
The genus Metarhabditis Tahseen, Hussain, Tomar, Shah and Jairajpuri, 2004 was proposed by Tahseen et al. (2004) under the family Rhabditidae Örley, 1880 with the type and only species Metarhabditis andrassyana Tahseen, Hussain, Tomar, Shah and Jairajpuri, 2004. This genus is characterized by having metastegostom with knobbed setose denticles and bursa bearing eight genital papillae. The genus was later revised by Sudhaus (2011) who transferred five species from the genera Rhabditis Dujardin, 1845 namely Rhabditis adenobia Poinar, 1971, R. blumi Sudhaus, 1974, R. costai Martins, 1985, R. freitasi Martins, 1985, and R. rainai Carta and Osbrink, 2005 and one species from Oscheius Andrássy, 1976 namely Oscheius amsactae Ali, Pervez, Andrabi, Sharma and Verma, 2011 into Metarhabditis. Recently, Abolafia and Peña-Santiago (2019a) described a new species, M. giennensis from Spain and provided a key for species identification. More recently, Tabassum et al. (2019) described other new species, M. longicaudata from Pakistan and its identity is discussed later in this paper.
One of the species recently transferred to the genus Metarhabditis is M. amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011. It is distinguished from all the other species of the genus by having large and robust posterior filiform part of the male tail (see keys to species identification provided by Abolafia and Peña-Santiago, 2019a). It was first described as Oscheius amsactae by Ali et al. (2011), who recovered some nematode specimens from a larva of the red-hairy caterpillar, Amsacta moori Butler (Lepidoptera: Arctiidae), collected in Kanpur, Uttar Pradesh, India. Since then, M. amsactae nematodes have been isolated from soil samples in different regions of India and Pakistan, many of them were previously identified as other species, however, such as Oscheius ciceri Shaheen, Ali and Asif, 2011, Oscheius hussainii Shaheen, Ali and Asif, 2011, Oscheius gingeri Pervez, Eapen, Devasahayan, and Jacob, 2012, and Oscheius amsactae Ali, Pervez, Andrabi, Sharma and Verma, 2011 and Metarhabditis longicaudata Tabassum, Salma and Nasir, 2019. Most of these studies, however, have characterized the species morphologically and morphometrically. Regarding molecular analysis, several Internal Transcribed Spacer (ITS) rDNA sequences, obtained from M. amsactae isolated in India, Philippines, and Pakistan have been deposited in the GenBank, but none of the nematode specimens used to obtain the sequences were morphologically characterized and vice versa. Hence, scanning electron microscopy images and reference molecular data for this species are still required. In this study, we therefore conducted the scanning electron microscopy (SEM) studies, and sequenced the ITS, and small-subunit (SSU) and large-subunit (LSU) rDNA of two M. amsactae isolated from Uttar Pradesh, India.
A survey to obtain the nematodes was conducted in soils of the district Shamli (29.6189° N, 77.4329° E; 280 m altitude), Uttar Pradesh, India. This location has a semiarid and moderate-to-tropical monsoon (humid subtropical) predominant climate. The type of soil is sandy loam and loamy and the pH of soil samples ranged from 6.5 to 8.4. A total of eighty-nine soil samples were taken from meadows, pastures, agricultural fields, open fields, and orchards.
Each soil sample consisted of 1 kg of soil that was a mixture of five soil subsamples collected at 15 to 20 cm depth in five locations within each field (one sample from each corner of the field, and one from the center of the field). The soil was first made fine to remove any debris (i.e., rocks, pieces of wood or bark, leaves, etc.) and then moistened with distilled water using a spray bottle to facilitate the movement of nematodes. To recover insect-associated nematodes from these soil samples, the ‘Galleria mellonella baiting’ method and the White (1927) trap method modified (Bedding and Akhurst, 1975) were used. Seven 4th instar Galleria mellonella larvae were buried in 250 ml of autoclaved plastic containers filled with the collected soil up to the brim. The plastic containers were then covered with tissue paper and muslin cloth. The containers were inverted upside-down and stored in the dark in an incubator at 27 ± 2°C for 7 days. The plastic containers were checked daily to recover dead insect larvae. Insect cadavers were rinsed with double-distilled water (ddH2O) to remove soil particles and disinfected with 0.1% sodium hypochlorite before being placed on the modified White traps to obtain emerging infective juveniles. The White traps were incubated at 27 ± 2°C in an incubator and checked daily for the emergence of third-stage juveniles from the cadavers. Emerged third-stage juveniles migrate after 5 to 7 days to water surrounding the Petri dish and nematodes were collected regularly until nematode emergence ceased after 10 to 20 days (Bhat et al., 2018, 2019).
Emerged IJs were sterilized with 0.1% sodium hypochlorite and washed with ddH2O, and finally stored in tissue culture flasks at 15°C. Third-stage juveniles were used within seven days after emergence (Aasha et al., 2019; Bhat et al., 2020a).
Nematode third-stage juveniles were surface-sterilized with 1% NaOCl. Then, greater wax moth (Galleria mellonella) larvae were injected with 100 juvenile nematodes in sterile Petri plates using a 1 ml of insulin syringe. The male, female, and juvenile (third-stage) nematode generation were recovered from White traps as described above. All nematode generations were heat-killed in Ringer’s solution and fixed in triethanolamine formalin (Courtney et al., 1955). Nematodes were infiltrated in glycerol by the Seinhorst method (Seinhorst, 1959) and processed further as described by Bhat et al. (2017). Briefly, the nematodes were kept in pure glycerol. Three females, specimens of five males, and 10 infective juvenile nematodes were mounted separately in a drop of glycerol on a clean glass slide. Paraffin wax was used to seal and to prevent the flattening of nematode specimens (Bhat et al. 2020b; Kajol et al., 2020). The morphology and morphometric analysis of the specimens was conducted using light compound microscope (Magnus MLX) and phase-contrast microscope (Nikon Eclipse 50i). Twenty specimens of adults (male and female) and 20 of juveniles were analyzed. Morphometric analyses were carried out with the aid of in-built software of the phase-contrast microscope (Nikon DS-L1). The best-preserved specimens were also photographed using a Nikon Eclipse 80i (Nikon, Tokyo, Japan) light microscope provided with differential interference contrast optics (DIC) and a Nikon Digital Sight DS-U1 camera. Micrographs were edited using Adobe® Photoshop® CS. Nematode species were identified based on morphological and morphometric characters using the key provided by Abolafia and Peña-Santiago (2019a). Demanian indices (de Man, 1881) and other ratios were calculated. The terminology used for the morphology of the stoma and spicules/gubernaculum follows the proposals by De Ley et al. (1995) and Abolafia and Peña-Santiago (2017), respectively.
For the SEM, male and female specimens preserved in glycerin were selected for observation and processed according to the Abolafia’s (2015) protocol. Thus, they were hydrated in distilled water, dehydrated in a graded mixture of ethanol-acetone series, critical point-dried with liquid carbon dioxide, and coated with gold. The mounts were examined with a Zeiss Merlin microscope (5 kV).
Genomic DNA was isolated from approximately five hundred infective juvenile nematodes by using the Qiagen Blood and Tissue Analysis Kit following the manufacturer’s protocol. A fragment of the rDNA gene containing the ITS regions (ITS1, 5.8S, ITS2) was amplified using primers 18S: 5′-TTGATTACGTCCCTGCCCTTT-3′ (forward), and 26S: 5′-TTTCACTCGCCGTTACTAAGG-3′ (reverse) (Vrain et al., 1992). The fragment containing the D2/D3 regions of the 28S rDNA gene was amplified using primers D2F: 5′-CCTTAGTAACGGCGAGTGAAA-3′ (forward) and 536: 5′-CAGCTATCCTGAGGAAAC-3′ (reverse) (Nadler et al., 2006). The 18S rDNA was amplified using primers NEM18SF: 5′-CGCGAATRGCTCATTACAACAGC-3′ (forward) and NEM18SR: 5′-GGGCGGTATCTGATCGCC-3′ (reverse) (Floyd et al., 2005). The Polymerase Chain Reaction (PCR) protocol for ITS, 18S, and D2/D3 rDNA gene amplification followed was described by Bharti et al. (2020) and Suman et al. (2020). Briefly, PCR master mix consisted of ddH2O 16.8 μl, 10x PCR buffer 2.5 μl, dNTP mix (10 mM each) 0.5 μl, 1 μl of each forward and reverse primers, dream taq green DNA polymerase 0.2 μl, and 3 μl of DNA extract. The PCR profiles used were 1 cycle of 94°C for 3 min followed by 40 cycles of 94°C for 30 sec, 52°C for 30 sec for LSU (28S) rDNA or 55°C for 30 sec for ITS rDNA or 54°C for 30 sec for SSU (18S) rDNA, 72°C for 60 sec, and a final extension at 72°C for 10 min. PCR was followed by electrophoresis (45 min, 100 volts) of 5 μl of PCR product in a 1% TAE (Tris–acetic acid–EDTA) buffered agarose gel stained with ethidium bromide (Bhat et al., 2020c; Rana et al., 2020a). The amplified products were purified and Sanger sequenced in both directions by Bioserve Technologies Ltd. (Hyderabad, India). The obtained sequences were manually curated, trimmed, and submitted to the Center for Biotechnology Information (NCBI) under accession numbers, MT873043, MT872508, and MT872503 for ITS, 28S (D2/D3) and 18S rDNA regions, respectively for the isolate CJ6, and MT873044, MT872509, and MT872504 for the same respective genes for the isolate CJ13.
The sequences were edited and compared with those already present in GenBank using the Basic Local Alignment Search Tool (BLASTN) of the National Center for Biotechnology Information (NCBI) (Altschul et al., 1990). The newly obtained ribosomal LSU (D2/D3 rDNA), SSU (18S rDNA), and ITS (ITS1, 5.8S, ITS2) rDNA sequences were manually edited using BioEdit 7.2.6 (Hall, 1999) and aligned with other relevant segments of same rDNA gene sequences available in GenBank using Clustal W alignment in the program MEGA7 (Kumar et al., 2016). Poorly aligned regions were removed from the alignments using MEGA7. The base substitution model was evaluated using jModeltest2.1.10 (Darriba et al., 2012). Phylogenetic trees were elaborated using the Bayesian inference method as implemented in the program MrBayes 3.2.7 (Ronquist et al., 2012). For analysis in jModeltest, the HKY + I + G model was selected for the ITS tree, the GTR + I + G model was selected for the 18S tree, and the GTR + G was selected for the 28S tree. The selected models were initiated with a random starting tree and ran with the Markov chain Monte Carlo (MCMC) for 1 × 106 generations. The Bayesian tree was ultimately visualized using the FigTree program 1.4.4 (Rambaut, 2018). Heterorhabditis downesi (KU573061) was used as the outgroup and to root the trees for ITS1-5.8S-ITS2 rDNA tree, Myolaimus byersi (KU180676) for LSU rDNA tree, and M. byersi (KU180665) for SSU rDNA tree.
The details of all the nematode species used in the molecular and phylogenetic study, including their updated nomenclature, accession numbers of rDNA genes, isolation source, and origin of the sequences are given in Table 4.
Nematodes of four genera: Metarhabditis, Steinernema, Heterorhabditis, and Oscheius were recovered from the eighty-nine soil samples collected in this study. Two soil samples, taken around the rhizosphere of sugarcane (Saccharum officinarum L.) and groundnut (Arachis hypogaea L.), contained Metarhabditis nematodes. Five samples were found positive for the presence of Steinernema abbasi, two for the presence of Heterorhabditis indica, and two for the presence of Oscheius sp. The rest of the samples were found negative for the presence of insect-associated nematodes. In this study, we characterized Metarhabditis nematodes. Steinernema, Heterorhabditis, and Oscheius nematodes are characterized in other studies (Bhat et al., 2020c, Rana et al., 2020b).
(Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011.
Morphometrics of Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 from India.
| Characters | Female | Male | L3 |
|---|---|---|---|
| n | 20 | 20 | 20 |
| Body length (L) | 939 ± 119 (718-1135) | 800 ± 91 (653-999) | 383 ± 55 (305-475) |
| a (L/MBW) | 16.4 ± 2.7 (10.9-21.1) | 16.7 ± 2.0 (12.7-20.7) | 18.9 ± 2.0 (15.7-22.9) |
| b (L/NL) | 5.4 ± 0.6 (4.5-6.6) | 4.9 ± 0.5 (3.8-5.9) | 3.4 ± 0.5 (2.7-4.3) |
| c (L/T) | 10.9 ± 1.7 (8.7-14.0) | 13.7 ± 2.7 (9.8-20.9) | 7.5 ± 1.2 (6.0-10.2) |
| c′ (T/ABW) | 4.1 ± 0.6 (2.9-5.2) | 2.9 ± 0.5 (1.5-3.8) | 4.8 ± 1.0 (3.0-6.0) |
| V (VA/L × 100) | 51.2 ± 2.1 (46-56) | – | – |
| Lip region width | 7.9 ± 1.4 (5-11) | 7.1 ± 1.1 (6-10) | 3.3 ± 0.5 (2-5) |
| Stoma length | 18.4 ± 2.3 (14-22) | 17.1 ± 2.3 (13-21) | 12.8 ± 1.1 (11-15) |
| Stomatal tube width | 5.0 ± 0.4 (2.5-3.5) | 3.5 ± 0.7 (2.5-5.5) | ? |
| Pharyngeal corpus length | 74 ± 5.6 (68-98) | 68 ± 4.5 (58-76) | 35 ± 2.9 (30-42) |
| Metacorpus length | 31 ± 3.9 (24-36) | 30 ± 2.6 (24-33) | 22 ± 2.4 (19-27) |
| Isthmus length | 40 ± 5.5 (40-48) | 39 ± 2.5 (35-41) | 27 ± 2.6 (23-31) |
| Bulb length | 29 ± 2.3 (26-34) | 29 ± 2.6 (24-35) | 16.8 ± 2.8 (12-24) |
| Pharynx length | 175 ± 13.3 (156-195) | 166 ± 7.3 (141-175) | 101 ± 6.5 (88-112) |
| Nerve ring – anterior end | 123 ± 15.0 (98-153) | 112 ± 9.1 (92-125) | 69 ± 10.9 (53-92) |
| Excretory pore–anterior end (EP) | 137 ± 18.2 (110-166) | 130 ± 9.1 (113-144) | 78 ± 11.7 (56-103) |
| Deirid–anterior end | 133 ± 16.0 (110-167) | 125 ± 11.0 (107-147) | ? |
| Neck length (stoma + pharynx, NL) | 173 ± 15.0 (148-195) | 163 ± 8.3 (144-176) | 114 ± 6.1 (101-124) |
| Body width at neck base | 43 ± 5.2 (32-50) | 38 ± 4.2 (32-50) | 18.7 ± 2.9 (14-25) |
| Mid-body width (MBW) | 58 ± 10.4 (43-81) | 48 ± 7.2 (40-66) | 20.5 ± 3.3 (16-29) |
| Anterior genital branch or Testis | 260 ± 38.2 (192-372) | 198 ± 15 (188-222) | – |
| Posterior genital branch | 278 ± 27.5 (229-321) | – | – |
| Vagina length | 24.5 ± 4.1 (17-30) | – | – |
| Vulva–anterior end (VA) | 480 ± 64 (380-579) | – | – |
| Rectum length | 31 ± 6.4 (22-42) | – | 15.0 ± 4.0 (9-23) |
| Anal body width (ABW) | 22 ± 2.7 (16-28) | 20.9 ± 2.5 (17-27) | 11.2 ± 2.2 (9-17) |
| Tail length (T) | 87 ± 10.5 (68-101) | 63 ± 8.2 (49-62) | 52 ± 5.9 (48-58) |
| Spicules length | – | 41 ± 7.5 (34-49) | – |
| Gubernaculum length | – | 19.6 ± 3.5 (16-28) | – |
Notes: = Character absent. ? = Measurement unknown. Measurements in μm (except n, ratio, and percentage) and in the form: mean±standard deviation (range).
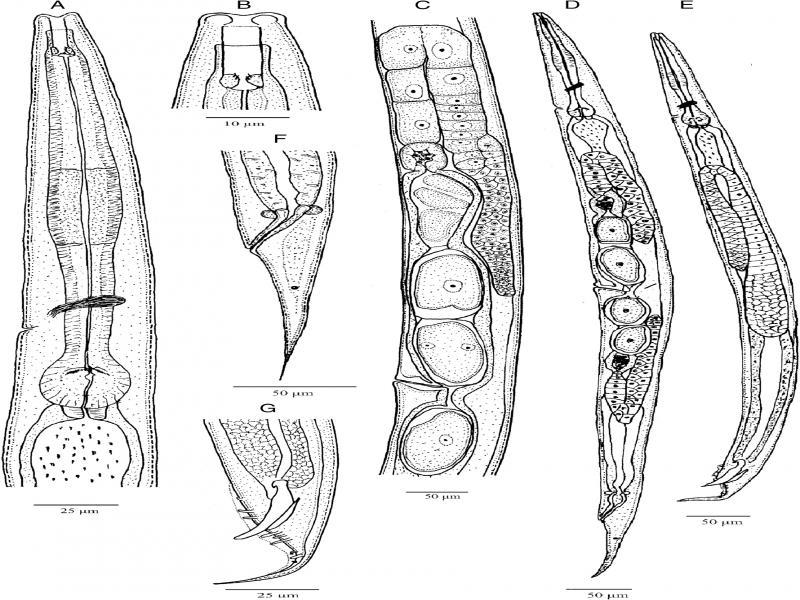
Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 (line drawing). A: Anterior region; B: Cephalic region; C: Anterior branch of the female reproductive system; D: Entire female; E: Entire male; F: Female posterior region; G: Male posterior region.
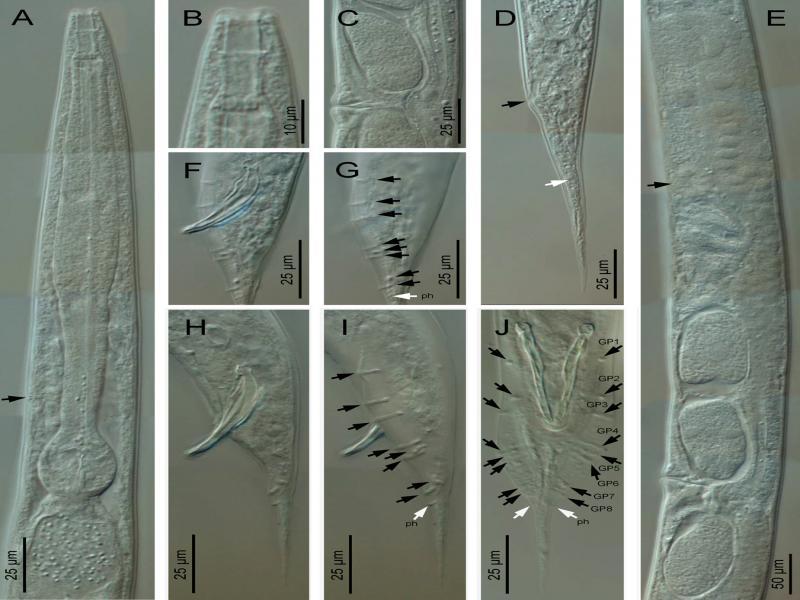
Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 (light microscopy). A: Anterior region (arrow pointing to the excretory pore); B: Cephalic region; C: Vagina region; D: Female posterior region (black arrow pointing to the anus, white arrow pointing to the phasmid); E: Anterior branch of the female reproductive system (black arrow pointing to the spermatheca); F-J: Male posterior region in lateral (F-I) and ventral (J) views, at spicules (F, H) and bursa (G, I) level (black arrows pointing to the genital papillae, white arrows pointing to the phasmids).
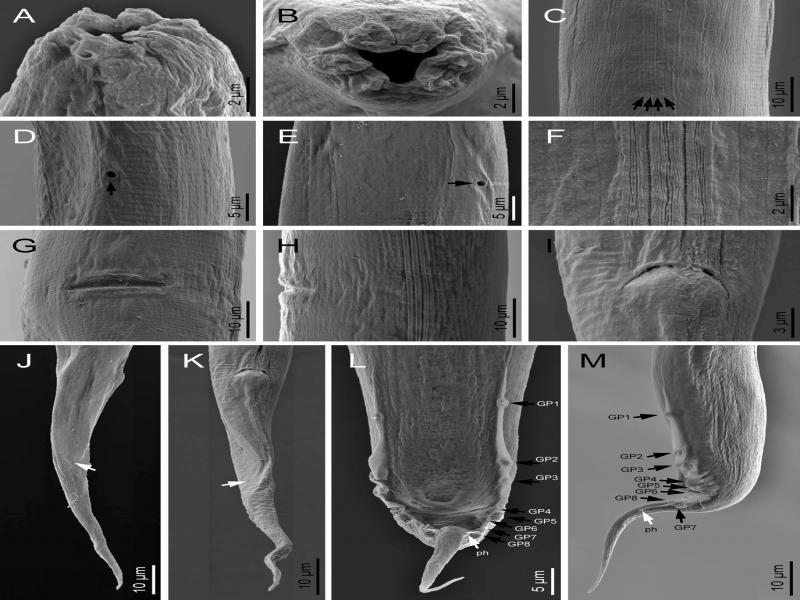
Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 (scanning electron microscopy). A, B: Cephalic and lip region in sublateral and frontal views, respectively; C, F: Lateral field (arrows pointing to the longitudinal incisures); D, E: Excretory pore (arrow) at ventral and lateral views, respectively; G, H: Vulva in ventral and lateral views, respectively; I: Female anus; J, K: Female posterior region in right lateral and ventral views, respectively (white arrows pointing to the phasmid); L, M; Male posterior region in ventral and lateral views (black arrows pointing to the genital papillae, white arrows pointing to the phasmids).
Adult: Body 0.72 to 1.14 mm long in females and 0.65 to 1.00 mm long in males, mostly straight rarely arcuate upon gentle heat killing with tapering to the anterior and posterior ends, more tapering toward the posterior end. The cuticle striated with scarcely prominent annuli 1.0 to 1.5 µm wide varying with body regions. Lateral fields were indistinct under light microscopy; however, four longitudinal lines are visible under scanning electron microscopy. The Lip region was almost continuous from contiguous body. Lips rounded and swollen, organized in doublets forming three pairs (one dorsal and two subventral) around the triradial oral orifice. Amphids small, oval, positioned at the base of lateral lips. Stoma rhabditoid type, 1.5 to 3.4 times the lip region width in length, with stomatal tube (gymno-promesostegostom) well developed. Cheilostom short with poorly refringent rhabdia; gymnostom tubular with cuticularized rhabdia, shorter than promesostegostom, this later surrounded by a thin pharyngeal collar; metastegostom isomorphic and isotopic having glottoid apparatus with three valves bearing two setose denticles per valve; telostegostom with minute rounded rhabdia. Pharynx rhabditoid, differentiated into cylinder-shaped pharyngeal corpus, 0.9 to 2.0 times the isthmus length, metacorpus slightly swollen, fusiform, isthmus relatively thick, weakly narrowing until its junction with the basal bulb, this more or less rounded, occasionally pyriform, with a weak to moderately developed valvular apparatus and faintly double-chambered haustrulum. Nerve ring surrounding the pharynx at the level of isthmus, 83 to 89% of neck length. Secretory–excretory pore at 79 to 86% of neck length, variable in position ranging from middle of isthmus to closely anterior to basal bulb. Deirids and hemizonid poorly visible, posterior to excretory pore, at 84 to 96% of neck length, at level of isthmus. Cardia small, conoid, surrounded by intestinal tissue. Intestinal lumen wider and dilated posterior to the basal bulb.
Female: Reproductive system didelphic-amphidelphic, the anterior and the posterior branches in sinistral and dextral position to intestine, respectively. Ovaries moderately developed, dorsally reflexed but with distal end not reaching to vulval level, anterior ovary slightly larger. Usually one or two small rounded pseudocoelomocytes observed in close proximity to the proximal end of ovaries. Oviducts proximally enlarged, connected to ovoid spermatheca frequently filled with sperm. Uteri well developed, differentiated into long glandular and muscular regions, filled with sperm and one to ten intrauterine eggs, 40−49 × 22−23 µm, in different stages of embryonation. Vagina thick-walled, often cuticularized, at right angle to the longitudinal body axis, with length equal to about one-third of the vulval body diameter. Vulva a wide transverse slit, with protruding lips, unremarkable or weak epiptygma but distinct cuticular flap. Rectum short, 1.0 to 1.8 times anal body diameter, allied with rectal glands at its junction with prerectum. Prerectum distinguishable from intestine in lacking prominent cell nuclei. Tail elongate conoid, gradually tapering to a fine terminus. Phasmids short tubular, located posterior to anus, about 38 to 42% of tail length.
Male: Similar to female in general morphology except for smaller size, posterior body curvature prominent and cuticular striations relatively fine. Reproductive system monorchic, with single testis reflexed ventrad anteriorly on the right side of the intestine. Vas deferens a broad tube, packed with sperm without delineation of seminal vesicles. Ejaculatory glands not observed. Spicules paired and symmetrical, ventrally arcuate, free with slightly bent ventrad manubrium, ventrally hooked, calamus short conoid and slightly ventrally curved lamina with ventrally bent finely rounded tip in lateral view. Gubernaculum well-developed, slightly ventrad curved with long manubrium and narrow corpus, 50 to 60% of spicule length. Three small gland-like cells are distinguishable around the anterior end of the cloaca. Tail conoid with posterior two-thirds abruptly tapering and reduced. Bursa anteriorly open, narrow, leptoderan, not enclosing large tail spike, having smooth margins and eight pairs (1 + 1 + 1/3 + 2 + ph) of genital papillae, with GP1 and GP2 spaced, precloacal, GP3 slightly posterior to cloaca in most specimens, pairs GP4 to GP6 located at conoid part of tail and GP7 to GP8 located at posterior part of the bursa, dorsally directed. Phasmid small, tubular, located posterior to the GP8, at 45 to 50% of tail length.
Juveniles: Third-stage juveniles ensheathed in a cuticle of second stage juveniles. Sheath free anteriorly in third-stage juveniles, firmly bound to the posterior region of the body. Body lean, from anus to tail terminus. Cuticle with transverse striae. Lip region smooth; stoma opening closed. Stoma tubular. Pharynx with pharyngeal corpus and isthmus both long and narrow, and basal bulb spheroid, valvate. Nerve ring and excretory pore located at isthmus level. Tail conoid with pointed terminus.
From the two populations of Metarhabditis amsactae molecularly analyzed in the present study from India, two sequences of 18S rDNA (865 and 869 bp), two of D2/D3 fragment of 28S rDNA (887 and 907 bp) and two of ITS1-5.8S-ITS2 rDNA (885 and 883 bp) have been obtained. Sequences of 18S and 28S rDNA are obtained for the first time for this species. A common aligned fragments resulted in 865 bp for the 18S rDNA, 879 bp for the 28S rDNA and 883 bp for the ITS rDNA, any of them show changes (substitutions, deletions or insertions) in their respective sequences.
Comparing with other sequences (unpublished) of the species available from GenBank, the 18S rDNA fragment, from a common aligned fragments with 794 bp, the present populations from India shows one (0.1%) change from the other sequence available from India (NM453373), 1 (0.1%) and 16 (2.0%) changes from the sequences submitted from Philippines (MT012150 and MT043860), respectively. For the 28S rDNA fragment, there are no other available sequences to compare. The ITS rDNA sequences, from a common aligned fragments with 645 bp, show 3 (0.5%) or 6 (0.9%) changes from other sequences submitted from India (KP834432/KP834433/KY083045 and MH392568), respectively; 1 (0.2%) or 2 (0.3%) changes with respect to two sequences from Philippines (MT422254 and MT576957), while other two sequences (MT452472 and MT576957) deposited from Philippines show too much changes (51 and 64, respectively); the sequence submitted from Pakistan (MK973071) show 24 (3.7%) changes, the most of them consistent in two long contiguous deletions in the Pakistani sequence (10 and 12 gaps, respectively, after aligning sequences), which must be considered as M. amsactae.
Twenty females and twenty males of each isolate were deposited at the museum of the Department of Zoology, Chaudhary Charan Singh University Meerut, India. Ten females and ten males were deposited at the nematode collection of the Department of Animal Biology, Plant Biology and Ecology of the University of Jaén, Spain.
Metarhabditis amsactae, including its synonyms, are characterized by having a body length of 0.72 to 2.07 mm in females and 0.65 to 1.50 mm in males, cuticle with very fine transverse striations; lips rounded and swollen grouped in pairs, stoma with metastegostoma bearing setose denticles, esophagus with metacorpus slightly swollen and fusiform, nerve ring and excretory pore located at isthmus level, female reproductive system didelphic-amphidelphic with vulva equatorial (V = 42-60), female tail conical-elongate with acute tip (65-148 µm long, c = 8.7-18.0, c′ = 2.5-8.0), female phasmids located about the middle length of the tail, male tail conical (32-76 µm long, c = 9.8-37.0, c′ = 1.0-3.8) with large and robust posterior filiform part, spicules free (24-60 µm long) with rounded manubrium slightly bent ventrad and hooked ventrally, gubernaculum 9-34 µm long, bursa open leptoderan with eight genital papillae (1 + 1/1/3 + 2 + ph) and phasmids posterior to the GP8.
The material examined in this study agrees well with the type material described by Ali et al. (2011) and the redescription of Asif et al. (2013) as M. amsactae. Morphologically, the present material does not show important morphological differences with previous described populations. With respect to other populations described from different geographical regions of India (Shaheen et al., 2011; Pervez et al., 2012 as Oscheius ciceri and O. hussaini; Asif et al., 2013 as Oscheius gingeri), the material examined now shows close similitude to each other with only variations in body length, pharyngeal corpus, and isthmus length in adult generations (see Tables 2 and 3). The variation in morphometry in the present Indian population compared with the other populations can be attributed to differences in their geographical origin.
Compendium of females of Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 populations and its synonyms.
| Species | M. amsactae | M. amsactae | M. amsactae as O. ciceri | M. amsactae as O. hussainii | M. amsactae as O. gingeri | M. amsactae | M. amsactae as M. longicaudata | M. amsactae as M. rainai | Metarhabditis sp. as M. amsactae |
|---|---|---|---|---|---|---|---|---|---|
| Reference | Present study | Ali et al. (2011) | Shaheen et al. (2011) | Shaheen et al. (2011) | Pervez et al. (2012) | Asif et al. (2013) | Tabassum et al. (2019) | Tabassum et al. (2019) | Tabassum et al. (2019) |
| Country | India | India | India | India | India | India | Pakistan | Pakistan | Pakistan |
| Habitat | Rhizosphere of sugarcane and groundnut | Rhizosphere of mungbean | Rhizosphere of chickpea | Rhizosphere of pigeonpea | Rhizosphere of ginger | Decaying matter | Rhizosphere of mango tree | Decomposed guava fruit | Rhizosphere of chicko |
| L | 718-1,135 | 658-786 | 964-1,018 | 902-989 | 1,418-1,813 | 786-902 | 1,366-1,684 | 1,769-2,078 | 1,546-1,694 |
| a | 10.9-21.1 | 19.7-22.9 | 20.1-22.5 | 24.4-25.5 | 18.5-21.2 | 19.2-23.5 | 14.0-18.0 | 11.7-20.0 | 15.0-18.0 |
| b | 4.5-6.6 | 4.1-4.8 | 5.7-5.9 | 3.8-4.3 | 5.1-5.3 | 4.1-5.0 | 5.5-7.8 | 7.0-8.0 | 6.0-8.0 |
| c | 8.7-14.0 | 8.9-12.1 | 10.3-12.9 | 10.2-12.7 | 12.1-13.2 | 9.6-11.2 | 12.0-16.0 | 12.0-18.0 | 13.0-19.0 |
| c′ | 2.9-5.2 | 4.1-4.6 | 3.6-4.3 | 3.3-4.8 | 4.6 | 3.8-4.5 | 2.5.-4.4 | 4.0-8.0 | 3.0-4.0 |
| V | 46-56 | 50-58 | 43a | 42a | 51-60 | 51-55 | 48-56 | 49-58 | 50-54 |
| Lip region width | 5-11 | 7-8 | 8-11 | 6-8 | 8-12 | 9-10 | 11-15 | 12a | 12a |
| Stoma length | 14-22 | 16-18 | 18-19 | 22-23 | 19-21 | 20-25 | 22-28 | 26-30 | 22-26 |
| Corpus length | 68-98 | 96-115 | 92-100 | 133 | 95-170 | 50a | 62-106 | 57a | 42a |
| Isthmus length | 40-48 | 32-44 | 30-40 | 44-56 | 28-57 | 35-45 | 30a | 27a | 33a |
| Bulb length | 26-34 | 24-35 | 26-31 | 33-45 | 44a | 25-35 | 19a | 16a | 17a |
| Nerve ring-ant. end | 98-153 | 99-112 | 111-134 | 170-179 | 178-203 | 110-143 | 154-190 | 76a | 95a |
| Excretory pore-ant. end | 110-166 | 109-130 | 134-136 | 166-172 | 187-223 | 121-160 | 148-250 | 98a | 112a |
| Pharynx length | 156-195 | 159-178 | 167-176 | 220-225 | 189-284 | 177-218 | 206-265 | 250-279 | 218-247 |
| Mid-body width | 43-81 | 32-39 | 39-46 | 32-40 | 75-89 | 26-40 | 78-108 | 92-111 | 88-105 |
| Anal body width | 16-28 | 16-17 | 20-28 | 19-23 | 25-28 | 19-22 | 25-33 | 12-30 | 30-36 |
| Tail length | 68-101 | 65-80 | 75-96 | 77-87 | 115-129 | 81-100 | 94-112 | 100-148 | 84-132 |
Notes: aMeasurement obtained from illustrations. All measurements are in µm (except ratio and percentage) and in the form of range.
Compendium of males of Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011 populations and its synonyms.
| Species | M. amsactae | M. amsactae | M. amsactae as O. cicero | M. amsactae as O. hussainii | M. amsactae as O. gingeri | M. amsactae | M. amsactae as M. longicaudata | M. amsactae as M. rainai | Metarhabditis sp. as M. amsactae |
|---|---|---|---|---|---|---|---|---|---|
| Reference | Present study | Ali et al. (2011) | Shaheen et al. (2011) | Shaheen et al. (2011) | Pervez et al. (2012) | Asif et al. (2013) | Tabassum et al. (2019) | Tabassum et al. (2019) | Tabassum et al. (2019) |
| Country | India | India | Pakistan | Pakistan | India | India | Pakistan | Pakistan | Pakistan |
| Habitat | Rhizosphere of sugarcane and groundnut | Rhizosphere of mungbean | Rhizosphere of chickpea | Rhizosphere of pigeonpea | Rhizosphere of ginger | Decaying matter | Rhizosphere of mango tree | Descomposed guava fruit | Rhizosphere of chicko |
| L | 653-999 | 594-804 | 754-973 | 855-889 | 673-821 | 683-868 | 1,154-1,325 | 1,100-1,392 | 1,234-1,498 |
| a | 12.7-20.7 | 16.6-19.3 | 19.4-20.8 | 25.0.-28.0 | 18.3-24.0 | 18.1-21.7 | 14.4-19.4 | 15.0-24.0 | 14.0-20.0 |
| b | 3.8-5.9 | 4.0-5.0 | 5.0-5.6 | 3.83-3.89 | 4.32-5.3 | 4.3-4.5 | 5.3-6.6 | 4.0-6.0 | 6.0-8.0 |
| c | 9.8-20.9 | 10.7-17.8 | 13.6-16.7 | 13.9-13.6 | 11.5-16.7 | 11.5-13.7 | 14.9-19.0 | 23.0-37.0 | 16.0-20.0 |
| c′ | 1.5-3.8 | 2.6-2.8 | 2.1-2.7 | 3.2 | 2.8-3.1 | 2.5-3.0 | 2.1-3.7 | 1.0-2.0 | 2.0-3.0 |
| Lip region width | 6-10 | 7-8 | 8-11 | 6-8 | 7a | 9-10 | 11-14 | ? | ? |
| Stoma length | 13-21 | 15-17 | 19 | 22-23 | 17-19 | 18-20 | 20-28 | 24-28 | 20-24 |
| Corpus length | 58-76 | 81-109 | ? | 134 | 71-112 | 72a | ? | ? | ? |
| Isthmus length | 35-41 | 27-42 | ? | 44-56 | 21-38 | 27a | ? | ? | ? |
| Bulb length | 24-35 | 20-36 | ? | 33-45 | ? | 23a | ? | ? | ? |
| Nerve ring-ant. end | 92-125 | 79-108 | 116-136 | 149-179 | 90-114 | 100-127 | 143-185 | ? | ? |
| Excretory pore-ant. end | 113-144 | 87-114 | 127-138 | 155-168 | 110-142 | 119-141 | 137-179 | ? | ? |
| Pharynx length | 141-175 | 134-169 | 149-172 | 223-228 | 142-187 | 155-190 | 184-256 | 211-256 | 204-236 |
| Midbody width | 40-66 | 31-45 | 39-46 | 30-35 | 32-39 | 26-40 | 64-69 | 54-88 | 70-98 |
| Anal body width | 17-27 | 16-20 | 26-30 | 19-21 | 16-19 | 20-24 | 21-40 | 26-34 | 27-32 |
| Tail length | 49-62 | 41-55 | 55-58 | 61-65 | 43-59 | 58-67 | 62-76 | 32-56 | 66-78 |
| Spicules length (SL) | 34-49 | 31-36 | 35-44 | 41-44 | 24-27 | 33-39 | 40-46 | 32-60 | 42-60 |
| Gubernaculum length (GL) | 16-28 | 13-17 | 19-20 | 14-18 | 9-10 | 14-20 | 20-34 | 13-23 | 16-22 |
| GL/SL × 100 | 50-60 | 43-46 | 45-54 | 34-40 | 36 | 42-51 | 50-74 | 40 | 37-38 |
Notes: aMeasurement obtained from illustrations; ? = Measurement unknown. All measurements are in µm (except ratio and percentage) and in the form of range.
Nematode species, GenBank accession number, and origin of the sequences used for phylogenetic study.
| GenBank accession number | ||||||
|---|---|---|---|---|---|---|
| Speciesa | 18S rDNA | 28S rDNA | ITS rDNA | Country | Isolation source | Reference |
| Ablechroiulus cristatus | EU196013 | EU195976 | USA | Unknown | Kiontke et al. (2007) | |
| Ablechroiulus dudichi | AF083012 | USA | Unknown | Fitch (unpublished) | ||
| Auanema rhodensis | EU196004 | USA | Unknown | Kiontke et al. (2007) | ||
| Buetschlinema nidrosiensis | EU196020 | USA | Unknown | Kiontke et al. (2007) | ||
| Bursilla belari | MK359049 | India | Soil | Palanisamy et al. (unpublished) | ||
| Bursilla sp. | EF990722 | USA | Unknown | Kiontke et al. (2007) | ||
| Caenorhabditis angaria | JN636068 | USA | Rotting fruits | Kiontke et al. (2011) | ||
| Caenorhabditis elegans | Z92784 | UK | Unknown | Sulston and Waterston (1998) | ||
| Caenorhabditis elegans | X03680 | USA | Unknown | Ellis et al. (1986) | ||
| Caenorhabditis sinica | JN636142 | USA | Rotting fruits | Kiontke et al. (2011) | ||
| Cephaloboides cf. armatus | EU196005 | USA | Unknown | Kiontke et al. (2007) | ||
| Crustorhabditis transita | EU195995 | USA | Unknown | Kiontke et al. (2007) | ||
| Cruznema tripartitum | U73449 | Unknown | Baldwin et al. (1997) | |||
| Diploscapter coronatus | AY593921 | Netherlands | Soil | Helder et al. (2006) | ||
| Diploscapter sp. | EU195959 | USA | Unknown | Kiontke et al. (2007) | ||
| Distolabrellus veechi | EF990725 | USA | Unknown | Kiontke et al. (2007) | ||
| Haematozoon subulatum | EU040125 | China | Soil | Li et al. (unpublished) | ||
| Haematozoon subulatum | AF083017 | USA | Unknown | Fitch (unpublished) | ||
| Haematozoon subulatum | EF990727 | USA | Unknown | Kiontke et al. (2007) | ||
| Heterorhabditis bacteriophora | KJ636408 | Netherlands | Unknown | van Megen (unpublished) | ||
| Heterorhabditis downesi | KU573061 | Ireland | Soil | Maher et al. (2016) | ||
| Heterorhabditis megidis | KJ636320 | Netherlands | Unknown | van Megen (unpublished) | ||
| Litoditis marina | AF083021 | USA | Unknown | Fitch (unpublished) | ||
| Litoditis marina | AM399053 | Belgium | Fucus sp. | Derycke et al. (2008a) | ||
| Litoditis marina | AM937053 | Greece | Decaying seaweeds | Derycke et al. (2008b) | ||
| Litoditis mediterranea | AF083020 | USA | Unknown | Fitch (unpublished) | ||
| Litoditis mediterranea | AM399068 | New Zealand | Unknown | Derycke et al. (2008a) | ||
| Mesorhabditis anisomorpha | AF083013 | USA | Unknown | Fitch (unpublished) | ||
| Mesorhabditis anisomorpha | EF990723 | USA | Unknown | Kiontke et al. (2007) | ||
| Mesorhabditis longespiculosa | EU196014 | EU195980 | USA | Unknown | Kiontke et al. (2007) | |
| Metarhabditis amsactae | MT872503, MT872504 | MT872508, MT872509 | MT873043, MT873044 | India | Soil | Present study |
| Metarhabditis amsactae | MT422254 | Philippines | Soil | Dichusa et al. (unpublished) | ||
| Metarhabditis amsactae | MT452472, MT452471 | Philippines | Soil | Sangcopan and Sumaya (unpublished) | ||
| Metarhabditis amsactae | MH392568, KY083045 KP834433, KP834432 | India | Soil | Chavan et al. (unpublished) | ||
| Metarhabditis amsactae | MK973071 | Pakistan | Soil | Tabassum et al. (unpublished) | ||
| Metarhabditis blumi | U13935 | Germany | Unknown | Fitch et al. (1995) | ||
| Metarhabditis blumi | MT043860 | Philippines | Soil | Andalan et al. (unpublished) | ||
| Metarhabditis blumi | MF989442 | Colombia | Bos indicus | Chaves et al. (unpublished) | ||
| Metarhabditis blumi | MT012150 | Philippines | Soil compost | Guadalquiver (unpublished) | ||
| Metarhabditis blumi | MN453373 | India | Soil | Kandhasamy and Muthugounder (unpublished) | ||
| Metarhabditis blumi | EU195965 | USA | Unknown | Kiontke et al. (2007) | ||
| Metarhabditis blumi | KM233152, KM233153 | Brazil | Cow | Bossi et al. (2015) | ||
| Metarhabditis blumi | DQ121436 | South Africa | Unknown | Jumba and Gray (unpublished) | ||
| Metarhabditis rainai | MT012135, MT012133 | Philippines | Soil | Kabalu et al. (unpublished) | ||
| Metarhabditis rainai | MT012153 | Philippines | Soil | Buldiman et al. (unpublished) | ||
| Metarhabditis rainai | JQ237848 | USA | Citrus groves | Campos-Herrera et al. (2012) | ||
| Metarhabditis rainai | AF083008 | USA | Unknown | Fitch (unpublished) | ||
| Metarhabditis rainai | EU195966 | USA | Unknown | Kiontke et al. (2007) | ||
| Metarhabditis rainai | KR011843, KR011846 | Brazil | Soil | de Brida et al. (2017) | ||
| Metarhabditis sp. | MK973071 | Pakistan | Soil | Tabassum et al. (unplublished) | ||
| Myolaimus byersi | KU180665 | KU180676 | Sweden | Unknown | Holovachov et al. (2015) | |
| Oscheius carolinensis | FJ547240 | FJ547239 | USA | Vermicompost | Ye et al. (2010) | |
| Oscheius chongmingensis | EF503692 | China | Soil | Zhang et al. (2008) | ||
| Oscheius chongmingensis | EU273599 | China | Soil | Liu et al. (2012) | ||
| Oscheius chongmingensis | MT548593 | China | Spodoptera frugiperda | Li et al. (unpublished) | ||
| Oscheius colombianus | AY751546 | Colombia | Cyrtomenus bergi | Stock et al. (2005) | ||
| Oscheius dolichura | EU196010 | USA | Unknown | Kiontke et al. (2007) | ||
| Oscheius dolichuroides | AF082998 | USA | Unknown | Fitch (unpublished) | ||
| Oscheius dolichuroides | EU195970 | USA | Unknown | Kiontke et al. (2007) | ||
| Oscheius guentheri | EU196022 | USA | Unknown | Kiontke et al. (2007) | ||
| Oscheius indicus | MF441252 | India | Soil | Kumar et al. (2019) | ||
| Oscheius insectivorus | AF083019 | USA | Unknown | Fitch (unpublished) | ||
| Oscheius insectivorus | EU195968 | USA | Unknown | Kiontke et al. (2007) | ||
| Oscheius myriophilus | AY602176 | USA | Unknown | Kiontke et al. (2004) | ||
| Oscheius myriophilus | MG742117 | Thailand | Soil | Nitjarunkul et al. (unpublished) | ||
| Oscheius myriophilus | U13936 | USA | Soil | Fitch et al. (1995) | ||
| Oscheius necromenus | KT884894 | Australia | Oncocladosoma castaneum | Carta et al. (2018) | ||
| Oscheius onirici | MG551687 | Portugal | Unknown | Campos-Herrera (unpublished) | ||
| Oscheius onirici | LN613263 | Italy | Soil | Torrini et al. (2015) | ||
| Oscheius rugaoensis | JQ002566 | China | Soil | Zhang et al. (2012) | ||
| Oscheius rugaoensis | KT884891 | Japan | Chamberlinius hualienensis | Carta et al. (2018) | ||
| Oscheius saproxylicus | MK959600 | MK959601 | Spain | Decaying wood | Abolafia and Pena-Santiago (2019b) | |
| Oscheius tipulae | EU195969 | USA | Unknown | Kiontke et al. (2007) | ||
| Oscheius tipulae | KP792649 | Brazil | Soil | Campos-Herrera and Pu˚ža (unpublished) | ||
| Parasitorhabditis obtusa | EF990724 | Germany | Bark beetles | Kiontke et al. (2007) | ||
| Parasitorhabditis obtuse | EU003189 | USA | Unknown | Kiontke et al. (2007) | ||
| Pellioditis typica | AF036946 | Kenya | Feces of an antelope | Adams et al. (1998) | ||
| Pelodera pseudoteres | EU196023 | EU195997 | USA | Unknown | Kiontke et al. (2007) | |
| Pelodera teres | AF083002 | USA | Unknown | Fitch (unpublished) | ||
| Pelodera teres | EU195979 | USA | Unknown | Kiontke et al. (2007) | ||
| Phasmarhabditis hermaphrodita | DQ639980 | Scotland | Nemaslug | MacMillan et al. (2006) | ||
| Phasmarhabditis neopapillosa | DQ639982 | Scotland | Arion ater | MacMillan et al. (2006) | ||
| Phasmarhabditis papillosa | KX267675 | South Africa | Deroceras reticulatum | Pieterse et al. (2017) | ||
| Poikilolaimus floridensis | AB370214 | USA | Termites | Kanzaki et al. (2009) | ||
| Poikilolaimus oxycercus | AF083023 | USA | Unknown | Fitch (unpublished) | ||
| Poikilolaimus oxycercus | EU195984 | USA | Unknown | Kiontke et al. (2007) | ||
| Poikilolaimus piniperdae | DQ059060 | Germany | Unknown | Hong et al. (2005) | ||
| Poikilolaimus regenfussi | AF083022 | USA | Unknown | Fitch (unpublished) | ||
| Poikilolaimus regenfussi | DQ059057 | Germany | Unknown | Hong et al. (2005) | ||
| Protorhabditis sp. | AF083001 | USA | Unknown | Fitch (unpublished) | ||
| Protorhabditis sp. | AY602168 | USA | Unknown | Kiontke et al. (2007) | ||
| Rhabditella axei | U13934 | Germany | Unknown | Fitch et al. (1995) | ||
| Rhabditella axei | AY602177 | USA | Unknown | Kiontke et al. (2004) | ||
| Rhabditis brassicae | EU196006 | USA | Unknown | Kiontke et al. (2007) | ||
| Rhabditis sp. | MN082353, MN082355 | Taiwan | Soil | Yang et al. (2020) | ||
| Rhabditoides inermis | AF082996 | Unknown | Fitch (unpublished) | |||
| Rhabditoides regina | EF990726 | USA | Unknown | Kiontke et al. (2007) | ||
| Rhomborhabditis regina | KX385908 | Mexico | White grub | Jiménez-Cortés et al. (2016) | ||
| Teratorhabditis mariannae | EF990716 | EF990721 | USA | Soil compost | Kanzaki et al. (2008) | |
| Teratorhabditis palmarum | U13937 | USA | Unknown | Fitch et al. (1995) | ||
Note: aSpecies names have been updated according their current nomenclature.
Recently, Tabassum et al. (2019) described a new species, Metarhabditis longicaudata Tabassum, Salma and Nasir, 2019 from Pakistan. According to its morphology, especially males with posterior filiform part well developed, robust, and bursa posteriorly appearing parallel along it at its proximal part (unfortunately, the LM Fig. 2C, D provided by these authors seems to be strongly stretched making the stoma too much long and narrow, and not agreeing with the line drawing Fig. 1C, D provided by these same authors), and morphometric characteristics (Tables 1 and 2), the specimens described are highly similar to M. amsactae. Given these considerations, we propose that Metarhabditis longicaudata is a junior synonym of Metarhabditis amsactae. Moreover, the specimens described as M. amsactae in the same study by these Pakistani authors do not present characteristics of this species, especially because the males lack posterior filiform part of the tail, spicules lack ventral bent or hooked manubrium, and females have a long rectum. In addition, the nematode population described as M. rainai (Carta and Osbrink, 2005) Sudhaus, 2011 in the same study by Tabassum et al. (2019) are morphologically very similar to M. amsactae nematodes (Tables 1 and 2) and, hence they were misidentified. ITS-phylogenetic trees support these conclusions as the sequences submitted to GenBank by these authors, Metarhabditis sp. (MK973071), cluster together with other M. amsactae (Fig. 4).
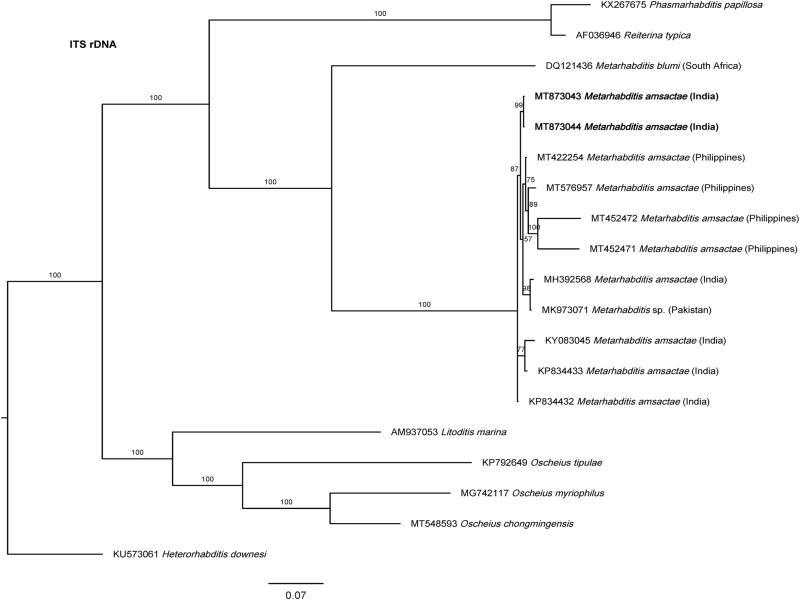
Bayesian Inference tree from previously and the newly sequenced Metarhabditis amsactae (bold) and other closely related species based on sequences of the Internal Transcribed Spacer (ITS1-5.8S-ITS2) rDNA region. Bayesian posterior probabilities (%) are given for each clade. The scale bar shows the number of substitutions per site.
According to this, the updated list of synonyms of Metarhabditis amsactae is as follows:
Metarhabditis amsactae (Ali, Pervez, Andrabi, Sharma and Verma, 2011) Sudhaus, 2011
= Oscheius amsactae Ali, Pervez, Andrabi, Sharma and Verma, 2011
= Oscheius ciceri Shaheen, Ali and Asif, 2011
= Oscheius hussainii Shaheen, Ali and Asif, 2011
= Oscheius gingeri Pervez, Eapen, Devasahayan and Jacob, 2012
= Metarhabditis longicaudata Tabassum, Salma and Nasir, 2019
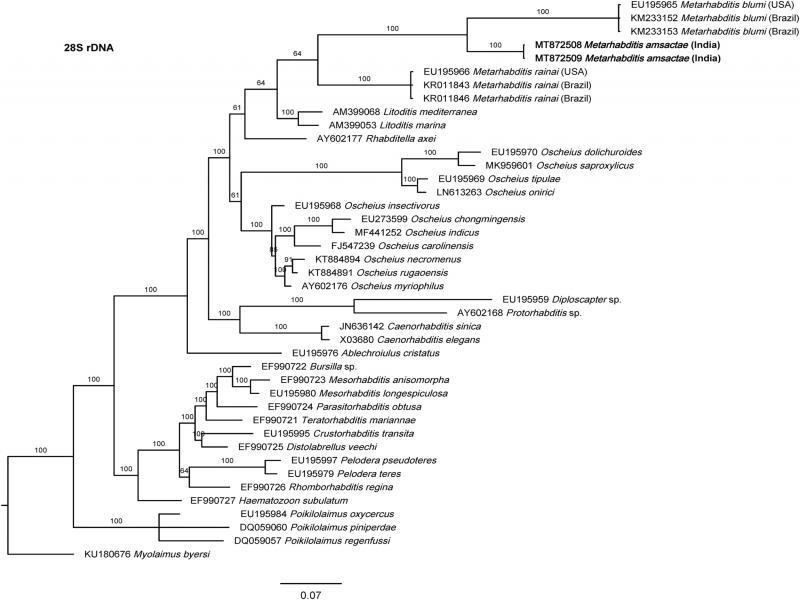
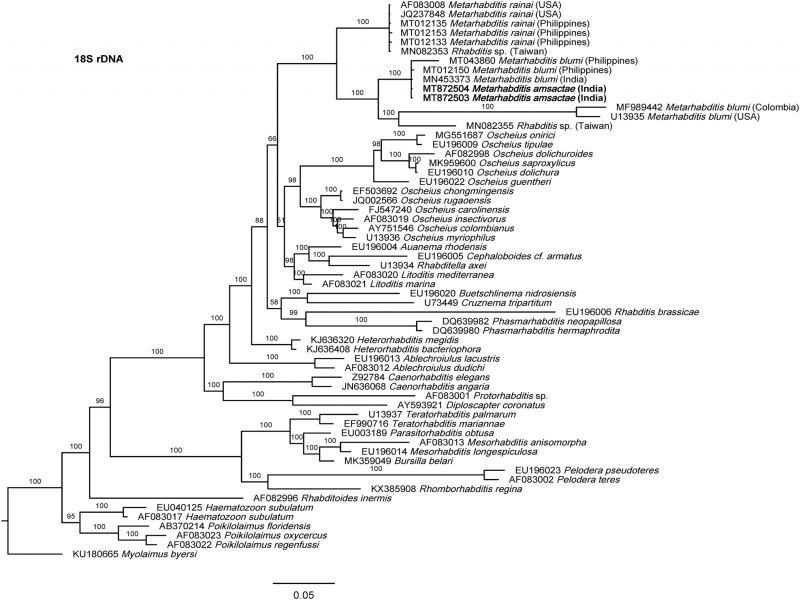
The phylogenetic relationships as inferred from the Bayesian Inference analysis between Metarhabditis amsactae and other closely related are provided based on ITS- (Fig. 4), 18S- (Fig. 5), and 28S- (Fig. 6) rDNA fragments. Based on the three phylogenetic trees, M. blumi (Sudhaus, 1974) Sudhaus, 2011 and M. rainai (Carta and Osbrink, 2005) Sudhaus, 2011 are sister species of M. amsactae.
Bayesian Inference tree from the newly sequenced Metarhabditis amsactae (bold) and other closely related species based on sequences of the small subunit (18S) of rDNA region. Bayesian posterior probabilities (%) are given for each clade. The scale bar shows the number of substitutions per site.
Bayesian Inference tree from the newly sequenced Metarhabditis amsactae (bold) and other closely related species based on sequences of the D2/D3 domain of large subunit (28S) of rDNA region. Bayesian posterior probabilities (%) are given for each clade. The scale bar shows the number of substitutions per site.
The phylogenetic tree inferred using 18S rDNA gene sequences, shows three clusters that contain sequences of nematodes that have been suggested to belong to Metarhabditis (Fig. 5). One cluster is composed of M. rainai (AF083008, JQ237848, MT012133, MT012135 and MT012153) and one nematode isolate that might have been misidentified as Rhabditis sp (MN082353) but could correspond to M. rainai. A second cluster composed of M. amsactae (MT872504, MT872503) and three nematode isolates that might have been misidentified as M. blumi (MT043860, MT012150, MN453373). A third cluster composed of M. blumi (MF989442, U13935), and Rhabditis sp. (MN082355). As M. amsactae isolates that correspond to accession numbers MT872504 and MT872503, and the M. blumi isolate that correspond to accession numbers U13935 have been morphologically and molecularly characterized, we conclude that nematodes isolates with NCBI accessions MT043860, MT012150 and MN453373 are actually M. amsactae instead of M. blumi, and the nematodes isolate with NCBI accession MN082355 identified as Rhabditis sp. should correspond to Metarhabditis sp. This conclusion is also supported by sequence identity analysis. Comparing the nucleotide composition of a common 18S rDNA gene fragment of 723 bp in length of the M. amsactae specimens examined in this study (MT872503-4) and the nucleotide composition of M. blumi (U13935), Rhabditis sp. (MN082355), M. rainai (AF083008, JQ237848, MT012133, MT012135 and MT012153), Rhabditis sp. (MN082353), and M. blumi (MT043860, MT012150 and MN453373), we found 57 genetic changes (insertions, deletions or substitutions) between M. amsactae and M. blumi, 69 genetic changes between M. amsactae and Rhabditis sp. (MN082355), 82 changes between M. amsactae and M. rainai, 82 changes between M. amsactae and Rhabditis sp. (MN082353), 95 changes between M. blumi and M. rainai, and fewer genetic changes between Rhabditis sp. (MN082353) and M. rainai, and between the M. amsactae (MT872503-4) and M. blumi (MT043860, MT012150 and MN453373).
The phylogenetic tree inferred using 28S rDNA gene sequences show also three clusters containing Metarhabditis nematodes: one with M. blumi, one with M. amsactae, and one M. rainai (Fig. 6). Comparing the nucleotide composition of a common 28S rDNA fragment of 343 bp in length of the M. amsactae specimens examined in this study (MT872508-9) and the nucleotide composition of M. blumi (EU195965, KM233152-3), and M. rainai (EU195966, KR011843-6), we found 75 genetic changes (insertions, deletions or substitutions) between M. amsactae and M. blumi, 80 genetic changes between M. amsactae and M. rainai, and 95 changes between M. blumi and M. rainai, suggesting that M. amsactae, M. rainai and M. blumi are sister species and that the nematode isolates characterized in this study belong to M. amsactae.
Finally, analyzing ITS rRNA gene sequences, we arrive to the same conclusions derived from the analysis of 28S- and 18S rDNA gene sequences. ITS-based phylogenetic tree show a clear cluster that separates M. amsactae and M. blumi (Fig. 4). Unfortunately, there are no available M. rainai sequences. Sequence comparisons show that a common ITS rDNA fragment of 802 bp in length of M. amsactae (MT873043-4) and of M. blumi (DQ121436) differ in 496 changes, which again support the status of the nematode isolates of this study as M. amsactae (Table 4).