
The impetus for the study was the initiative by the Management Unit of Protected Areas of Thessaly (formerly Management Body of Karla-Mavrovouni-Kefalovryso Velestinou-Delta Piniou) to initiate management actions related to protected sites, species, and ecotopes within its jurisdiction. This action was focused on two Natura 2000 sites within the jurisdiction of the Management Unit, with considerable diversity in native orchids (family Orchidaceae). The two Natura sites are GR1430008 (Oros Pilio) and GR1420004 (Karla-Mavrovouni-Kefalovryso Velestinou-Neochori), altogether comprising 796 km2. The two sites roughly correspond to the geographical extent of Mounts Pelion and Mavrovouni, respectively. The objective reported here was to record the distribution of native orchids in the study area with emphasis on five taxa of conservation interest, namely Ophrys cornutula (syn. O. oestrifera subsp. cornuta, O. scolopax subsp. cornuta), Spiranthes spiralis, Ophrys umbilicata subsp. umbilicata, Orchis provincialis, and O. simia.
Initially, we collected data about the terrestrial habitats in the study area. The following broad habitat groupings derive from the Standard Data Forms of the two Natura 2000 sites, with contribution in parenthesis (European Environment Agency, 2024a, 2024b):
Broad-leaved deciduous woodland (34,030 ha)
Shrubland such as maquis, phrygana, etc. (23,005 ha)
Arable land with herbaceous and woody crops (13,784 ha)
Evergreen woodland (2,500 ha)
Grassland (1,922 ha)
Mixed woodland (1,721 ha)
Coniferous woodland (539 ha)
For Mount Pelion, Fotiades (2005) reports 14 ecotopes, without reference to their respective contribution:
Precipitous, rocky cliffs with Mediterranean vegetation
Short communities of Euphorbia near precipitous, rocky cliffs
Phrygana of Sarcopoterium spinosum
Sclerophyllous woodland used for grazing (dehesas) of Quercus ilex
Degraded shrubland of kermes oak (garrigues)
Beech woodland
Chestnut woodland
Quercus frainetto woodland
Platanus orientalis woodland
Caves not exploited for tourism
Quercus ilex woodland
Azonal vegetation along rivers of permanent or interrupted flow, in the form of gallery dominated by Salix sp. and Populetea albae
Thickets of Juniperus communis on calcareous undisturbed land
Thickets of laurel
Evidently, ecotopes such as caves are unlikely to host orchids; the remaining ecotopes can be easily transferred to the broad habitat groupings derived from the Natura 2000 Standard Data Forms.
The broader study area belongs to the Meso-mediterranean bioclimatic zone. We witnessed a sharp ecological gradient in the overall direction NE to SW. This is the result of Mounts Pelion and Mavrovouni catching most of the humidity that derives from the Aegean Sea before it reaches the hinterland. This ecological gradient results in a rich variety of ecosystems unfolding within a span of only a few kilometres (for details, see Raus, 1979a: 569 ff. and Figs. 5 & 6).
The Aegean coast is quite forested, with maquis of holly oak (Quercus ilex) and strawberry tree (Arbutus unedo) at sea level, gradually being replaced by chestnut (Castanea sativa) and Hungarian oak (Quercus frainetto), culminating in beech (Fagus sylvatica), up to the mountain ridge. As we descend from the mountain ridge to the drier plains of Thessaly, tall deciduous forest is gradually being replaced by pseudomaquis of kermes oak (Q. coccifera), that may further deteriorate into phrygana or grassland (for details, see Raus, 1979b).
The geology appears to be heterogeneous. The mountain range is predominantly composed of Paleozoic crystalline rocks, i.e., metamorphic rocks, especially gneisses and mica schists, partly interspersed with marble. The crystalline rocks are mostly found in the mid-elevations and in the W of Pelion they extend down to the coastal zone. Mesozoic limestones and dolomites overlie the crystalline rocks, mainly at higher elevations. Additionally, these can also appear on a small scale in the coastal zone, possibly due to tectonic disturbances. The soils are typically brown soils and reddish-brown Mediterranean soils with highly variable pH values (Biel & Rudolph, 1992).
Since most of the orchid species of conservation interest are photophilic, we concentrated our sampling effort on the SW slope of Mounts Pelion and Mavrovouni and excluded the extensive tracts of tall, broad-leaved and mixed forest in the NE and central part of the study area. Broad-leaved and mixed forest, especially of beech and Hungarian oak, is so dense that little light reaches the forest floor; during our preliminary reconnaissance in these ecosystems, we observed the occasional orchids only in clearings and along the roadside.
We tried to find historical surveys of the family Orchidaceae within the study area. As a starting point, we consulted the Atlas of Greek Orchids (Antonopoulos & Tsiftsis, 2017; Tsiftsis & Antonopoulos, 2017). The distribution maps are microscopic thumbnails where each location is represented by a dot 10 km wide, not useful for planning a detailed survey.
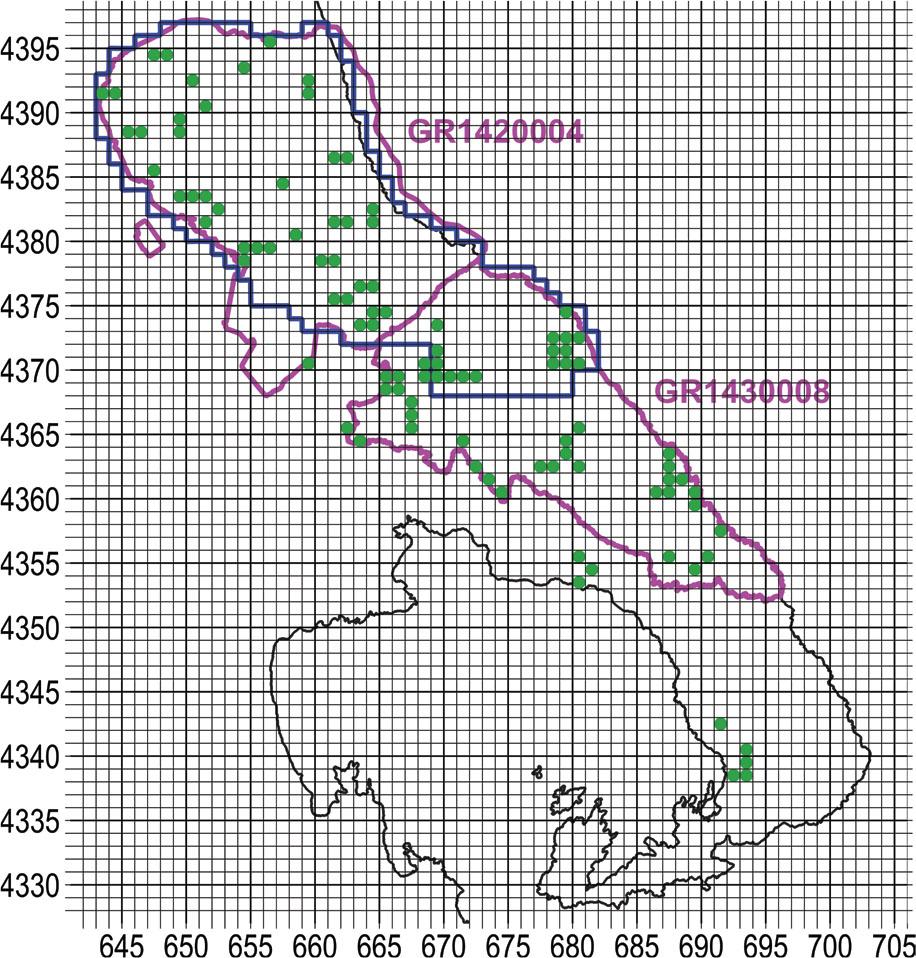
The useable reference that we could find with respect to the orchids of Pelion and Mavrovouni was by Biel & Rudolph (1992), albeit with dated information of 30 years ago. A span of 30 years is long enough for cultivated fields to be reforested, or, conversely, for forest to be cleared; moreover, urban and tourist development encroach on natural ecosystems. Still, the work of Biel & Rudolph (1992) offered a very useful advantage, in that locations were recorded with precision of 0.1 km. Their survey covered the entire Natura site GR1430008 (Oros Pilio), while from Natura site GR1420004 (Karla-Mavrovouni-Kefalovryso Velestinou-Neochori) only the SE portion was incorporated (Figure 1).
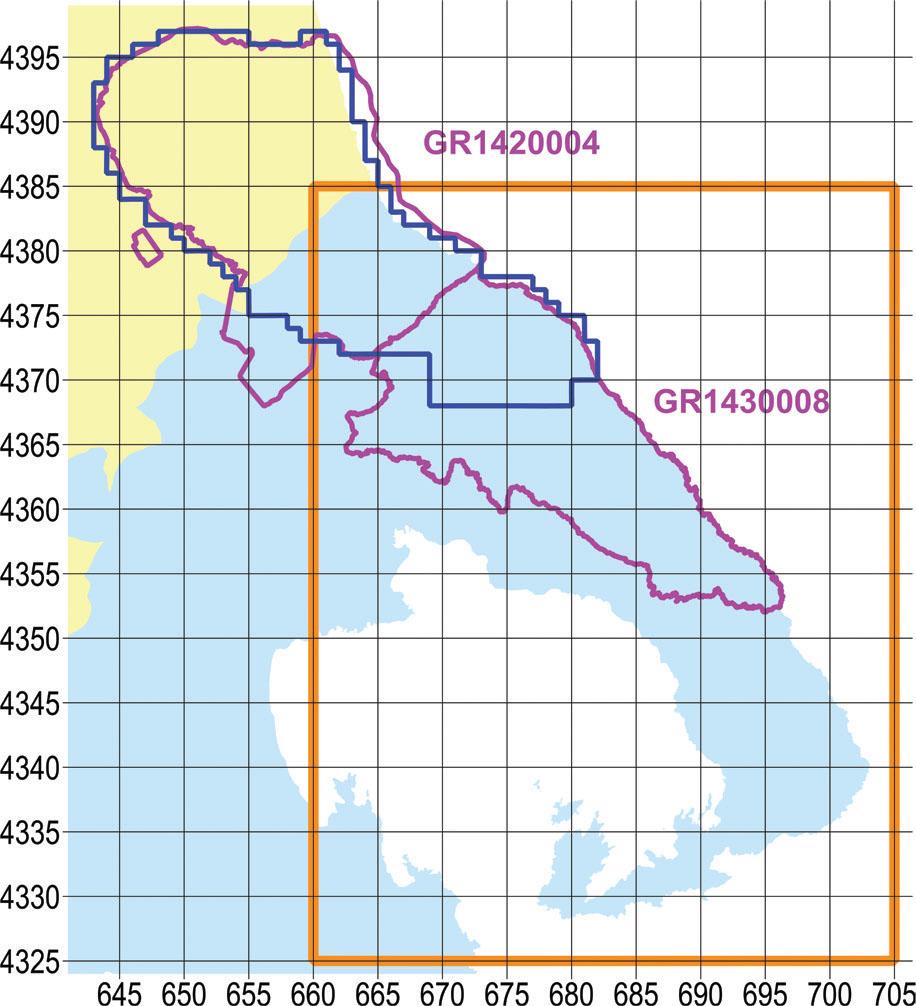
Map of the broader study area. With yellow ochre fill the prefecture of Larissa, with light blue fill the prefecture of Magnesia, with magenta outline the two Natura 2000 sites, with dark blue the outline of the narrow study area, and with orange outline the study area of Biel & Rudolph (1992). UTM grid spacing 5 km.
Slika 1: Zemljevid širšega preučevanega območja. Z rumeno je označena prefektura Larissa, s svetlomodro pa prefektura Magnesia, s škrlatno barvo obe območji Natura 2000, s temno modro barvo je označeno ožje preučevano območje in z oranžno preučevano območje v članku Biel & Rudolph (1992). Velikost kvadrantov UTM mreže je 5 km.
We tallied the findings of Biel & Rudolph (1992). In the two Natura 2000 sites, the authors altogether report 36 orchid taxa (species and subspecies) and one nothotaxon (Table 1). In general, we follow the orchid nomenclature of POWO (2024). In particular, this includes the name change of Ophrys sphegodes subsp. mammosa to O. sphegodes subsp. taurica. The first use of the epithet taurica at subspecific rank in 1890 antedates the first use of the epithet mammosa at subspecific rank in 1926 (Art. 11.4 Shenzhen Code); for details, see Niketić et al. 2018, accepted by POWO 2024 based on Güner et al. (2022). As an exception from POWO, we use Ophrys sicula instead of O. lutea subsp. galilaea based on Hennecke (2016).
List of taxa (species and subspecies) recorded during the sampling period 2021–22 within the broader study area. In bold are those recorded within the narrow sampling area. Site numbers refer to Appendix I. Checkmark indicates mention of the taxon in the Flora Hellenica Database within the narrow sampling area.
Tabela 1: Seznam taksonov (vrst in podvrst) vzorčenih v obdobju od 2021 do 2022 v preučevanem območju. Številke nahajališč se nanašajo na Prilogo I. Kljukica označuje oznako taksona v bazi Flora Hellenica znotraj ožjega preučevanega območja.
| S/N | TAXON | SITES | FLORA HELLENICA |
|---|---|---|---|
| 1 | Anacamptis coriophora | 99 | ✓ |
| 2 | Anacamptis morio subsp. caucasica | 21, 24, 35, 43, 46, 48, 49, 52, 53, 55, 59, 80, 99, 103, 104, 105, 106 | ✓ (sub A. morio) |
| 3 | Anacamptis papilionacea subsp. papilionacea | 16, 21 | |
| 4 | Anacamptis pyramidalis | 27, 79 | |
| 5 | Cephalanthera longifolia | 29, 32, 33, 37, 46, 68, 71, 74, 76, 77, 82, 84, 85, 88, 89, 90, 96, 100, 104 | ✓ |
| 6 | Cephalanthera rubra | 32, 73, 77 | ✓ |
| 7 | Dactylorhiza romana | 10, 21, 45, 53, 54, 83, 87, 89, 90, 106 | ✓ |
| 8 | Dactylorhiza saccifera | 67, 69, 70, 72, 73, 75, 76, 82, 89, 90 | ✓ |
| 9 | Epipactis helleborine | 64, 68 | ✓ |
| 10 | Epipactis microphylla | 23, 32, 36, 73, 82, 89, 91 | ✓ |
| 11 | Himantoglossum calcaratum subsp. rumelicum | 62, 66 | ✓ (sub. H. jankae) |
| 12 | Himantoglossum robertianum | 3, 4, 28, 46 | |
| 13 | Limodorum abortivum | 3, 5, 8, 11, 14, 43, 66, 77, 84, 90, 94, 100 | ✓ |
| 14 | Neotinea maculata | 27, 32, 60, 76, 77, 89, 90, 91, 92, 94 | ✓ |
| 15 | Neotinea tridentata | 24, 40 | ✓ |
| 16 | Neottia nidus-avis | 73, 77 | ✓ |
| 17 | Ophrys × delphinensis | 12 | |
| 18 | Ophrys apifera | 99 | |
| 19 | Ophrys ferrum-equinum subsp. ferrum-equinum | 33 | |
| 20 | Ophrys reinholdii subsp. reinholdii | 13, 43, 44, 53 | ✓ (sub O. reinholdii) |
| 21 | Ophrys scolopax subsp. cornuta | 13, 18, 20, 25, 41, 43, 58, 65, 66, 77, 78, 81, 99, 103 | ✓ |
| 22 | Ophrys sicula (syn. O. lutea subsp. galilaea) | 12, 18, 26, 29, 33, 37 | |
| 23 | Ophrys speculum subsp. speculum | 6, 7 | |
| 24 | Ophrys sphegodes subsp. sphegodes (syn. O. hebes) | 43 | |
| 25 | Ophrys sphegodes subsp. spruneri | 26 | ✓ |
| 26 | Ophrys sphegodes subsp. taurica | 1, 2, 3, 5, 6, 8, 12, 18, 22, 25, 26, 43, 46, 60, 66 | ✓ (sub O. sphegodes subsp. mammosa) |
| 27 | Ophrys tenthredinifera | 1, 2, 3, 6 | |
| 28 | Ophrys tenthredinifera × O. sphegodes subsp. taurica | 3 | |
| 29 | Ophrys umbilicata subsp. umbilicata | 1, 2, 6, 25, 26 | |
| 30 | Orchis anthropophora | 24, 28, 29, 33, 37, 39, 41, 42, 45, 47, 60, 63 | ✓ |
| 31 | Orchis italica | 3, 12, 17, 18, 21, 33, 37, 38, 40, 42, 45, 46, 47, 50, 51, 52, 53, 56, 57, 66, 79, 80, 81, 86, 101 | |
| 32 | Orchis pauciflora | 24, 28, 42, 63 | ✓ |
| 33 | Orchis provincialis | 9, 35, 60, 77, 83, 89, 90, 91, 92, 93, 95 | ✓ |
| 34 | Orchis quadripunctata | 18, 28, 29, 30, 31, 34, 41, 42, 56, 57, 61, 63 | ✓ |
| 35 | Orchis simia | 24, 29, 34, 39, 42 | |
| 36 | Platanthera bifolia | 76 | |
| 37 | Platanthera chlorantha | 32, 77 | ✓ (sub P. chlorantha subsp. chlorantha) |
| 38 | Serapias bergonii | 103, 104 | ✓ (sub S. vomeracea subsp. laxiflora) |
| 39 | Serapias lingua | 104, 105, 106, 107 | ✓ |
| 40 | Serapias vomeracea | 2, 12, 48, 102 | ✓ (sub S. vomeracea subsp. vomeracea) |
| 41 | Spiranthes spiralis | 3, 5, 9, 10, 15, 18, 19, 31, 35, 66, 97, 98, 99 | ✓ |
In the spring season of 2021, we implemented a preliminary reconnaissance. The objectives were (i) to explore as many different ecosystems as possible in order to get a general picture of orchid diversity in the study area and (ii) make well founded decisions on the following main phase of the survey.
We adopted a gradient-directed transect sampling or “gradsect” (Austin & Heyligers, 1989). In a mountainous region, an obvious gradient is that of elevation, which in turn determines the gradients of climatic factors and, by extension, vegetation. In our study area there is an elevation gradient with overall direction NE to SW (Figure 2). We selected paved roads that more or less followed the elevation gradient, meaning that they traversed the two Natura sites in the general direction NE to SW.
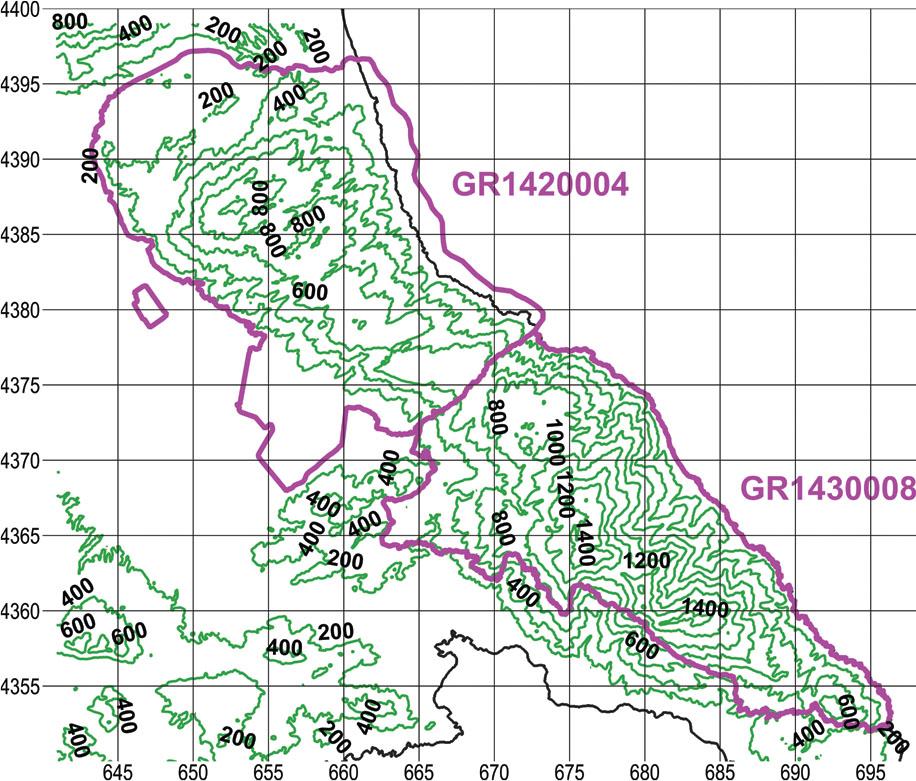
Elevation map of the two Natura 2000 sites, outlined in magenta. Contour interval 200 m, UTM grid spacing 5 km.
Slika 2: Karta nadmorskih višin obeh območij Natura 2000 označenih s škrlatno barvo. Izohipse so v intervalu 200 m, velikost kvadrantov UTM mreže je 5 km.
The southern boundary of our more narrowly delimited intensive investigation area had been agreed with the supervisory authority to coincide with the imaginary line that follows the brook between the villages of Zagora and Pouri, passes through Leschiani bridge and ends up at the hamlet of Ano Kerasia. The area thus described includes the whole of Mount Mavrovouni and the northern slope of Mount Pelion and covers 550 km2 in total (Figure 1).
To map the study area we opted for the Universal Transverse Mercator (UTM) projection system (“Universal Transverse Mercator coordinate system,” 2024) with WGS84 as geodetic reference system. UTM coordinates facilitated comparison of results with the earlier survey of Biel & Rudolph (1992).
As base maps we used digitised versions of georeferenced hiking maps (Anavasi, 2008, 2022), the digitised administrative boundaries of Greece (Harvard Library, 2024), and the digitised outlines of Natura 2000 sites (Ministry of Environment and Climate Change, 2024).
According to Delforge (2006), the five taxa of conservation interest prefer open habitats of thin, short vegetation, including open woodland, except for Orchis provincialis that exhibits a preference for damper environments, a fact that was confirmed by our preliminary scouting. In Figure 3 we mark the preliminary findings of Orchis provincialis on a precipitation map (Hellenic National Meteorological Service, 2024). It seems that O. provincialis favours the wetter foothills of E Mount Pelion at elevations of 200–600 m asl.
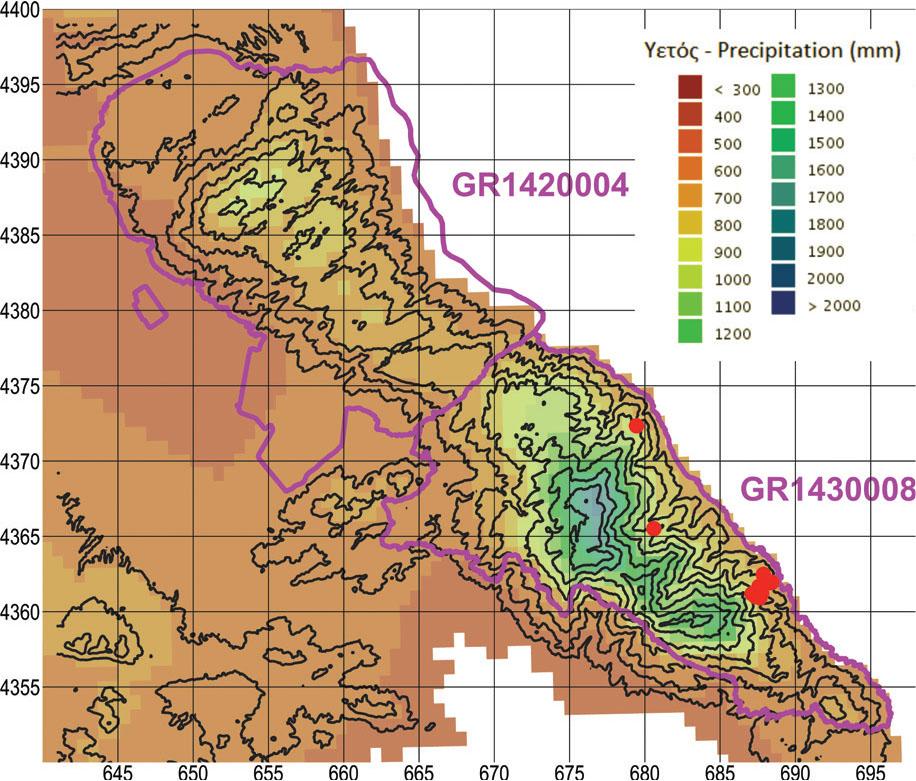
Precipitation map of the two Natura 2000 sites, outlined in magenta. Red dots represent findings of Orchis provincialis during the preliminary scouting in the spring of 2021. UTM grid spacing 5 km.
Slika 3: Karta padavin dveh območij Natura 2000 označenih s škrlatno barvo. Rdeče točke predstavljajo nahajališča vrste Orchis provincialis med preliminarnim pregledom spomladi leta 2021. Velikost kvadrantov UTM mreže je 5 km.
We estimated that project resources (time and money) were sufficient to sample 37 square cells 1 km wide or nearly 7% of the narrow study area of 550 km2. Thus, the narrow study area was divided into 550 square cells 1 km2. For the main sampling phase, approximately one-third of the 37 sampling units were allocated according to the following criteria:
- (i)
Findings of the supervisory authority: We assigned one sampling unit to each of three biotopes where the staff of the supervisory authority had recorded specimens of Ophrys cornutula (syn. O. scolopax subsp. cornuta), O. umbilicata subsp. umbilicata, and Orchis simia.
- (ii)
Literature: We assigned one sampling unit to each of three biotopes where Biel & Rudolph (1992) had recorded specimens of Ophrys cornutula (syn. O. scolopax subsp. cornuta). The mentioned authors did not record Spiranthes spiralis, Ophrys umbilicata subsp. umbilicata, or Orchis simia within the narrow study area.
- (iii)
Most promising habitats: Assuming that the five species of interest are of equal importance, six of the 31 remaining sampling units were allocated to Orchis provincialis in the wetter zone of E Mount Pelion at elevations of 200–600 m asl.
The remaining 25 sampling units (approximately two-thirds) were randomly assigned to the most promising habitats of the other four species of conservation interest. These are habitats of thin, short vegetation. The pros and cons of random sampling designs for terrestrial surveys are analysed by Webster & Oliver (1990).
To identify habitats of thin, short vegetation, we used the georeferenced, ortho-corrected maps of Corine Land Cover (Hellenic Mapping and Cadastral Organization, 2015). The CLC classes that correspond to shrub and/or herbaceous vegetation associations are:
3.2.1. Natural grassland
3.2.2. Moors and heathland
3.2.3. Sclerophyllous vegetation
3.2.4. Transitional woodland/shrub
To the above we added “2.2.3 Olive groves,” based on our experience that low input olive groves are potentially rich orchid habitats, as long as they are not plowed or sprayed with herbicides (Katsalirou et al., 2016b), though olive groves covered very little of the study area.
Subsequently, we assigned a unique numerical ID to each of 172 square cells within the narrow study area that were covered by at least 50% by one or more of the aforementioned CLC classes. Of the 172 cells, 25 were randomly selected by a random number generator (Numbers v. 10.3.9, Apple Inc.). Figure 4 depicts the final sampling design implemented during the main phase.
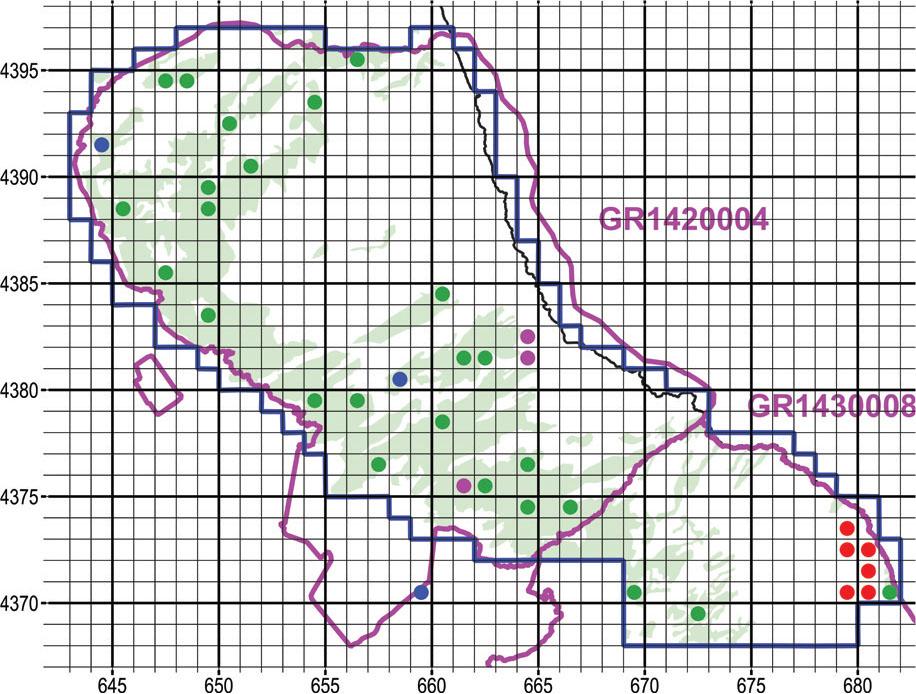
Map of the sampling design during the main sampling phase, i.e., fall of 2021 (for Spiranthes spiralis) to spring of 2022. Olive green fill: Corine Land Cover classes that correspond to open habitats, i.e., a combination of scrub and/or herbaceous vegetation. Blue dots: Locations suggested by the supervisory authority. Magenta dots: Locations with Ophrys scolopax subsp. cornuta by Biel & Rudolph (1992). Red dots: Locations likely to host Orchis provincialis based on the preliminary scouting. Green dots: Sampling units selected at random. UTM grid spacing 1 km.
Slika 4: Zemljevid prikazuje način vzorčenja med glavno fazo, to je od jeseni 2021 (za vrsto Spiranthes spiralis) do spomladi 2022. Olivno zelena: razredi Corine Land Cover, ki predstavljajo odprte habitate, to je kombinacijo grmiščne in/ali zeliščne vegetacije. Modre točke: lokacije, ki so jih predlagale nadzorne oblasti. Škrlatne točke: lokacije z Ophrys scolopax subsp. cornuta iz publikacije Biel & Rudolph (1992). Rdeče točke: lokacije, na osnovi predhodnega pregleda terena, kjer se verjetno pojavlja Orchis provincialis. Zelene točke: naključne vzorčne točke. Velikost kvadrantov UTM mreže je 1 km.
The preliminary reconnaissance phase lasted from 25.04.2021 to 04.08.2021, with diminished findings toward the end of this period.
Spiranthes spiralis flowers from August to October throughout its broad distribution range, from the Atlantic coasts of Europe to Iran and from S Sweden to N Africa (Delforge, 2006). In the Mediterranean, we would expect the onset of flowering to coincide with the onset of the wet season, which varies from year to year. In our case, we found that the optimum time window was from 20.10.2021 to 13.11.2021.
The main sampling phase for the spring-flowering species lasted from 16.02.2022 to 04.07.2022, with diminished findings toward the end of this period. A few supplementary observations were taken on 19.04.2023 and 20.04.2023.
At each location we photographed the specimens and filled forms with data such as names of investigators, date, nearest settlement, place name, plant community, names of taxa, number of specimens, development stage, coordinates, altitude, parent rock, soil type, nearby plant species, and main anthropogenic threats.
In total, 129 square cells 1 km2 wide were explored, mostly within the two Natura 2000 sites, of which 100 yielded orchid findings (Figure 5). The tally of sampled cells far exceeds the target of 37 that was decided during the design of the main sampling phase. This is because the tally includes:
- (i)
cells sampled during the preliminary reconnaissance phase, 25.04.2021 to 04.08.2021
- (ii)
circumstantial samplings, i.e., promising biotopes explored on the way to and from the 37 designated sampling units
- (iii)
biotopes sampled during the off-hours of the field task force
Map with sampling units 1 km wide that yielded orchid findings within the broader study area during 2021–2022. With magenta outline the two Natura 2000 sites, with dark blue the outline of the narrow study area. UTM grid spacing 1 km.
Slika 5: Zemljevid kvadrantov velikosti 1 km, kjer smo v letih 2012–2022 našli orhideje v večjem preučevanem območju. S škrlatno barvo sta označeni območji Natura 2000, s temno modro pa ožje preučevano območje. Velikost kvadrantov UTM mreže je 1 km.
The descriptions of the sites are in Appendix I. The total number of sites listed are 107, as some sampling units contained more than one distinct biotope that have been described separately.
In total, we identified 39 orchid taxa (species and sub-species) plus two nothotaxa (Table 1, nomenclature according to POWO, 2024).
During the final stage of the manuscript, we received notice of the new edition of the Atlas of the Hellenic Flora (Strid, 2024). As with the Atlas of Greek Orchids (Antonopoulos & Tsiftsis, 2017; Tsiftsis & Antonopoulos, 2017), the scale of the maps is too coarse to pinpoint the exact location of the findings. Fortunately, the author was so kind to share the records from the Flora Hellenica Database on which the maps of the Atlas are based (Prof. Arne Strid, personal communication 14 May 2024).
Before comparing our findings with the Flora Hellenica Database, we processed its records as follows:
- (i)
We omitted records with only approximate location. For example, records in the Database originating from the Atlas of Greek Orchids were read off dot maps of 10 km precision, not useable for our purposes.
- (ii)
We improved and supplemented the records originating from the survey of Biel & Rudolph (1992). The Flora Hellenica Database listed only approximate locations read off dot maps. Since we had access to the original hard copy, we replaced the approximate locations with locations of 0.1 km precision and added several missing records.
The useable records from the Flora Hellenica Database that fall within our narrow study area, besides those contributed by Biel & Rudolph (1992), are mostly older observations contributed by Thomas Raus (Raus 1979b, 1980) and Eckhard Willing (Willing & Willing 2003, 2009). The comparison of our findings to the Flora Hellenica Database clearly demonstrates that there are extensive tracts of land that our task force explored for the first time (Figure 6). Further, the Flora Hellenica Database reports only 20 of the 33 taxa that we recorded within the narrow study area (Table 1, names in bold). One explanation is the strong random component of our sampling design, which included tracts of the study area that are normally inaccessible.
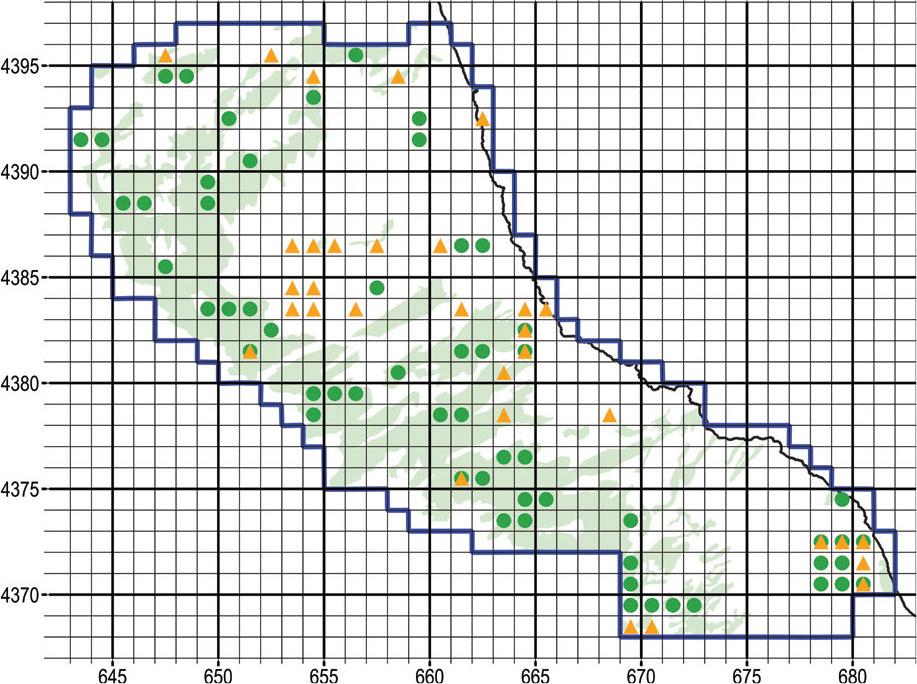
Map with square cells 1 km wide that yielded orchid findings within the narrow study area during 2021–2022. Green dots represent our findings, orange triangles represent records from the Flora Hellenica Database, mostly predating 1992. UTM grid spacing 1 km.
Slika 6: Zemljevid kvadrantov velikosti 1 km, kjer smo v letih 2012–2022 našli orhideje v večjem preučevanem območju. Zelene točke predstavljajo naše najdbe, oranžni trikotniki so podatki iz Flora Hellenica Database, predvsem pred letom 1992. Velikost kvadrantov UTM mreže je 1 km.
It is interesting that Orchis simia was missed by previous surveys although it occurs quite abundantly in its respective habitats. We hypothesise that decades of deforestation and intensive grazing have thinned out formerly forested biotopes, thus attracting photophilic species for the first time.
On the other hand, the Flora Hellenica Database reports nine taxa within our narrow study area that we did not find: Anacamptis coriophora, Epipactis helleborine, Gymnadenia conopsea, Himantoglossum calcaratum subsp. rumelicum (syn. H. jankae), Neotinea ustulata, Neottia ovata, Ophrys sphegodes subsp. spruneri, Serapias bergonii (sub S. vomeracea subsp. laxiflora), and S. lingua. Every taxon that we missed is reported from only one location each, indicating that their presence within the narrow study area is rather uncommon. As far as Epipactis helleborine and Neottia ovata are concerned, our sampling scheme heavily favoured open ecosystems of relatively low vegetation, whereas E. helleborine and N. ovata prefer dense forests made up by Quercus ilex, Castanea sativa and Fagus sylvatica (Raus, 1979b, 1980); in fact, we were able to confirm sizeable populations of E. helleborine further S, in chestnut and beech forests.
To improve the conservation status of the family Orchidaceae, we propose a series of conservation measures.
Herds of wild pigs and their hybrids with domesticated pigs roam freely on Mounts Pelion and Mavrovouni (Figure 7). Feral pigs represent a major threat to biodiversity as they dig up extensive tracts of land, uprooting the herbaceous vegetation (Figure 8). We frequently witnessed orchid tubers unearthed by pigs (Figure 9). It is not known whether pigs forage on orchid tubers or whether their destruction is collateral damage during the search for some other food.

Fearless of humans, feral swine dig up important orchid biotopes at site Psili Rachi I on Mount Mavrovouni.
Slika 7: Divji prašiči, ki se ljudi ne bojijo, so prekopali pomembna rastišča orhidej na lokaciji Psili Rachi I na gori Mavrovouni.

Feral swine devastate large tracts of land near the hamlet of Ano Kerasia on Mount Pelion. In the fall of 2021, the supervisory authority tipped us off about a sizeable population of Spiranthes spiralis. By the time we got there, the site was all dug up.
Slika 8: Divji prašiči so uničili velike površine zemljišč blizu naselja Ano Kerasia na gori Pelion. Jeseni 2021 so nas odgovorni obvestili o veliki populaciji vrste Spiranthes spiralis, vendar je bilo rastišče do našega prihoda že razrito.

Specimens of Orchis italica dug up by feral pigs at site Metochi of S Magnesia.
Slika 9: Primerki vrste Orchis italica, ki so jih izkopali divji prašiči na lokaciji Metochi v južnem delu območja Magnesia.
Besides feral pigs, we encountered herds of domesticated, free-range pigs, that similarly cause major damage to the ecosystem. We witnessed sizeable herds roaming unattended in the forest ecosystem (Figure 10).

Domesticated, free-range pigs graze unattended at site Kalamaki of Larissa.
Slika 10: Udomačeni prašiči, na lokaciji Kalamaki v območju Larissa, ki se prosto pasejo.
Recognising the problem, the national authorities have instituted an emergency measure to contain the over-population of feral swine invading rural and suburban areas. The so-called “persecution crews,” essentially ad hoc extermination squads made up of licensed hunters, are supposed to hunt feral pigs. This measure is in addition to the harvest quota allowed hunters during hunting season, as it can be invoked by local authorities at any time throughout the year. Whether this measure is effective, it remains to be seen. We predict that, because the measure is implemented on a voluntary basis, without financial incentive, its success will be limited. In fact, offering a financial incentive could be worse, as part of the public may object to tax money subsidising the slaughter of animals. In addition, there is certainly a liability issue related to encouraging consumption of game meat that has not been inspected by veterinarians. Feral swine are vectors of numerous diseases and parasites infecting livestock and humans (Finzel & Baldwin, 2015).
Instead of invoking piecemeal measures such ad hoc “persecution crews,” whose success largely depends on the goodwill of volunteers, the most successful feral swine management strategies employ a diversity of tactics in a comprehensive, integrated approach. Some solutions tried in other parts of the world, other than shooting, are trapping (Sharp, 2012) and installation of feeders delivering contraceptives (National Hog Farmer, 2021). We think that restoring populations of natural predators should be considered as an additional measure within the framework of an integrated wildlife management strategy.
We witnessed intensive grazing by small ruminants (sheep and goats) as well as bovines within the narrow study area, especially on Mount Mavrovouni. The damage by ruminants is twofold: (i) direct consumption of the aboveground plant parts before they set seed, and (ii) inadvertent trampling during the search for some other food (Figure 11). Administratively, the narrow study area belongs almost entirely to the Municipalities of Ayia, Riga Ferreou, and Zagora-Mouresi. From aggregate statistics of the Payment and Control Agency for Guidance and Guarantee Community Aid (2024) we tallied the populations of small ruminants and bovines by municipality. Of the bovines, 98–99% are beef cattle, mostly free-range. The results for the three Municipalities of interest are mapped in Figure 12. The Municipalities of Ayia and Riga Ferreou are grazed by several thousand small ruminants and a few thousand cattle; this is consistent with our observations of severely degraded ecosystems in grazed lands. On the other hand, in the Municipality of Zagora-Mouresi, where almost no animal husbandry is practised, we have not observed degradation of ecosystems due to grazing. Unfortunately, we could not find data tallied by Municipal Unit (a subdivision of Municipality) that would yield a more accurate picture.

Beef cattle force their way through the pseudo-maquis, trampling numerous orchids in the process. Site Agrioachladia on Mount Mavrovouni.
Slika 11: Govedo, ki se prebija skozi pseudomakijo, potepta številne orhidje. Lokacija Agrioachladia na gori Mavrovouni.
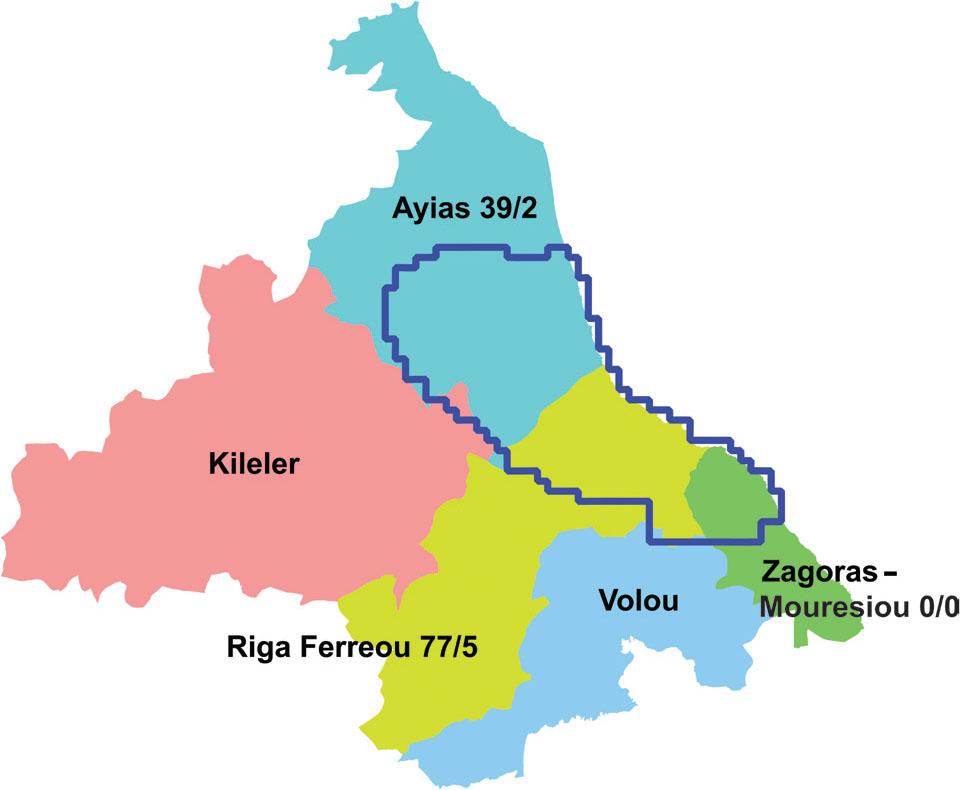
The narrow sampling area, outlined in dark blue, is shared among the Municipalities of Ayia, Riga Ferreou, and Zagora-Mouresi. The numbers after each Municipality are tallies of small ruminants/bovines, in thousand heads.
Slika 12: Ožje preučevano območje, označeno s temno modro, sega v občine Ayia, Riga Ferreou in Zagora-Mouresi. Številke ob imenu občine so seštevki malih prežvekovalcev/goveda v tisočih glav.
It is difficult to propose management measures for grazing. A major obstacle is the institutional framework that governs EU subsidies for animal husbandry that does not hold stockbreeders accountable for their practices. Most livestock operations are kept afloat with generous injections of subsidies, without regard to their inherent economic viability or environmental sustainability (Lorent et al., 2008; Tsiouni et al., 2021). The root cause of the problem is a notorious algorithm concocted by national authorities titled the “technical solution.” Simply put, it means that stockbreeders earn subsidies based on a hypothetical apportionment of a national inventory of rangelands that may be located several hundred kilometres away, rather than the actual lands they graze. Because the subsidies are disjointed from the actual land they graze, stockbreeders are not responsible for sound land management practices.
At the very least, we propose delimiting biotopes of high orchid diversity and enact controls on grazing, so that at least some of the populations are protected. Our findings can be used as a starting point to define “islands” of high orchid diversity that are worth conserving, based on the relative rarity and conservation interest for each species.
Nevertheless, we do not advocate a general ban on grazing, as grazing is not without its merits:
- (i)
Owing to deruralisation, declining human fertility, and subsequent human population decline, much arable land in the countryside has been abandoned. Most orchid species are photophiles, meaning that they thrive in open ecosystems with sparse woody vegetation, such as thin maquis, phrygana, and rangeland. Grazing may contribute to preserving these habitats by preventing afforestation.
- (ii)
Our research has demonstrated that moderate grazing may preserve and even enhance the soil microbial functions that promote soil nutrient cycling and, by extension, soil health (Katsalirou et al., 2010; Katsalirou et al., 2016a).
In summary, while overgrazing must be contemplated as a major driver for declining orchid diversity, moderate grazing may be beneficial to orchid conservation. Therefore, in areas of high orchid diversity we propose a ban on grazing during the reproductive stage, from budding to seed maturity, which for most species spans the interval from March to June. In areas where Spiranthes spiralis abounds, we propose a ban on grazing from October to December.
To ensure a long-term supply of genetic material for future in vitro propagation efforts, we propose the geographic delineation of genetic micro-reserves for the five taxa of conservation interest. These micro-reserves must be fenced with solid fences to resist grazing animals. Based on our findings, we propose the following locations:
- (i)
Ophrys scolopax subsp. cornuta: Although it is not an altogether unusual taxon within the narrow study area, we did not find sizeable populations. The largest population was recorded at location Petralona (site 13 in Appendix I).
- (ii)
Ophrys umbilicata subsp. umbilicata: The largest population was recorded near Lake Karla (site 26 in Appendix I). Currently, it is under direct and constant threat by a nearby sheep farm.
- (iii)
Orchis provincialis: Most locations are unsuitable as genetic reserves because they are within inhabited areas or logged forests with frequent traffic. The best option would be location Stavros II (site 9 in Appendix I).
- (iv)
Orchis simia: The largest population was recorded at location Agrioachladia (site 24 in Appendix I). It is under direct and constant threat from grazing and trampling by free-range cattle.
- (v)
Spiranthes spiralis: Although it is not an altogether unusual taxon within the narrow study area, we did not find sizeable populations. The largest population was recorded at location Skiti II (site 19 in Appendix I).
Road verges are important wildlife corridors linking habitats. Throughout much of the world, road banks and medians are respected as valuable genetic reserves of native flora. If managed correctly, road verges can support remarkable collections of orchids. In Mounts Pelion and Mavrovouni, spectacular stands of Orchis italica and various Ophrys adorn the road banks in springtime. One reason is that roads interrupt the thick, impenetrable forest allowing sunlight to reach the ground. Roadside verges are sanctuaries where orchids “escape” from intensive farming and grazing. Proper management involves simply doing less: Give the roadside vegetation some space, avoid herbicides, and allow plants to set seed before mowing (Fekete et al., 2017).
Unfortunately, local authorities in charge of roadside maintenance are not educated about this valuable genetic reserve. Along the Aegean coast of Mount Pelion, we frequently observed maintenance crews indiscriminately mowing the roadsides in early summer in preparation for the tourist season.
On some paved roads, such as from Chania to Kissos, clearcutting a wide strip on either side of the road is unnecessary; a shoulder has been provided between the pavement and the kerb that doubles as an emergency lane (Figure 13). Mowing the vegetation well beyond this shoulder does not confer any additional benefit to traffic safety. The banks of this road are rich in species such as Dactylorhiza saccifera and Epipactis microphylla that should be protected at least until seed maturity.

The verges on either side of the road from Chania to Kissos on Mount Pelion are rich in orchids such as Dactylorhiza saccifera and Epipactis microphylla. A lone inflorescence of D. saccifera escaped mowing.
Slika 13: Na cestnih brežinah na obeh straneh ceste od naselja Chania do naselja Kissos na gori Pelion, so bogata nahajališča orhidej, kot sta Dactylorhiza saccifera in Epipactis microphylla. Le eno socvetje D. saccifera ni bilo pokošeno.
A different situation arises along the narrow, convoluted roads that connect the traditional villages of Pelion along the Aegean coast, such as from Zagora to Chorefto. There, the width of the pavement is just enough for two cars, and sometimes not even that. While in this case mowing of the road banks is unavoidable to maintain traffic safety, it should be delayed as much as possible, ideally after most orchids have set seed, and certainly not before 30 June.
Finally, along the network of logging trails on the NE slope of Mounts Pelion and Mavrovouni, mowing of the trail banks can be delayed until well after 30 June, as they do not serve tourist traffic.
Natural chestnut forest can be rich in orchids, as our preliminary reconnaissance from the Aegean coast of Mount Pelion attests. Chestnut forest often is sparser than beech, Hungarian oak, or maquis, so that more sunlight reaches the ground. Encouraged by abundant findings in natural chestnut woods, our field task force decided to expand its search to cultivated chestnut groves on Mount Mavrovouni, E of the village of Potamia. During the preliminary reconnaissance phase, we traversed eight square cells 1 km2 wide (Figure 14).
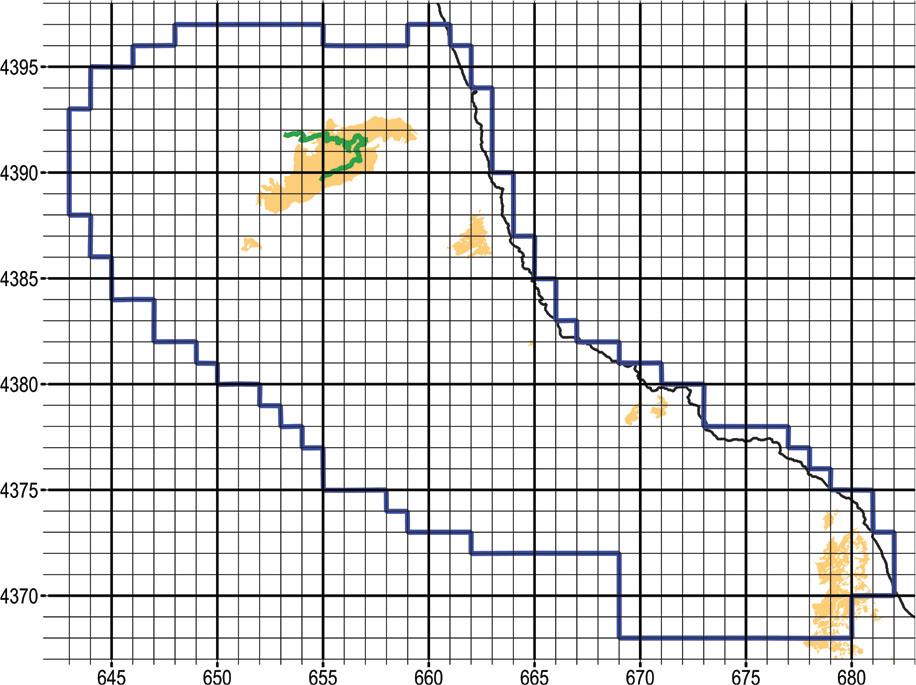
Orange fill indicates chestnut woods (natural and cultivated) within the narrow study area. Since Corine Land Cover is not so detailed, photo-interpretation was provided by the Management Unit of Protected Areas of Thessaly. We surveyed along the green trail that crosses the expansive chestnut plantations E and S of the village of Potamia. UTM grid spacing 1 km.
Slika 14: Oranžna barva označuje kostanjeve gozdove (naravne in kultivirane) v ožjem preučevanem območju. Ker Corine Land Cover ni tako natančen, smo uporabili foto-interpretacijo, ki nam jo je posredovala Uprava zavarovanih območij Tesalije. Terenski pregled smo opravili ob zeleni poti, ki prečka kostanjeve nasade vzhodno in južno od vasi Potamia. Velikost kvadrantov UTM mreže je 1 km.
Our survey yielded zero orchid findings, a sobering realisation give the overall high orchid diversity on Mount Mavrovouni. We soon realised that, in cultivated chestnut groves, the understory is meticulously mowed, while pesticides are liberally applied with tractor-drawn air-blast sprayers. Under these conditions, survival of orchids – or any herbaceous vegetation for that matter – is questionable. We recommend that mowing of the understory is delayed until after seed set, certainly after 30 June. Meanwhile, we recommend organic chestnut culture as an alternative method to help restore native biodiversity.
The N part of the narrow study area that is flatter, and the SE part that is centred around the village of Pouri, are largely covered by intensely cultivated apple orchards that may intermingle with patches of natural vegetation. The corresponding Corine Land Cover classes are “2.2.2 Fruit trees and berry plantations” and “2.4.3 Land principally occupied by agriculture, with significant areas of natural vegetation” (Figure 15).
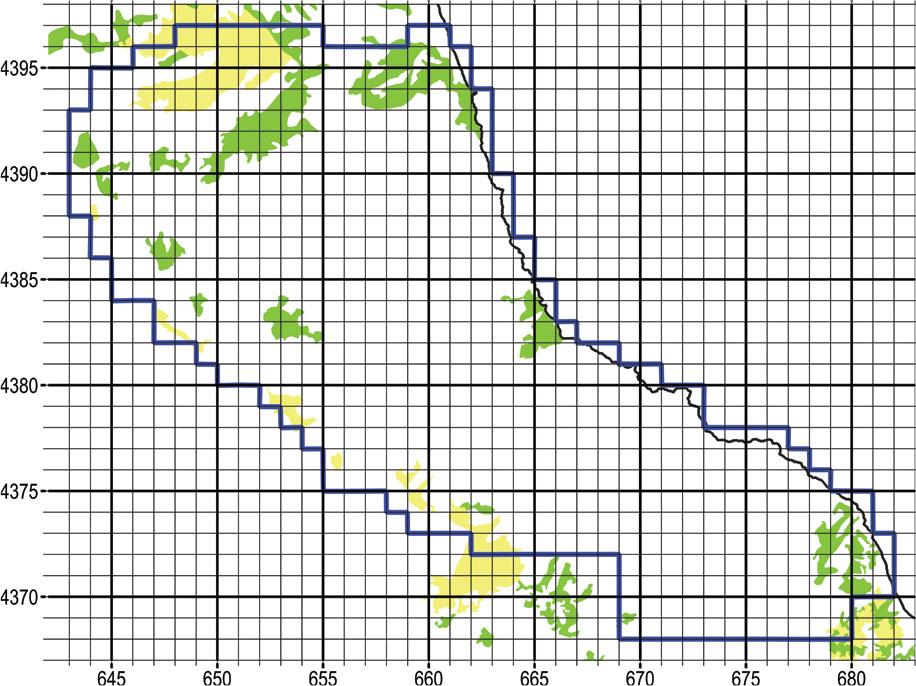
Yellow fill indicates Corine Land Cover class “2.2.2 Fruit trees and berry plantations”. Green fill indicates class “2.4.3 Land principally occupied by agriculture, with significant areas of natural vegetation.” At the N and the SE part of the narrow study area, these two classes correspond primarily to apple orchards that may be interspersed with patches of natural vegetation. UTM grid spacing 1 km.
Slika 15: Z rumeno je označena Corine raba tal “2.2.2 Sadovnjaki in nasadi jagodičja”. Zelena predstavlja rabo “2.4.3 Kmetijska zemljišča z večjim deležem naravne vegetacije”. Na severnem in jugovzhodnem delu ožjega preučevanega območja ta tip rabe predstavljajo predvsem sadovnjaki jablan, ki se izmenjujejo s pasovi naravne vegetacije. Velikost kvadrantov UTM mreže je 1 km.
In apple orchards, the ground is meticulously mowed, while pesticides are liberally applied with tractor-drawn air-blast sprayers. Under these conditions, the occasional orchids find refuge within the remnants of native forest (Figure 16). We recommend that borders of native vegetation are preserved around the apple orchards. We recommend organic apple culture as an alternative method to help restore native biodiversity.

Hillslopes around the village of Pouri largely covered by intensive apple orchards. The occasional orchids find refuge within patches of native chestnut forest (top).
Slika 16: Pobočja pri vasi Pouri, kjer so intenzivni nasadi jablan. Orhideje občasno uspevajo v zaplatah naravnega kostanjevega gozda (zgoraj).
A sizeable population of Ophrys umbilicata subsp. umbilicata, one of the taxa of conservation interest, was located atop the hill NE of the village of Neochori (Plasia) of Larissa, along with specimens of Ophrys sphegodes subsp. taurica, Ophrys tenthredinifera, and Serapias vomeracea, altogether comprising an important island of orchid diversity on a hill surrounded on three sides by intensive farming.
We recorded the population of Ophrys umbilicata subsp. umbilicata on 04.04.2022. By 16.05.2022, already land surveyors were mapping the hillside. By 29.06.2022, the first solar panel mounts were driven to the ground. In the process, we witnessed the land being meticulously cleared with fire and bulldozers. Currently, a large photo-voltaic park covers the hillside (Figure 17).

A large photovoltaic park on the hill NE of the village of Neochori (Plasia) of Larissa, once a rich orchid biotope. The native vegetation has been cleared with fire and bulldozers.
Slika 17: Velik fotovoltaični park na SV hribu pri vasi Neochori (Plasia) v območju Larissa, kjer je bilo nekoč z orhidejami bogato rastišče. Naravno vegetacijo so odstranili z ognjem in mehanizacijo.
Projects of such scale effect a dramatic change in the landscape and incur massive loss of biodiversity that does not seem to matter to policy makers, despite that the orchid family has been afforded institutional protection. Most solar parks are so remote that they encounter little reaction by the public; according to the 2011 census, the village of Neochori counts 41 residents. Besides, the public has been convinced that renewable energy is good for the planet. Actually, these projects stretch the very notion of “renewable,” in the sense that the energy itself may be renewable yet the ecosystem is irrevocably damaged. At the very least, mitigating actions should be considered. A small proportion of the budget of such projects could be allocated to actions for recording, preserving, and restoring native biodiversity in disturbed ecosystems.
On 5 Sep. 2023, storm Daniel, the fiercest Mediterranean hurricane in recorded history, hit the study area with full force. Severe erosion and mudslides occurred throughout the study area. It will take years to assess the damage to natural resources. We hope that our study will serve as baseline to assess the effects of this extreme weather event in the years to come.
During the fieldwork of the present project 129 square cells 1 km2 wide were explored, mostly within the Natura sites of Mounts Pelion and Mavrovouni, altogether comprising 796 km2. Of the sampled cells, 100 yielded orchid findings. In total, 39 orchid taxa (species and subspecies) plus two nothotaxa were recorded. While both Natura sites were explored, the sampling effort was intensified within a narrow study area of 550 km2 encompassing the N part of Mount Pelion and the whole of Mount Mavrovouni. In the narrow study area, 33 orchid taxa and nothotaxa were recorded. Of those, 13 constitute new additions to the Flora Hellenica Database: Anacamptis papilionacea subsp. papilionacea, A. pyramidalis, Himantoglossum robertianum, Ophrys × delphinensis, O. sicula (syn. O. lutea subsp. galilaea), O. speculum subsp. speculum, O. sphegodes subsp. sphegodes (syn. O. hebes), O. tenthredinifera, O. tenthredinifera × O. sphegodes subsp. taurica, O. umbilicata subsp. umbilicata, Orchis italica, O. simia, and Platanthera bifolia.
The main environmental threats facing orchids are rooting by feral and domesticated free-range pigs, over-grazing by ruminants, clearing of road verges, intensive chestnut and apple culture, and the encroachment of photovoltaic farms. A novel threat is extreme weather events such as storm Daniel. We recommend containment of feral and domesticated free-range pigs, limitations on grazing by ruminants, the establishment of micro-genetic reserves in orchid-rich areas, restrictions on mowing of the road verges until after the orchids have set seed, promoting organic chestnut and apple culture, and measures mitigating the impact of massive photo-voltaic projects.