
Rice (Oryza sativa L.) is one of the major cereal crops produced in Pakistan and is cultivated on an area of 2,900,600 hectares with a production of 11,174,700 tons (FAO, 2017). As a staple food, its consumption exceeds 100 kg per capita annually in most of the Asian countries (Seck et al., 2012). Several biotic and abiotic constraints limit the yield and quality of rice. Among biotic constraints, plant-parasitic nematodes are an emerging threat to rice production. Root-knot nematodes (RKNs) are the most destructive plant-parasitic nematodes in upland, lowland, and deep-water rice cultivation systems (Panwar and Rao, 1998; Bridge et al., 2005). RKNs possess the ability to penetrate the roots, induce root galling, suppress plant defense mechanisms, hijack the plant’s metabolic system, and establish giant cells for the sake of their own benefit (Kyndt et al., 2014; Ali et al., 2017a, 2017b, 2018). As an outcome, plants gradually lose vigor, ultimately leading to substantial yield loss (Bridge et al., 2005; Win et al., 2015; Ali et al., 2015). Among various RKNs, Meloidogyne graminicola (Golden and Birchfield) has emerged as the most serious pest of rice (Mantelin et al., 2017). M. graminicola was first reported in Pakistan by Munir and Bridge (2003) during a survey of rice fields of Sheikhupura, Punjab, Pakistan. The subtropical climate of Pakistan and warm sandy soils are favorable for the development and reproduction of RKNs (Khattak, 2008). But plant-parasitic nematodes in these regions have received little attention in the past, and only a few surveys have been conducted to estimate their prevalence and incidence (Kafi, 1963; Ahmad and Khan, 1973; Saeed and Ashrafi, 1973; Khan et al., 2005; Zarina and Shahina, 2010; Anwar and McKenry, 2012). All varieties of rice grown in South Asian and Southeast Asian countries, whether upland or lowland, have exhibited susceptibility to M. graminicola (Padgham et al., 2004; Das et al., 2011; Win et al., 2013). Currently, there is little information about the association of M. graminicola with rice in Pakistan.
Compared with other phytopathogens, nematodes are difficult to control because they attack belowground parts of plants, resulting in reduced growth and yield loss (Williamson and Hussey, 1996). Moreover, nematodes are polyphagous pests that attack over 5,500 plant species, including several economically important crops (Moens and Perry, 2009; Ali et al., 2015; Ali et al., 2019). Successful control of Meloidogyne species can only be achieved by rapid and accurate identification of the nematode. Traditional techniques of nematode identification based on morphological features (Eisenback, 1985) are challenging because they are laborious and require extensive training and expertise. Historically, RKNs have been identified based primarily on morphological features. Currently, there is no information available about the molecular identification of Meloidogyne spp. in Pakistan, especially in regard to M. graminicola.
Several molecular techniques are available for the identification of Meloidogyne species (Blok and Powers, 2009). Among these techniques, the polymerase chain reaction (PCR) is a sensitive, quick, and accurate tool (Niu et al., 2011). Adam et al. (2007) developed a molecular diagnostic key that uses several molecular approaches to identify seven economically important RKNs that are frequently encountered in diagnostic laboratories. Moreover, with the increase in DNA-based sequencing, the tandem repeat unit segments of the 18 S, ITS1, 5.8 S, ITS2, and 28 S regions of the ribosomal DNA array (rDNA) and mitochondrial DNA (mtDNA) have proved to be efficient diagnostic tools for accurately identifying of RKNs (Landa et al., 2008; Naz et al., 2012).
Accurate identification of M. graminicola, as well as its prevalence and distribution spectra, is fundamental for applying management strategies in the field. Therefore, we used morphological and molecular approaches to identify RKNs in order to determine the distribution, prevalence, and disease intensity of M. graminicola in Chiniot and Faisalabad districts of Central Punjab, Pakistan.
A survey of rice-growing areas of Faisalabad and Chiniot districts of central Punjab, Pakistan, was conducted during September 2014, 2015, and 2016. Five root samples of rice plants and 1000-ml composite soil samples were collected from each sampling site, preserved in polythene bags, and labeled. The samples were transported to the Nematology Lab, Department of Plant Pathology, University of Agriculture, Faisalabad, for further processing. The roots were gently washed with distilled water to remove adhering soil and plant debris. Nematode prevalence was determined by using the formula of Norton (1978):
Second-stage juveniles (J2s) of M. graminicola were isolated from rice roots. The root samples from each field were pooled, chopped into pieces, and mixed thoroughly. Three subsamples of 5-g roots were taken, and juveniles were isolated from infected roots using the Baermann funnel method and collected in a beaker. Nematode extraction from soil samples was carried out using Whitehead’s tray method (Whitehead and Hemming, 1965). Soil samples were mixed thoroughly, and a 100-ml composite sample was placed in a plastic bucket and 1–2 liters of water were added. Plant debris, heavy soil particles, and rocks were drained manually. The supernatant was sieved through a 50-mesh-size sieve in a separate bucket. The procedure was repeated by adding 1,000-ml water again and agitated properly. After that, the suspension was left to settle and poured again into another basket using a 100-mesh sieve. The process was repeated twice again using 250- and 325-mesh sieves. The final suspension was transferred to a 500-ml beaker and the supernatant was allowed to settle for one hour. Three subsamples of 2 ml each were taken and examined in a counting dish. Second-stage juveniles (J2) were counted using a stereomicroscope (Olympus SZ2-ILGB). Mature females of M. graminicola were identified based on micrographs of the internal and external perineal patterns (Jepson, 1987). Stained hooked gall tissues were teased apart with a pointed needle to separate mature females. The neck region of individual nematodes was excised, and the posterior part was dipped in 45% lactic acid solution to remove body tissues. The perineal patterns were trimmed and transferred on a transparent glass slide in a glycerin drop. External views of the perineal patterns were visualized using scanning electron microscopy (SEM) at Brigham Young University, Provo Utah, USA. Accordingly, specimens were dehydrated via ascending ethanol series prior to critical point drying. The animals were then mounted on stubs and gold-sputtered. Micrographs were recorded via Helios Nanolab 600 SEM (Thermo-Fisher Scientific, Hillsborough, USA).
Data were subjected to statistical analysis using Statistix (Ver. 8.1). The experimental design for the analysis of galling index, juveniles/soil sample, juveniles/root sample, stylet, and non-stylet-bearing nematodes was by a completely randomized block design with treatment means separated using a LSD test at P = 0.05.
Isolates derived from single egg masses were used to extract DNA. Twenty larvae were picked, rinsed thrice with sterile distilled water, transferred to a microcentrifuge tube, and crushed in 500 µL of SDS extraction buffer containing Tris–HCl (1 M, pH 7.5), EDTA (0.5 M, pH 8.0), SDS (10% w/v), and dd H2O with a grinding plastic stick (Mitkowski et al., 2003). Proteins in the solution were digested by adding 20 µL proteinase K, 100 µg ml−1. Later, 500 µL of phenol (25:24:1, phenol/chloroform/isoamyl alcohol) was added and vortexed briefly. The tube was centrifuged at 14,000 rpm for 5 min and the supernatant was collected and transferred to another sterile microcentrifuge tube. Sodium acetate (3 M), 50 µL and ice-cold isopropanol, 500 µL, were added to the supernatant, vortexed gently, and centrifuged at 14,000 rpm for 5 min. The supernatant was discarded to save the pellet. Five-hundred µL of ethanol (96%) was added to the pellet and centrifuged for 3 min at 13,000 rpm. The supernatant was discarded again, and the pellet air-dried in a laminar flow chamber for 2 hr. DNA was dissolved in 30 µL of TE buffer (1 ml of 1 M Tris base (pH 8.0) and 0.2 ml of EDTA (0.5 M), dd H2O) and stored at –20°C until required for PCR reactions.
The PCR reaction mixture was prepared by using 10 µL of 10X PCR buffer, 2-µL dNTPs (10 mM), 2-µL forward primer (10 µM), 2-µL reverse primer (10 µM), 2-µL template DNA (> 50 ng), 1-µL Taq polymerase, and 81-µL ddH2O, for a total reaction volume of 100 µL. PCR reactions were carried out in a thermal cycler (BIO-RAD T100TM) under the following conditions: (1) predenaturation at 94 °C for 5 min, (2) denaturation at 94°C for 1 min, (3) annealing at 64°C for 1 min, (4) extension at 72°C for 1 min, (5) 35 cycles of this process, and (6) final extension at 72°C for 5 min. The sequences of primers used for amplification of the internal transcribed spacer region are 1) forward primer rDNA2 (5’–TTGATTACGTCCCTGCCCTTT–3’) (Vrain et al., 1992) and reverse primer rDNA1. 5.8 s (5’–ACGAGCCCGAGTGATCCACCG–3’) (Cherry et al., 1997). Primers were synthesized by Sigma Genosys Inc, St. Louis, MO, USA.
The amplified PCR product was viewed on a gel stained with ethidium bromide under a UV transilluminator. DNA was excised from the gel and purified for sequencing using quick Gel Extraction Kit (Qiagen Gel Extraction Kit, Qiagen, Hilden, Germany). Purified DNA was sent for sequencing to L.G.C. Genomics, Germany, or the DNA sequencing center at Brigham Young University, Provo, Utah, USA.
Due to the high degree of interspecific variation in nucleotide sequences of nematodes (Hu and Gasser, 2006), the deduced internal transcribed spacer region sequences were used for phylogenetic study. The ITS sequences of the eight isolates were used to query the most similar curated sequences on GenBank by performing open-nucleotide BLAST (Basic Local Alignment Search Tool, Camacho et al., 2009). The nearest matches as well as those from closely related species of Meloidogyne (McClure et al., 2012) were downloaded from GenBank and used for subsequent phylogenetic analysis. Multiple-sequence alignment was generated using MUSCLE (Edgar, 2004). The alignment settings included optimization of profile-dependent parameters. Sequences were first grouped by similarity with anchor optimization. Iteration 1 = kmer4_6 with pctid_kimura for subsequent interactions using the UPMGB clustering method and pseudo-tree rooting. CLUSTALW was used for the distance weighting scheme, with 32-base anchor spacing and 24-base minimum diagonals. Bayesian analyses were carried out using MrBayes 3.2.6 (Huelsenbeck and Ronquist, 2001). A general time-reversible model of sequence evolution with four categories of gamma rate variation was chosen as the optimal model of sequence evolution as per Posada and Crandall (1998). Bayesian analysis was initiated with a random starting tree with four heated chains of chain length 1,100,000, burn-in length of 100,000, and subsampling frequency of 200. Branch lengths were unconstrained.
The field survey data during the rice cropping season of 2014, 2015, and 2016 indicated that both districts Chiniot and Faisalabad have M. graminicola infestation during all the seasons. The GPS coordinates for each sample were used to plot the presence and absence of M. graminicola at the sampled sites in Faisalabad and Chiniot (Figure 1). The prevalence of M. graminicola at forty surveyed locations of Chiniot and Faisalabad is given in Tables 1 and 2. Maximum prevalence of M. graminicola (22.5 and 27.5%) was observed in Chiniot and Faisalabad, respectively, during the rice-growing season of 2016, while the minimum prevalence (17.5 and 20%, respectively) was recorded in Chiniot and Faisalabad during the cropping season of 2014. During 2014 to 2016 at Chiniot among 40 surveyed locations, the presence of M. graminicola was observed at 7, 8, and 9 locations respectively. Similarly, 40 locations were surveyed in Faisalabad during 2014–2016, and M. graminicola prevalence was recorded at 8, 10, and 11 locations, respectively. M. graminicola showed typical hook-shaped galls on rice roots that are a characteristic feature of M. graminicola infection (Figure 2).
Prevalence of M. graminicola in Chiniot district.
| Prevalence | |||||||
|---|---|---|---|---|---|---|---|
| Sr # | Locations | Elevation | Latitude | Longitude | 2014 | 2015 | 2016 |
| 1 | 133 J.B. | 178 V | 31.57931 | 072.97683 | − | − | − |
| 2 | 134 J.B. | 173 V | 31.59705 | 072.96338 | + | + | + |
| 3 | 223 J.B. | 169 V | 31.62043 | 072.96727 | − | − | − |
| 4 | Moza Rajoya | 176 R | 31.66198 | 072.97448 | + | + | + |
| 5 | M.Thattian | 184 R | 31.68778 | 072.97489 | − | − | − |
| 6 | M. Johdpur | 180 R | 31.69026 | 072.97472 | − | − | − |
| 7 | Ahmad Nagar | 182 R | 31.77321 | 072.89966 | − | − | − |
| 8 | Ahmad Pur | 187 R | 31.79090 | 072.87729 | − | − | − |
| 9 | M. kanwewala | 183 R | 31.81763 | 072.82544 | − | − | − |
| 10 | Moza Judhi | 186 R | 31.82471 | 072.81122 | − | − | − |
| 11 | 202 J.B. | 192 V | 31.84161 | 072.83868 | − | − | − |
| 12 | Kote Qazipur | 190 R | 31.84774 | 072.84970 | − | − | − |
| 13 | Biewal | 187 R | 31.90920 | 072.94682 | − | − | − |
| 14 | Kalowal | 182 R | 31.90423 | 072.95624 | − | − | − |
| 15 | Jand wala | 185 R | 31.88589 | 072.99015 | − | − | − |
| 16 | Pungu Morr | 184 R | 31.84625 | 072.96008 | + | + | + |
| 17 | Moza Johdabad | 183 R | 31.83006 | 072.93907 | − | − | − |
| 18 | M.Hussainabad | 188 R | 31.82157 | 072.92782 | + | + | + |
| 19 | M.Mohsanabad | 178 R | 31.79400 | 072.89456 | − | − | − |
| 20 | Moza Vaisaan | 176 R | 31.78678 | 072.88346 | − | − | − |
| 21 | 241 J.B. | 178 V | 31.72044 | 072.95988 | − | − | − |
| 22 | Moza Bukharian | 177 R | 31.70576 | 072.95381 | − | − | − |
| 23 | Abbas nagar | 180 R | 31.68935 | 72.93636 | − | − | − |
| 24 | Kandiwal | 177 R | 31.67671 | 72.90760 | − | − | − |
| 25 | 241 J.B. | 179 V | 31.66678 | 72.87928 | − | − | − |
| 26 | Thatta Jahania | 177 R | 31.66310 | 72.86660 | + | + | + |
| 27 | Adil wala bangla | 169 V | 31.66029 | 72.85530 | − | − | − |
| 28 | 194 J.B. | 171 V | 31.65541 | 72.83943 | − | − | − |
| 29 | 94 J.B. | 179 V | 31.64864 | 72.82515 | − | − | − |
| 30 | 202 J.B. | 179 V | 31.62264 | 72.77077 | + | + | + |
| 31 | Moza Nitthar | 167 R | 31.59611 | 72.70949 | − | − | − |
| 32 | 254 J.B. | 168 V | 31.58803 | 72.69275 | − | − | − |
| 33 | Jamya abad | 168 T | 31.55409 | 72.64627 | + | + | + |
| 34 | Gondlan wali | 167 R | 31.53430 | 72.66215 | − | − | − |
| 35 | Chenab mills Ltd | 166 C | 31.51384 | 72.67929 | − | − | − |
| 36 | Bhawana city | 170 C | 31.50520 | 72.68622 | − | − | − |
| 37 | 129 J.B. | 172 V | 31.50039 | 72.69027 | − | − | − |
| 38 | 41 J.B. | 165 V | 31.49171 | 72.69737 | − | + | + |
| 39 | 223 J.B. | 169 V | 31.45835 | 72.72484 | − | − | + |
| 40 | Rahmuana | 181 V | 31.42409 | 72.75250 | − | − | − |
Note: Villages with official code (V), JB, stand for canal irrigation official code. R stands for River Chenab basin areas and C stands for City/ Town area. Positive (+) sign indicates presence of RKN; negative (−) indicates absence of RKN in the rice field.
Prevalence of M. graminicola in district Faisalabad.
| Prevalence | |||||||
|---|---|---|---|---|---|---|---|
| Sr # | Locations | Elevation | Latitude | Longitude | 2014 | 2015 | 2016 |
| 1 | 217 R.B. | 175 V | 31.44325 | 073.00077 | − | − | − |
| 2 | 61 J.B. | 176 V | 31.45346 | 072.97887 | − | − | − |
| 3 | 57 J.B. | 177 V | 31.45328 | 072.98147 | − | − | − |
| 4 | 58 J.B. | 186 V | 31.46937 | 072.99122 | + | + | + |
| 5 | 52 J.B. | 181 V | 31.49570 | 073.00120 | − | − | − |
| 6 | 54 J.B. | 186 V | 31.500394 | 072.99582 | − | − | − |
| 7 | 51 J.B. | 180 V | 31.51203 | 072.98117 | − | − | − |
| 8 | 53 J.B. | 180 V | 31.52299 | 072.97618 | − | − | − |
| 9 | 49 J.B. | 186 V | 31.56101 | 072.98678 | − | − | − |
| 10 | 218 J.B. | 173 V | 31.43074 | 072.97399 | + | + | + |
| 11 | 64 J.B. | 173 V | 31.39198 | 072.93964 | − | − | − |
| 12 | 68 J.B. | 174 V | 31.37310 | 072.92706 | − | − | − |
| 13 | 66 J.B. | 178 V | 31.40154 | 72.97310 | − | − | − |
| 14 | 275 J.B. | 169 V | 31.34735 | 72.83235 | − | − | − |
| 15 | 70 J.B. | 181 V | 31.36304 | 72.90952 | − | + | + |
| 16 | 289 R.B. | 172 V | 31.33934 | 072.99238 | − | − | − |
| 17 | 245 R.B. | 177 V | 31.33586 | 073.01232 | − | − | − |
| 18 | 296 R.B. | 179 V | 31.33620 | 073.04997 | − | − | − |
| 19 | 254 R.B. | 166 V | 31.33576 | 073.00168 | − | − | − |
| 20 | 269 R.B. | 167 V | 31.19880 | 072.99097 | − | − | − |
| 21 | 135 G.B. | 169 V | 31.14848 | 072.98298 | − | − | − |
| 22 | UAF | 179 C | 31.43735 | 073.07427 | + | + | + |
| 23 | Samundari City | 168 C | 31.05557 | 072.97504 | − | − | − |
| 24 | 71 G.B. | 165 V | 31.05584 | 072.97427 | − | − | + |
| 25 | 73 G.B. | 165 V | 31.05005 | 072.99210 | − | − | − |
| 26 | 442 G.B. | 172 V | 31.04441 | 073.00982 | + | + | + |
| 27 | 393 G.B. | 172 V | 31.02308 | 073.07303 | − | − | − |
| 28 | 423 G.B. | 178 V | 31.07426 | 073.13823 | − | − | − |
| 29 | 424 G.B. | 172 V | 31.08084 | 073.14034 | + | + | + |
| 30 | 430 G.B. | 171 V | 31.11562 | 073.15054 | − | − | − |
| 31 | 172 G.B. | 178 V | 31.14319 | 073.14831 | − | − | − |
| 32 | 171 G.B. | 178 V | 31.16456 | 073.13882 | − | − | − |
| 33 | 39 G.B. | 176 V | 31.20585 | 073.17356 | + | + | + |
| 34 | 41 G.B. | 182 V | 31.20572 | 073.17418 | − | − | − |
| 35 | 54 G.B. | 183 V | 31.26873 | 073.29616 | − | − | − |
| 36 | 21 G.B. | 191 V | 31.30716 | 073.38556 | − | − | − |
| 37 | 205 R.B. | 186 V | 31.43253 | 073.23050 | + | + | |
| 38 | 66 G.B. | 181 V | 31.34015 | 073.33852 | + | + | + |
| 39 | 216 R.B. | 183 V | 31.38091 | 073.22599 | − | − | − |
| 40 | 229 R.B. | 184 V | 31.39439 | 073.20733 | + | + | + |
Note: Villages with official code (V), JB, RB and GB stand for canal irrigation official code. C indicates a City/ Town area. Positive (+) sign indicates presence of RKN and negative (−) indicates absence of RKN in the rice field.
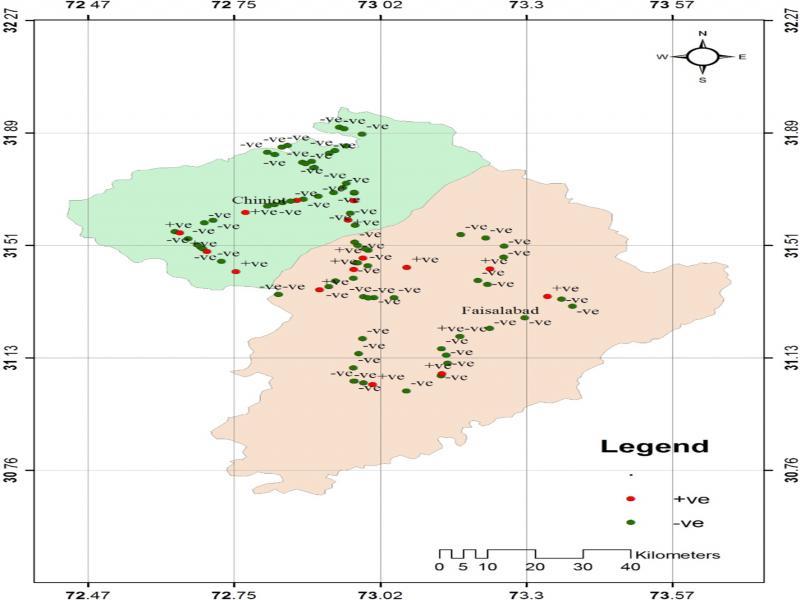
GPS map of Faisalabad and Chiniot districts. Red dots indicate locations of samples infested with M. graminicola.

Rice roots infected with M. graminicola. The roots are showing typical symptoms of galls or knots.
The severity of M. graminicola attack on rice roots in infested fields was based on different attributes like galling index, juveniles per root system, juveniles per 100 mL of soil, number of stylet-bearing nematodes (SBN), and the number non-stylet-bearing nematodes (NSBN) from both of the surveyed districts (Tables 3 and 4). The highest galling index was observed in Chiniot and Faisalabad during 2016 and 2014, respectively. M. graminicola attack was observed at both districts with significantly high galling index indicative of high infestation of M. graminicola. In Chiniot, the highest juvenile/root sample population was recorded in 2016. The highest population of juveniles/soil sample and non-stylet-bearing nematodes was observed in 2014, while the maximum recorded number of stylet-bearing nematodes was from Chiniot in 2015. During 2016, the highest population of juveniles/root sample and stylet-bearing nematodes was observed in Faisalabad, while the highest number of juveniles/soil sample and non-stylet-bearing nematodes was recorded during 2015.
The infection of M. graminicola was recorded on eleven alternate hosts. All examined alternate hosts showed varying degrees of M. graminicola infestation (Table 5). Among them, the maximum juveniles/root sample, juveniles/soil sample, number of stylet-bearing nematodes, and number of non-stylet-bearing nematodes were recovered from Echinochloa crusgalli. The lowest number of juveniles/root sample, juveniles/soil sample, and non-stylet-bearing nematodes were observed in Brassica oleracea. The lowest number of stylet-bearing nematodes was recorded in Trigonella foenum-graecum. Most of the infested fields were canal-irrigated and a few were tube-well irrigated. Both types of irrigation systems generally favor nematode attack, but nematode populations were higher for in-canal irrigation than tube-well. Most of the farmers have a wheat-rice cropping system and only a few of them use a rice-vegetable cropping system (data not shown). In cropping systems that include vegetables, nematode infection was higher on alternate hosts. Grasses like Cyperus rotundus, Dactylocteniuma egyptium, Echinochloa crusgalli, Eclipta alba, and Paspalum distichum were reported as alternate hosts for rice-vegetable cropping systems, while Avena fatua, Phalaris minor, and Rumex dentatus were recorded as alternate hosts in the rice-wheat cropping system.
The morphological examination of perineal patterns among obtained isolates showed that they were oval to circular shaped, dorsoventral, moderate in height of the arc, and with no lateral incisures or gaps. The tail tip showed prominent, coarse, and well-separated striae. The obtained perineal patterns were similar to previously described patterns for M. graminicola (Figure 3).
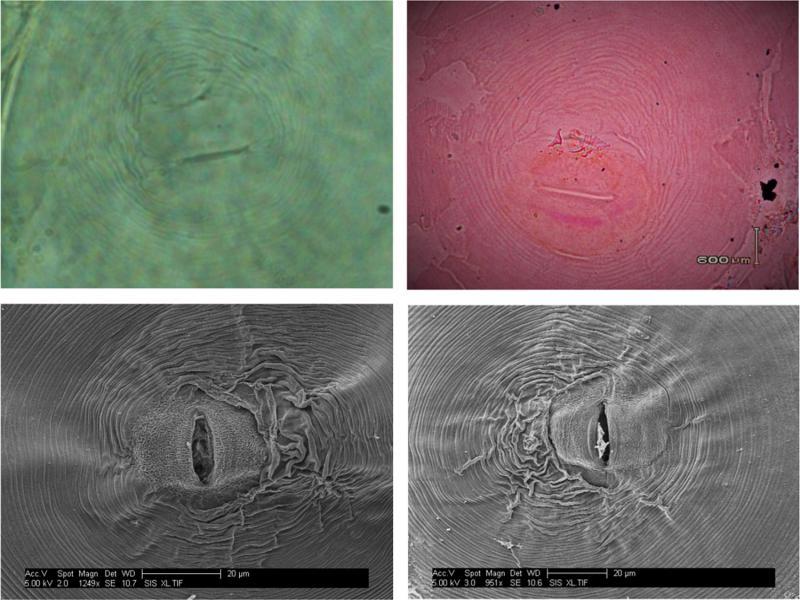
Perineal pattern of M. graminicola by light (above) and electron (below) microscopy.
PCR amplification and sequencing yielded eight unique sequences. The isolates gave similar-sized bands of 326–328 bp on a gel stained with ethidium bromide (Figure 4). GenBank accession numbers for each of the obtained sequences are given in Tables 3–6. M. graminicola isolates JB1CT, JB2 UAF, JB3FSD1, JB3FSD2, JB3FSD3, JB3FSD4, JB3FSD5, and JB3FSD6 (accessions KX757064, 326 bp, KX757065, 328 bp, KX757066, 326 bp, KX757067, 326 bp, MH057345, 326 bp, MH057346, bp, MH057347, bp, and MH057348, 326 bp, respectively) showed 98–100% similarity with GenBank accession numbers KF250491, DQ909030, KF250491, MG773553, KF250481, KJ572383, JF949754, and DQ909040, which were previously curated as M. graminicola. The multiple-sequence alignment of the eight isolates demonstrates that there were very few single-nucleotide polymorphisms (SNPs) between the ITS sequences of these isolates (Figure 5). When trimmed for phylogenetic analysis, JB3FSD1 was no longer unique and was removed from subsequent analyses.
Incidence of M. graminicola at different locations of district Chiniot.
| Year | Location | Galling index | Juveniles/root sample | Juveniles/soil sample | Stylet bearing nematodes | Non-stylet bearing nematodes |
|---|---|---|---|---|---|---|
| 2014 | 134 J.B | 2.20 b | 300.80 b | 140.40 d | 4.20 bc | 2.80 e |
| Moza Rajoya | 3.80 a | 348.60 ab | 280.80 b | 3.80 c | 9.40 cd | |
| Pungu Mor | 2.60 ab | 290.40 b | 108.80 d | 1.60 d | 11.40 bcd | |
| Jamya Abad | 3.20 ab | 390.40 a | 119.00 d | 3.80 c | 12.00 bc | |
| Jandwala | 4.00 a | 297.20 b | 211.40 c | 5.20 ab | 8.40 d | |
| 223 JB-2 | 3.80 a | 336.60 ab | 347.20 a | 5.80 a | 14.00 ab | |
| Rahmuana | 3.20 ab | 294.00 b | 184.00 c | 5.40 ab | 17.20 a | |
| 2015 | 134 J.B | 3.40 a | 172.20 a | 117.80 f | 6.20 cd | 7.20 d |
| Moza Rajoya | 3.20 a | 117.40 c | 242.60 c | 2.80 f | 9.20 cd | |
| Pungu Mor | 2.80 a | 163.20 ab | 136.40 e | 7.40 bc | 13.00 a | |
| Jamya Abad | 3.20 a | 126.40 bc | 135.80 e | 9.20 a | 10.00 bc | |
| Jandwala | 2.80 a | 156.80 abc | 107.00 g | 5.80 de | 10.40 bc | |
| 223 JB-2 | 3.20 a | 134.00 abc | 194.60 d | 4.80 e | 9.20 cd | |
| Rahmuana | 3.60 a | 130.80 abc | 279.60 b | 7.80 b | 11.80 ab | |
| 202 JB | 3.40 a | 131.80 abc | 292.20 a | 7.40 bc | 10.40 bc | |
| 2016 | 134 J.B | 3.40 ab | 278.00 bc | 218.00 b | 5.20 c | 4.00 c |
| Moza Rajoya | 3.20 ab | 268.60 c | 298.80 a | 9.00 a | 10.20 ab | |
| Pungu Mor | 2.80 b | 332.80 abc | 185.20 e | 8.20 ab | 7.20 bc | |
| Jamya Abad | 3.00 b | 349.40 abc | 213.80 bc | 7.40 b | 9.40 ab | |
| Jandwala | 3.40 ab | 380.80 a | 199.20 d | 5.80 c | 8.80 b | |
| 223 JB-2 | 4.20 a | 292.60 bc | 172.60 f | 5.20 c | 9.00 b | |
| Rahmuana | 3.80 ab | 280.80 bc | 211.40 bc | 5.60 c | 6.80 bc | |
| 202 JB | 3.80 ab | 362.00 ab | 204.80 cd | 5.00 c | 8.00 b | |
| 129 JB | 2.80 b | 330.60 abc | 187.20 e | 7.20 b | 12.60 a |
Note: The means followed by the same letters in a column are not significantly different by LSD test (c level).
Incidence of M. graminicola at different locations of Faisalabad district.
| Year | Location | Galling index | Juveniles/root sample | Juveniles/soil sample | Stylet bearing nematodes | Non-stylet bearing nematodes |
|---|---|---|---|---|---|---|
| 2014 | 58 J.B. | 2.60 b | 334.80 ab | 157.20 e | 5.60 bc | 9.00 c |
| 218 R.B. | 3.60 ab | 352.20 a | 131.60 g | 4.80 cd | 8.80 c | |
| 70 J.B. | 1.40 c | 340.00 a | 219.80 b | 4.80 cd | 11.00 ab | |
| 442 G.B. | 3.60 ab | 332.40 ab | 147.60 f | 4.80 cd | 10.60 abc | |
| 39 G.B. | 3.00 b | 266.20 b | 238.00 a | 6.00 abc | 10.00 bc | |
| 424 G.B. | 2.60 b | 352.60 a | 172.20 d | 6.40 ab | 9.20 bc | |
| 205 R.B. | 3.20 ab | 320.00 ab | 184.00 c | 7.00 a | 12.00 a | |
| UAF | 4.20 a | 280.80 ab | 113.60 h | 4.20 d | 10.40 abc | |
| 2015 | 58 J.B. | 2.60 ab | 336.00 a | 205.60 b | 5.40 cd | 9.20 c |
| 218 R.B. | 2.20 b | 342.20 a | 130.20 e | 5.20 cd | 8.80 c | |
| 70 J.B. | 3.00 ab | 331.00 a | 137.40 e | 4.20 d | 9.80 bc | |
| 442 G.B. | 2.40 ab | 338.40 a | 148.80 de | 5.20 cd | 9.80 bc | |
| 39 G.B. | 3.20 ab | 276.20 a | 185.00 bc | 7.80 a | 10.00 bc | |
| 424 G.B. | 3.80 a | 339.60 a | 128.60 e | 4.80 cd | 9.20 c | |
| 205 R.B. | 2.20 b | 330.60 a | 308.40 a | 7.00 ab | 10.60 abc | |
| UAF | 3.40 ab | 292.60 a | 127.80 e | 5.80 bc | 9.40 bc | |
| 209 R.B. | 2.80 ab | 349.40 a | 172.60 cd | 8.20 a | 11.80 ab | |
| 66 G.B. | 2.60 ab | 352.80 a | 100.40 f | 7.20 a | 13.00 a | |
| 2016 | 58 J.B. | 2.00 d | 361.20 a | 106.80 g | 5.20 de | 9.00 bc |
| 218 R.B. | 2.40 cd | 340.00 ab | 90.60 h | 4.80 de | 9.60 abc | |
| 70 J.B. | 4.00 a | 340.00 ab | 190.80 b | 5.20 de | 10.40 abc | |
| 442 G.B. | 2.20 cd | 328.40 ab | 168.00 c | 9.00 a | 10.20 abc | |
| 39 G.B. | 3.20 bc | 277.80 b | 209.60 a | 5.00 de | 9.80 abc | |
| 424 G.B. | 3.40 ab | 394.00 a | 158.80 d | 7.20 abc | 8.80 bc | |
| 205 R.B. | 2.40 bcd | 362.00 a | 150.60 e | 4.20 de | 8.40 c | |
| UAF | 3.20 abc | 380.80 a | 207.00 a | 6.00 bcd | 9.40 abc | |
| 209 R.B. | 2.80 bcd | 360.40 a | 120.20 f | 5.40 cde | 9.20 abc | |
| 66 G.B. | 3.40 ab | 368.60 a | 162.60 d | 7.60 ab | 10.80 ab | |
| 71 G.B. | 2.60 bcd | 358.00 a | 123.20 f | 3.60 e | 11.20 a |
Note: The means followed by the same letters in a column are not significantly different by LSD test (P = 0.05 level).
Incidence of M. graminicola on different alternate hosts.
| Sr # | Local name | Botanical name | Juveniles /root sample | Juveniles/soil sample | Stylet bearing nematodes | Non-stylet bearing nematodes |
|---|---|---|---|---|---|---|
| 1 | Della | Cyperus rotundus | 826.6 bc | 145.6 b | 11.2 de | 6.6 b |
| 2 | Madhana Grass | Dactyloctenium aegyptium | 700.8 cd | 100.0 de | 11.2 de | 4.4 cd |
| 3 | Barnyard grass | Echinochloa crusgalli, | 4914.6 a | 424.0 a | 29.6 a | 9.4 a |
| 4 | False Daisy | Eclipta alba | 646.0 d | 100.6 de | 22.4 b | 6.4 bc |
| 5 | Knotgrass | Paspalum distchum | 494.2 e | 91.2 e | 14.6 cd | 5.4 bcd |
| 6 | Toothed Dock | Rumex dentatus | 783.8 cd | 121.0 cd | 23.0 b | 4.6 bcd |
| 7 | Jangli gai | Avena fatua, | 776.2 cd | 101.6 de | 18.8 bc | 5.0 bcd |
| 8 | Dumbi Sitti | Phalaris minor | 942.8 b | 129.4 bc | 8.4 e | 4.0 d |
| 9 | Cauliflower | Brassica oleracea | 142.2 f | 43.4 g | 10.6 de | 3.4 d |
| 10 | Coriander | Coriandrum sativum L. | 143.8 f | 85.4 ef | 10.2 de | 3.6 d |
| 11 | Maithi | Trigonella foenum-graecum L. | 178.6 f | 64.4 fg | 7.6 e | 5.2 bcd |
Note: The means followed by the same letters in a column are not significantly different by LS D test (p = 0.05 level).
ITS sequences of M. graminicola isolates with GenBank accession numbers.
| Location | Elevation | Latitude | Altitude | Isolate | GenBank Accession | Nucleotide |
|---|---|---|---|---|---|---|
| Rajoya | 176 | 31.66198 | 072.97448 | JB1CT | KX757064.1 | 326 bp |
| UAF | 179 | 31.43735 | 073.07427 | JB2 UAF | KX757065.1 | 328 bp |
| 58 J.B | 186 | 31.46937 | 072.99122 | JB3FSD1 | KX757066.1 | 326 bp |
| 218 J.B | 173 V | 31.43074 | 072.97399 | JB3FSD2 | KX757067.1 | 326 bp |
| 442 G.B | 172 V | 31.04441 | 073.00982 | JB3FSD3 | MH057345.1 | 326 bp |
| 424 G.B | 172 V | 31.08084 | 073.14034 | JB3FSD4 | MH057346.1 | 326 bp |
| 205 R.B | 186 V | 31.43253 | 073.23050 | JB3FSD5 | MH057347.1 | 326 bp |
| 70 J.B | 181 V | 31.36304 | 72.909520 | JB3FSD6 | MH057348.1 | 326 bp |
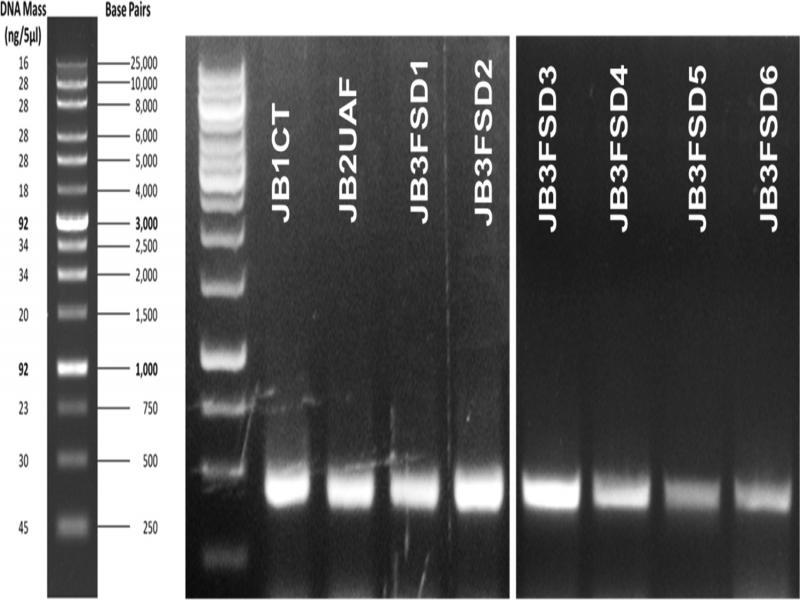
ITS rDNA PCR amplification products using forward primer rDNA2 and reverse primer rDNA1.58 s. Isolate names are given in white text on the upper side of their respective band in the gel; DNA ladder is to the left of the gel.

Multiple alignment of ITS sequences from different isolates collected in Faisalabad and Chiniot. The NCBI Genbank accession numbers and isolate names are given in the start of sequences. Complete bars at the top of the sequences show the degree of conservation of different nucleotides in the ITS sequence among different isolates. Similarly, base conservation is also denoted by the capitalized nucleotide alphabet.
The Bayesian solution is presented in Figure 6. Bayesian posterior probabilities, represented as a percentage, are mapped at nodes where support is greater than 50%. The resulting phylogeny shows that six of the isolates sequenced in this study form a monophyletic clade with the other Asian isolates, including those from India, Nepal, China, and Vietnam. Isolate KX757067 belongs to an unresolved polytomy that contains isolates from Asia as well as Europe and South America. The relationships among the isolates are poorly resolved due to lack of synapomorphies. Three isolates from Pakistan, KX757067, MH057348, and MH057345, have several unique nucleotide substitutions, and thus longer branch lengths, relative to the other isolates of M. graminicola (Figure 6).
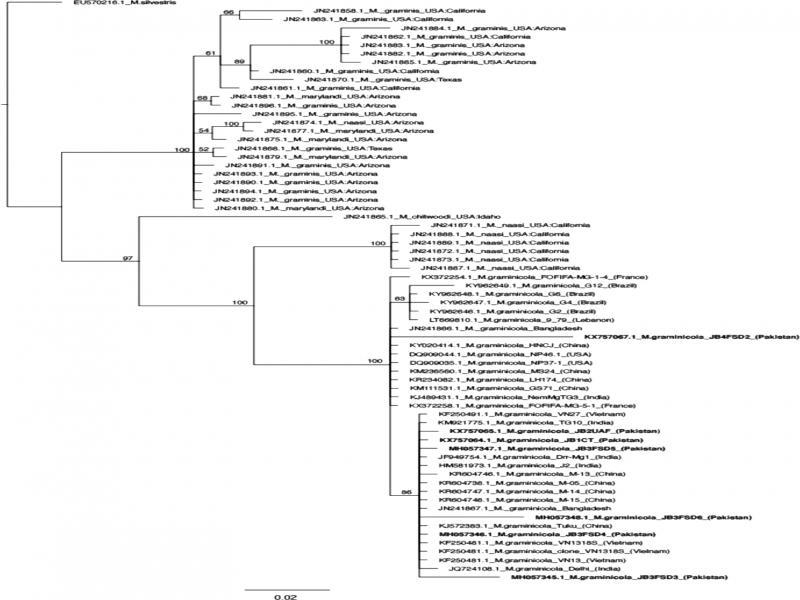
Phylogenetic tree of M. graminicola isolates based on ITS sequences. Sequences from this study are in bold. Taxon labels are Genbank accession numbers followed by species epithet, isolate code, and geographic location of the sequenced isolates. Scale bar represents 2% sequence divergence.
Chiniot and Faisalabad are agriculturally important and are considered some of the most fertile districts in Central Punjab. Rice is also cultivated in these districts with rice-wheat cropping systems. The results of this study reveal variation in the prevalence and infestation rate of M. graminicola in Faisalabad and Chiniot rice fields. Several reports have documented the prevalence of M. graminicola in rice-cropping systems in different countries (Pokharel et al., 2007; Win et al., 2011; Anil et al., 2011; Pascual et al., 2014). M. graminicola has been predominantly reported from lowland production conditions of rice that are common in Chiniot and Faisalabad districts (Pokharel et al., 2007). Most of the rice fields in the present study were infested with M. graminicola that is not common in Punjab, Pakistan, because previously M. incognita has been reported as the predominant RKN prevailing in agroecosystems of Pakistan, which primarily attacks vegetable crops (Anwar et al., 2012). Faisalabad and Chiniot districts have sandy loam soil, a semiarid environment, and are located in the central region of Punjab, Pakistan. The geographic distribution of RKNs depends on environmental factors such as moisture, soil type, and temperature (Sasser and Triantaphyllou, 1977). Nematode abundance and distribution are directly influenced by soil properties and type of irrigation (Nielsen et al., 2014). Sandy soils generally show higher penetration and development of Meloidogyne spp. (Ogbuji, 2004). Soils with a higher percentage of sand had higher abundances of M. incognita (Lawrence et al., 1997). Therefore, sandy loam soil of the surveyed regions could be considered important for enhanced development and penetration of M. graminicola. RKNs are poikilothermic in nature and require elevated temperatures to increase their rates of development on different cropping systems (Van der Waals et al., 2013). The subtropical climate of Chiniot and Faisalabad districts favors the development of M. graminicola in rice-cropping systems.
Meloidogyne species have an extensive host range, including grasses, weeds, field, and vegetable crops (Sasser and Freckman, 1987). During our survey, seven alternate hosts of M. graminicola were also recorded. Plants were classified as alternate hosts based on prevalence, galling index, RKNs per g root, and RKNs per g soil. Previously, we have reported these plant species as alternate hosts of M. graminicola in Pakistan (Jabbar et al., 2016). These alternate hosts help nematodes to persist through summer and winter and act as an important reservoir of nematodes (Queneherve et al., 1995).
Morphological identification based on perineal patterns has been the standard criteria used to identify Meloidogyne species since 1949 (Chitwood, 1949). Based on light and scanning electron microscopy, the perineal pattern of isolates from Pakistan is similar to previously described patterns of M. graminicola (Yik and Birchfield, 1978; Bernard and Eisenback, 1997) with slight variation, overlapping with the patterns of M. trifoliophila and M. oryzae. However, perineal patterns alone are insufficient to confirm the identity of Meloidogyne spp. as perineal patterns for M. graminicola, M. oryzae, and M. trifoliophila may be conflated (Maas et al., 1978; Pokharel et al., 2007).
To the best of our knowledge, this is the first report of ITS sequences of M. graminicola from Pakistan. Sequence similarity analysis was concordant with morphological analyses, and phylogenetic analysis confirmed that our isolates nested well within the other M. graminicola sequences available in GenBank, confirming that they are conspecific.
Several studies have demonstrated that ITS sequencing is not only a useful diagnostic approach for Meloidogyne, Globodera, Heterodera, Longidorus, Xiphinema, and Pratylenchus spp. (Powers, 2004), but also for phylogenetic analysis of a number of species in Heterodera, Meloidogyne, and Bursaphelenchus (Hugall et al., 1999; Powers, 2004). Variability in ITS sequences among the isolates of Pakistan was also observed based on SNPs. The variability in the ITS sequences indicated that amplified ITS regions of Meloidogyne species might be useful for population studies. The variability in ITS sequences within an isolate observed in this study could be due to the presence of multiple alleles and/or multiple copies of the sequences that are reported previously in other RKN species (Powers, 2004). Indeed, the high degree of genetic diversity among the Pakistani isolates relative to other regions suggests Pakistan as a possible ancestral area for the Asian isolates of this species (Figure 6).
Our results confirmed that all Pakistani sequenced isolates of root-knot nematode collected from rice were M. graminicola. The populations are quite morphologically homogeneous, with only slight variations in morphometric characters and virulence. Our results also confirm the utility of ITS sequences to differentiate M. graminicola from other common species of RKNs and reveal considerable variation among Pakistani isolates and relative isolates from other parts of Asia. Our findings confirm the need for further studies on M. graminicola biology, genetics, and management.
The rice-growing fields of Chiniot and Faisalabad, Central Punjab, Pakistan are infested with M. graminicola. The subtropical climate, monoculture, high cropping intensity, and sandy loam soils of the areas surveyed likely contribute to the widespread prevalence of M. graminicola in Pakistan. M. graminicola from our survey displayed infestation on seven alternate hosts in rice-wheat and rice-vegetable cropping systems that provide an alternate means for survival in the absence of agricultural crops. We show that a combination of molecular and morphological traits is a quick and reliable means to accurately identify M. graminicola. Phylogenetic analysis and genetic diversity suggests Pakistan as a putative ancestral area. M. graminicola is a significant pest of the rice-cropping system of Central Punjab, Pakistan, warranting the adoption of necessary control measures for its management.