taken from fields flooded in 2024 in the Lower Silesia and Opole regions were analysed. These are pilot studies that only monitor basic parameters of biological soil quality after floods have occurred.
Soils from two fields affected by flooding in 2024 were selected for analysis (Table 1, Figure 1).
Soil samples using in research.
| Sample | Localisation | Before flood# | After flood# |
|---|---|---|---|
| O1 | Kantarowice (50°44′41.4″N, 17°35′32.1″E), municipality Lewin Brzeski, Opole voivodeship; floodwater from Nysa Kłodzka | winter wheat conventional cultivation | 2024: Ploughing; microbiological preparation; |
| 2025: nitrogen fertiliser, chicken manure, disc harrowing, maize sowing | |||
| D3 | Krosnowice (50°24′04.5″N, 16°38′10.4″E), municipality Kłodzko, Lower Silesia voivodeship; floodwater from Nysa Kłodzka | winter wheat conventional cultivation | 2024: Ploughing; |
| 2025: NPK+S+Mg fertiliser, soybean sowing | |||
Information from farmers.

Photographs from fields selected to research: A) field O1, year 2024 ; B) field D3, year 2024 (auhor’s photographs, K.F.).
The selected locations were affected by flooding in September 2024 (Figure 1). In both cases, the water stagnated in the fields for about 7 days. The selected fields were visited by IUNG-PIB employees during a study visit in September 2024. Soil samples were taken twice: in September 2024 (approx. 7 days after the flooding subsided), and in April 2025 (approx. 7 months after the disaster). Soil was collected from random points (> 6) in a given field, from a depth of 0–20 cm using soil probe, in a total amount of approx. 0.5 kg from each field, into sterile zip-lock bags, then placed in a refrigerator (4 °C) and transported to the laboratory, where was sieved through a 2 mm sieve.
The pH was measured potentiometrically (1:2.5; 96 mV) after 24 h of incubation soil in sterile water (1:1) at room temperature (edge® Multiparameter pH meter, HANNA Instrument, Woonsocket, RI, USA).
The activity of enzymes in soils was measurement spectrophotometrically (Spectrophotometer UV-Vis Evolution™ 60, Thermo Fisher Scientific, Waltham, MA, USA). For dehydrogenases (DHa) determination the 2,3,5 – triphenyl-tetrazolium chloride (TTC) was using as a substrate (Casida et al., 1964).
For analysis of alkaline (AlP) and acid (AcP) phosphatases activities an ρ-nitrophenyl phosphate (ρ-NPP) was used as a substrate (Tabatabai, 1982).
Analyses of enzymatic activity were performed in triplicate on each soil samples.
The diversity of the metabolic potential of the soil bacterial community was determined using the Biolog Eco-Plate™ (Biolog Inc., Hayward, CA, USA). The 96-well plates contain 31 carbon substrates classified into five biochemical groups: carbohydrates, carboxylic and ketonic acids, amines, polymers, and amino acids, in three replicates (Gałązka, Furtak, 2019). 1 g of fresh soil sample was suspended in 99 mL of sterile water, shaken for 20 min. and incubated at 4 °C for 30 min. (Weber, Legge, 2009). Next, each of the 96 wells of the microplate was inoculated with 120 μl of soil inoculums. All EcoPlates™ were incubated at 25 °C for 144 h. The intensity of the wells’ colour development was determined spectrophotometrically at λ = 590 nm (Insam, Goberna, 2004) for a period 144 h at 24-intervals using a MicroStation ID (Biolog Inc., Hayward, CA, USA) plate reader at OD590. The most intensive metabolic activity was observed after 120 h of incubation and these results are presented in manuscript. Analysis were replicated three times.
Statistical analyses were performed using the packet Statistica.PL ver. 10.0 (StatSoft. Inc., Tulsa, OK, USA). Significant differences were calculated according to post hoc Tukey’s HSD test at the P ≤ 0.05 significance level. Diagrams were performed using MS Excel software (Microsoft Corporation, 2021). The results obtained at 120 hours incubation of EcoPlates™ Biolog® were used to calculate the AWCD index (average well colour development) and Shannon’s diversity index (H’) according to Furtak et al. (2019).
In the case of soil from field D3, the pH was 6.8 in 2024 and 6.7 in 2025. However, soil pH measurements in field O1 showed a change from 6.9 in 2024 to 7.6 in 2025.
Analysis of the enzymatic activity of soil from field O1 revealed that the values in 2024 were statistically significantly lower than those in 2025 (Figure 2A). The most pronounced difference was observed in alkaline phosphatase activity, which is clearly related to the soil’s pH value.
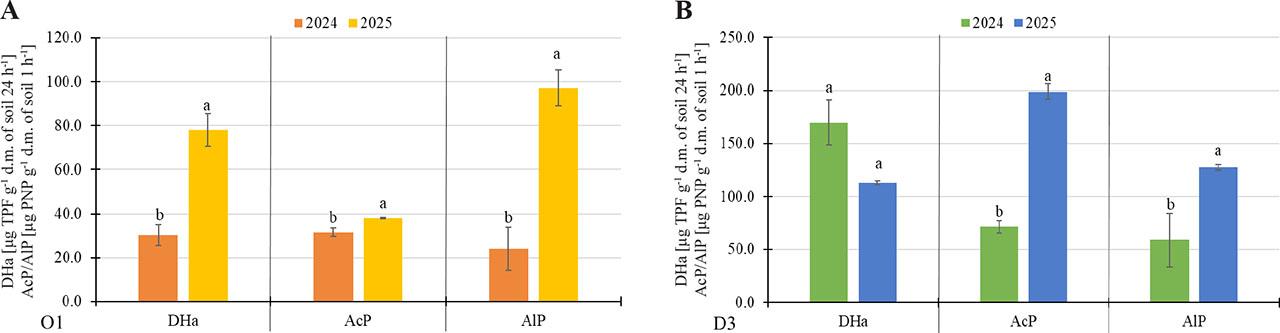
Enzymatic activity in selected soils at two sampling dates – September 2024, and April 2025. (A) Field O1 from the Opole voivodeship; (B) Field D3 from the Lower Silesia voivodeship. DHa – dehydrogenases activity [μg TPF g−1 d.m. of soil 24 h−1]; AcP – acid phosphatase activity [μg PNP g−1 d.m. of soil 1 h−1]; AlP – alkaline phosphatase activity [μg PNP g−1 d.m. of soil 1 h−1]. The values show the mean of n = 3 ± SD. Different letters (a-b) indicate statistically significant differences at P ≤ 0.05 for a given enzyme (One Way ANOVA, Tukey’s HSD test).
Although the difference was not statistically significant, dehydrogenases activity in the soil from field D3 was higher in 2024 (Figure 2B). In contrast, the activity of both phosphatases was significantly higher in 2025.
The metabolic activity of soil microorganisms, as measured by the AWCD index, decreased in both soils in 2025, though the difference was only statistically significant in the case of soil from field O1 (Figure 3). Metabolic diversity (H’ index) in the two analysed soils ranged from 3.30 to 3.36 (see Figure 3). For soil from field D3, the H’ index value was statistically higher in samples collected in 2025.
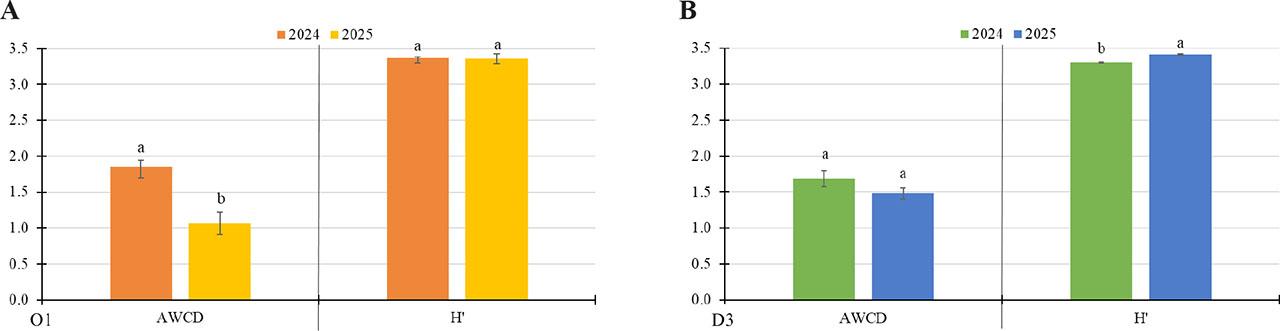
Comparison of metabolic activity (AWCD) and diversity (H’) of soil microorganisms at two sampling dates – September 2024, and April 2025. (A) Field O1 in the Opole voivodeship; (B) Field D3 in the Lower Silesia voivodeship. AWCD – index of metabolic activity; H’ – metabolic diversity index. Values show the mean of n = 3 ± SD. Different letters (a-b) indicate statistically significant differences at P ≤ 0.05 for a given index (One Way ANOVA, Tukey’s HSD test).
As mentioned in the introduction, little research has been conducted on the biological activity of soils after flooding (Furtak et al., 2020b). Flood-affected areas are typically only monitored for sanitary safety by determining the presence of indicator bacteria, such as faecal bacteria (Singh, 2023). Flooding is a complex issue, and studying its impact on soil is challenging. The unpredictability of floods, the variability of floodwaters and the lack of data on pre-flood conditions present major obstacles (Basri et al., 2022; Mishra et al., 2022). As enzymatic activity depends on soil type, organic matter content and land use (Szostek et al., 2022), no attempt was made to compare field O1 with D3. The sole aim of the study was to analyse changes in the values of the biological parameters studied over time in a given soil.
Unfortunately, the studies did not include data from before the flooding, making it difficult to fully understand the changes taking place in the soil. However, analysing data from the period immediately after the flooding (2024) and from a later date (2025) revealed that D3 soil had higher dehydrogenases activity at the first sampling date than seven months later. This is not surprising, as the soil is in an anaerobic state with increased fermentation immediately after a flood, and studies report higher dehydrogenases activity in such conditions, as well as the persistence of increased activity as long as anaerobic conditions persist (Furtak, Gałązka, 2019b; Tate, 1979; Wolińska, 2010). At the same time, this result is consistent with the statistically significantly lower metabolic activity of microorganisms, as expressed by the AWCD index. Previous research conducted using a microcosm experiment revealed a significant increase in dehydrogenase activity during simulated flooding, reaching up to 14-fold after 14 days of water stagnation (Furtak et al., 2019a). Gu et al. (2009) also observed a 90% increase in DHa in flooded soil. In contrast, the dehydrogenases activity measured in soil O1 in 2025 was statistically significantly higher than in 2024. This may be related to the fertilisation applied in spring. According to the survey, mineral, natural (chicken manure) and microbiological fertilisers were used on the field. At the same time, a decrease in the metabolic activity of microorganisms, as measured by the AWCD index, was observed. This may suggest the presence of either (1) “relict” dehydrogenases, originating from dead microorganism cells (Casida, 1977); or (2) a large number of ‘lazy’ microorganisms, which have received ample nutrients through fertilisation and exhibit simple metabolism (Batool et al., 2024; Yadav et al., 2022). It is also worth noting that dehydrogenases activity is a highly sensitive parameter influenced by various factors, such as soil type and organic matter (Bandyopadhyay, Maiti, 2021). However, these factors were not analysed in this pilot study. This prevents accurate tracking of the factors determining dehydrogenase activity in the tested soils.
The activity of phosphatases is related to soil pH, the activity of phosphorus-solubilising microorganisms and the abundance of phosphorus in the soil (Campdelacreu Rocabruna et al., 2024; Rawat et al., 2020; Tian et al., 2021). In soil O1, the activity of both phosphatases was significantly higher in 2025 than in 2024, with alkaline phosphatase activity dominating. This is related to the higher pH of this soil. Additionally, the addition of manure likely increased the pool of organic substrates for phosphatases, resulting in increased activity (Margalef et al., 2017). In soil D3, the activity of both phosphatases was also significantly higher during the second sampling period; however, acid phosphatase dominated in this soil. This is also related to the soil pH and the use of compound mineral fertiliser containing potassium, phosphorus, ammonium nitrogen, and sulphur, which slightly acidifies the soil (Zhang et al., 2022). Another factor contributing to the higher acid phosphatase activity in this sample may be the secretion of this enzyme by soybean roots that had already been sown in the field. Under conditions of limited phosphorus availability, it has been proven that the activity of both intracellular and alkaline phosphatase increases in the root zone of soybean (Zhu et al., 2020). A model experiment showed that phosphatase activity decreases during flooding (Furtak et al., 2019a), which corresponds to the results obtained immediately after the flood in 2024. Studies indicate that the longer the flooding lasts, the greater the inhibition of phosphatase activity, but also of urease and invertase (Gu et al., 2019).
The EcoPlate™ method is commonly used to assess soil microbiomes (Gałązka, Furtak, 2019) and allows the activity of microorganisms to be determined. Analysis of microbial metabolic activity based on the EcoPlate™ method showed that, while there was no significant change in metabolic activity in soil from field D3, microbial diversity increased significantly in 2025. In soil O1, metabolic activity decreased significantly in 2025, but no change in diversity was observed. The increase in the metabolic diversity (H’ index) of microorganisms in soil D3 may be due to a larger pool of various organic compounds in this soil from sediments deposited by floodwater (Bastida et al., 2021), as well as soybean root exudates (Li et al., 2022). Before the flood, wheat was grown in this field. Then, ploughing and mineral fertilisation were carried out, followed by sowing soybeans. These treatments, particularly the change in crop and the introduction of a leguminous species, which increases the nitrogen pool in the soil and secretes a variety of compounds (including organic acids, flavonoids, amino acids, sugars and polyphenols), result in an increase in the metabolic diversity of microorganisms (Badri, Vivanco, 2009; Chen et al., 2008; Huang, 2024; Taylor et al., 2020).
In 2025, there was a significant decrease in the metabolic activity (AWCD index) of soil O1, but no change in the metabolic diversity of microorganisms. This was unexpected given that the soil had been fertilised, crop rotation had been implemented, agrotechnical treatments had been carried out, and a microbiological preparation had been used (Table 1). According to model studies (Furtak et al., 2020b), the AWCD index decreases as a result of flooding. Therefore, low metabolic activity could be expected in both soils sampled in 2024, immediately after the flood subsided. Reasons for the decrease in metabolic activity include soil acidification (Gough, Stahl, 2011), low C and N content, lack of oxygen (Davidson, Janssens, 2006), and various types of pollution (Jacobsen, Hjelmsø, 2014). In the case of the soil under study, however, no such disturbances were observed, making it very difficult to identify the cause. However, the use of chicken manure may have caused the decline in microbial activity. Studies have shown that poultry manure significantly alters the structure of soil microbial communities, decreasing diversity and increasing the abundance of bacteria belonging to the genera Bradyrhizobium, Streptomyces and Pseudomonas, which are characteristic of the analysed manure (Męcik et al., 2023). However, there is also information suggesting that the use of chicken manure increases soil microbiological diversity (Minkina et al., 2023).
The latest literature includes numerous studies analysing plants, their metabolism, and their resistance pathways in situations of hydrological stress (Bispo, Vieira, 2022; Khan et al., 2022; Komatsu et al., 2023, 2024). This is a valuable area of research focusing on how crops respond to flooding. However, these analyses must be linked to analyses of the soil itself, since changes in the soil environment affect plant processes. This suggests that these two areas of research should be combined and analysed together.
This study compared the basic parameters of biological activity in each soil at two sampling dates. Due to the fact that they are different soil types from different locations and crops, no attempt was made to compare the results between soils. Therefore, the impact of flooding on these parameters could be completely different, resulting not from the parameters of the floodwater or the duration of flooding, but from the edaphic properties of the soil (Datt, Singh, 2019; Furtak et al, 2020b). Unfortunately, no results were available for these soils prior to the flood, so it was not possible to make a comparison. This is one of the greatest limitations of post-flood soil research: studies of soils prior to flooding are rarely available, making it difficult to determine precisely what changes are taking place and whether they are reversed once the stress has subsided. Studies have shown that changes in biological activity in flooded soil can be influenced by either soil type (Furtak et al., 2020b), vegetation cover (González Macé et al., 2016), soil organic matter content (Geng et al., 2017), and C/N ratio in soil (Ou et al., 2019). This also highlights the need to examine other parameters, particularly the physical and chemical properties of the soil, to gain a full understanding of the changes occurring.
The results clearly show that the values of the selected biological soil parameters change significantly within a few months of flooding. These changes are influenced by time, the soil’s natural regenerative ability, and human activities such as fertilisation and agrotechnical treatments. It should also be noted that high enzymatic activity does not necessarily indicate high metabolic activity of microorganisms in the soil.
This study is the result of very preliminary and basic research, which should be repeated and continued. Further research is needed to understand these changes, including the long-term impact of flooding on the soil environment and microbiome structure. Estimates suggest that the frequency and severity of floods will increase in the future. It is essential to observe changes in the soil resulting from flooding in order to understand the impact of flooding on soil quality, microorganisms and plant physiology. This is important for mitigating flood stress in sensitive species and ecosystems.