
Rubus L. is one of the rather large and widespread genera of the Rosaceae family and is represented predominantly by erect to trailing shrubs or scramblers, rarely by herbs (Takhtajan, 2009). The genus is economically and ecologically important as fruit crops, ornamental plants, and as pioneers in early forest succession (Alice & Campbell, 1999). It has also been revealed that Rubus readily invades natural areas (Daehler, 1998; Alice & Campbell, 1999). At the same time when exploring the pollen characteristics of the invasive in Poland anthropophyte species R. laciniatus, Lechowicz et al. (2021) noted that pollen morphology probably does not affect the invasive traits of this species. Rubus species were a food source and medicinal plants for native peoples soon after the Ice Age (Connolly, 1999), and the medicinal use of brambles was documented in the writings of Aeschylus and Hippocrates, between 500 to 370 BCE (Hendrickson, 1981; Hummer, 2010). It has been suggested that within subfamily Rosoideae Martinov the crown ages of its six tribes (including tribe Rubeae Dumort.) range from 45.02 to 10.42 Ma (the middle Eocene to the late Miocene) (Chen et al., 2020). According to Lu (1983, p. 25): “It is clearly shown that center of distribution lies in North America at present time”, from where the last common ancestor migrated to Central and South America, and Europe and Asia (Carter et al., 2019).
According to various sources, the genus includes 250 to 900 + species, with around 750 species that are documented in Europe (Kurtto et al., 2011; Mabberley, 2017; Huang et al., 2023). Brambles exhibit a high level of morphological diversity and are often characterized by hybridization, frequently associated with polyploidy and apomixis (Alice, 2002; Sochor et al., 2015). The basic chromosome number is x = 7, and the genus has been reported to have different levels of ploidy in nature including 2x–18x as well as aneuploid genomes. Both natural and human-made hybrids are prevalent (Nybom, 1986; Foster et al., 2019; Gao et al., 2023). Specifically, “… except for a few sexual species occurring in Europe and its nearest vicinity… the subgenus Rubus is exclusively represented on the continent by polyploids of hybrid origin (2n = 21, 28, 35, and 42), where sexual reproduction has been almost entirely supplanted by apomixis” (Zieliński, 2004, p. 1).
In some classification systems, Rubus is categorized under the subfamily Ruboideae Thomé (Takhtajan, 1997, 2009) and the tribe Rubeae (Angiosperm Phylogeny Website, https://www.mobot.org/mobot/research/apweb/), among others. The global taxonomic classification of the genus was established early in the 20th century, leading to the definition of 12 subgenera (Focke, 1910, 1911, 1914). However, a recent phylogenetic study of 145 Rubus species, including cultivars and hybrids, identified 10 subgenera (Huang et al., 2023). This study found that the genus is not monophyletic; only R. subgenus Anoplobatus Focke forms a monophyletic group, while the remaining nine subgenera are either para- or polyphyletic (Chen et al., 2016; Huang et al., 2023). At the same time, according to Carter et al. (2019) R. subgenus Orobatus (Focke) Focke is also monophyletic.
In Armenia, the total number of Rubus species is 13–14, the vast majority of which belong to the subgenus Rubus (= Eubatus Focke) (Mulkijanyan, 1958), while species R. idaeus L. belongs to the subgenus Idaeobatus Focke. Two rare endemic species, R. takhtadjanii Mulk. and R. zan gezurusMulk., are listed in the Red Book of Plants of Armenia under the EN (endangered) category (Tamanyan et al., 2010). For the species R. takhtadjanii, only one population is known in the Zangezur floristic region (in the vicinity of the villages Tsav and Srashen) on the territory of the Shikakhokh State Reserve and the Plane Grove Nature Reserve. The range of R. zangezurus is fragmented, as the species grows both in the Ijevan floristic region (Berd) and in the Zangezur floristic region (Goris, Tsav) (Hayrapetyan et al., 2017). In the “Habitats of Armenia” (Fayvush & Aleksanyan, 2016) the genus is categorized under F2.33 (Subalpine mixed brushes), F2.338–AM (Subalpine crook stem forest), F3.2477–AM (Dewberry dense scrub), FB.3 (Shrub plantations for ornamental purposes or for fruit, other than vineyards), G1.371–AM (Plane grove in Tsav river valley), G1.927–AM (Aspen groves of North Armenia), G3.4E (Ponto–Caucasian Scots pine forests) and G5.85 (Shrubby clearings).
Pollen morphological features are often used to address complex taxonomical problems regarding interrelationships between various taxa, particularly with reference to the families, subfamilies, tribes, genera, and species. The most distinctive characters include aperture type, exine ornamentation and the size of mature pollen grains. The earliest mention of the peculiarities of Rubus pollen morphology (using LM) can be found in the mid-20th century (Erdtman, 1952). Subsequently, numerous studies using a LM (e.g., Ikuse, 1955; Reitsma, 1966; Kuprianova, 1978) and a SEM (e.g., Hebda & Chinnappa, 1990, 1994; Li et al. 2001; Wronska-Pilarek et al., 2012; Hanchana et al., 2023) have been conducted. In most cases, the authors highlight the size and shape of pollen grains as well as the exine ornamentation as the main diagnostic features. A large-scale study of pollen morphology involving 155 species and 13 variations representing all 12 subgenera of Rubus was carried out by Xiong et al. (2019). The authors noted that pollen cluster analysis tree does not align well with the traditional macromorphological classification and molecular phylogenetic trees based on DNA fragments. Other studies have similarly shown that, in general, pollen characters did not support the currently accepted taxonomic division of the genus Rubus into subgenera, sections and series (Lechowicz et al., 2022), as well as that “… pollen morphology alone is not sufficient to elucidate or reconstruct the taxonomic relationships within Rubus…” (Xiong et al., 2019, p. 705). For that reason, pollen features in Rubus should be treated as auxiliary in taxonomy (Lechowicz et al., 2020).
In Armenia, Avetisyan & Manukyan (1958) studied the pollen morphology of representatives of the genus Rubus using LM. The authors generally characterized the pollen of all investigated species as 3-zonocolporate with large pores and granulate exine ornamentation.
The main purpose of this study was to conduct a comparative palynomorphological analysis of 10 species of the genus Rubus in Armenia using LM and SEM to identify the main diagnostic characters that allow for the differentiation of individual taxa.
Pollen morphological characteristics of 18 samples from 10 species of the genus Rubus in Armenia were analyzed. Unopened mature flower buds were obtained from the herbarium of Institute of Botany after A. Takhtajyan, National Academy of Sciences of Republic of Armenia (ERE).
The descriptions of pollen grains by light microscopy (LM) for each investigated species are based on acetolyzed material (Avetisyan, 1950) and also on grains stained with basic fuchsine (Smolyaninova & Golubkova, 1950) with obligatory fixation of pollen in glycerin jelly.
Six morphological characters, namely length of polar axis (P), length of equatorial diameter (E), length of colpi, apocolpium diameter, mesocolpium width and exine thickness, were measured.
Measurements under LM (AmScope 2000X LED, China, 2015) were taken on 20 pollen grains for each specimen. The studies were carried out at ×200, ×400 and ×1000 magnifications. For descriptions of pollen shape, size, and aperture stained pollen grains were used; the details of exine structure and ornamentation were studied on acetolyzed pollen grains. For scanning electron microscopy (SEM), non-acetolyzed pollen grains were washed with alcohol, placed on a metal stub, and sputter coated with gold (10 nm). Samples were imaged under SEM (JEOL JSM-6390), with a 15 kV electron beam.
Statistical analysis of all studied species was performed using Microsoft Excel with two indicators: ± SD – standard deviation and CV% – coefficient of variation. The sample is weakly variable in case of CV≤10%, with CV from 10% to 20% it is moderately variable, in case of CV≥20% it is being considered as highly variable and with CV≥30%, the highest degree of variability is noted.
For each of the species presented below, depending on the amount of available pollen material, one to four specimens were studied. The list of investigated species and specimens is presented in Table 1; their names fully corresponding to those presented in The Plant List (http://www.theplantlist.org/). Data on some palynological features of the genus Rubus in Armenia are presented in Table 2; the average sizes of pollen grains (μm) of the studied species (P × E) using both our data and information from the literature are shown in Figure 1. The comparison of the average values for some pollen characteristics of the species studied, namely, polar axis (P), equatorial diameter (E), apocolpium diameter and mesocolpium width, was carried out using Simpson and Roe test (Van der Pluym & Hideux, 1997) (Figures 2–5).
List of investigated species and specimens of the genus Rubus (ERE – Herbarium of the Institute of Botany after A. Takhtajyan, National Academy of Sciences, Yerevan, Armenia).
Tabela 1: Seznam preučevanih vrst in primerkov rodu Rubus (ERE – Herbarij Botaničnega inštituta A. Takhtajyan, Nacionalna akademija znanosti, Erevan, Armenija).
| Species | Herbarium sheet information |
|---|---|
| R. armeniacus Focke | ERE, 80977; ERE, 93906 |
| R. caesius L. | ERE, 56383; ERE, 63234; ERE, 63238 |
| R. candicans Weihe ex Rchb. | ERE, 58307; ERE, 63240 |
| R. canescens DC. | ERE, 58846; ERE, 98402 |
| R. cartalinicus Juz. | ERE, 63221 |
| R. caucasicus Focke | ERE, 58313 |
| R. ibericus Juz. | ERE, 135970 |
| R. idaeus L. | ERE, 98394 |
| R. sanctus Schreb. | ERE, 56378; ERE, 56381; ERE, 63978; ERE, 98366 |
| R. takhtadjanii Mulk. | ERE, 64509 |
Some palynological and karyological features of the representatives of the genus RubusL. of the flora of Armenia (± SD – standard deviation, CV% – coefficient of variation, LM – light microscopy, SEM – scanning electron microscopy).
Tabela 2: Nekatere palinološke in kariološke značilnosti predstavnikov rodu RubusL. v flori Armenije (± SD – standardni odklon, CV% – koeficient variacije, LM – svetlobni mikroskop, SEM – vrstični elektronski mikroskop).
| Species | Polar axis (P) (μm) ± SD (CV%) | Equatorial diameter (E) (μm) ± SD (CV%) | P/E | Colpus | Exine ornamentation | Chromosome number (2n) | ||
|---|---|---|---|---|---|---|---|---|
| apocolpium diameter(μm) ± SD (CV%) | mesocolpium width (μm) ± SD (CV%) | LM | SEM | |||||
| R. armeniacus Focke | 23.5–25.8 / 24.5 ± 0.7 (2.7%) | 16.9–22.8 / 19.4 ±1.8 (9.2%) | 8/7–8/6 (subprolate) | 4.8–5.9 / 5.3 ± 0.4 (7.1%) | 8.4–12.5 / 10.4 ± 1.0 (9.8%) | finely reticulate | finely striate-perforate or finely striate-microreticulate | 28 (Krahulcová & Holub, 1997) |
| R. caesius L. | 22.2–28.9 / 25.1 ± 1.8 (7.2%) | 19.7–24.1 / 20.9 ± 1.6 (7.4%) | 8/7–8/6 (subprolate) | 4.5–5.3 / 4.9 ± 0.3 (5.3%) | 14.1–15.8 / 14.9 ± 0.4 (2.8%) | finely reticulate | finely striate-perforate or finely striate-microreticulate | 28 (Krahulcová & Holub, 1997) |
| R. candicans Weihe ex Rchb. | 20.3–25.4 / 22.9 ± 1.9 (8.4%) | 17.5–22.1 / 19.0 ± 1.6 (8.7%) | 8/7–8/6 (subprolate) | 5.0–5.6 / 5.3 ± 0.1 (2.8%) | 14.2–15.4 / 14.8 ± 0.3 (2.0%) | granulate | finely striate-perforate | 21 (Gustafsson, 1943) |
| R. canescensDC. | 23.3–25.8 / 24.5 ± 0.7 (3.0%) | 17.7–20.5 / 19 ± 0.9 (4.9%) | 8/7–8/6 (subprolate) | 4.2–5.1 / 4.7 ± 0.2 (4.8%) (sometimes synaperturate) | 10.5–14.3 / 12.4 ± 1.0 (7.7%) | finely striate reticulate | finely striate-perforate, striae linked together by bridges | 14 (Krahulcová & Holub, 1997) |
| R. cartalinicus Juz. | 21.8–30.1 / 25.8 ± 3.1 (11.8%) | 22.1–23.5 / 22.9 ± 0.5 (2.0%) | 8/8–8/7 (prolate spheroidal) | 2.8–4.5 / 3.6 ± 0.4 (11.6%) (sometimes synaperturate) | 14.4–16.8 / 15.6 ± 0.6 (3.8%) | finely striate reticulate | finely striate-perforate | – |
| R. caucasicus Focke | 21.7–33.2 / 26.3 ± 4.1 (15.7%) | 16.8–22.5 / 18.0 ± 3.6 (19.8%) | 8/6–8/4 (prolate) | 4.7–5.9 / 5.24 ± 0.4 (7.5%) | 18.5–19.5 / 19.0 ± 0.3 (1.7%) | granulate | finely striate-perforate | – |
| R. ibericus Juz. | 20.5–22.5 / 21.4 ± 0.7 (3.1%) | 14.6–18.1 / 16.2 ± 1.2 (7.6%) | 8/6–8/4 (prolate) | 3.5–4.8 / 4.2 ± 0.3 (7.8%) | 11.8–14.6 / 13.2 ± 0.7 (5.3%) | finely reticulate | – | – |
| R. idaeus L. | 18.5–25.3 / 21.8 ± 2.5 (11.6%) | 14.1–18.9 / 16.5 ± 1.6 (9.6%) | 8/6–8/4 (prolate) | 3.8–5.7 / 4.8 ± 0.5 (10.0%) | 10.5–14.9 / 12.7 ± 1.1 (8.7%) | finely reticulate | – | 14, 21 (Chen et al., 1993; Ivanova, 2005) |
| R. sanctus Schreb. | 15.5–33.5 / 19.2 ± 5.3 (27.7%) | 12.1–17.5 / 14.4 ± 1.6 (11.3%) | 8/6–8/4 (prolate) | 4.8–6.1 / 5.4 ± 0.3 (6.0%) | 10.9–12.5 / 11.7 ± 0.4 (3.4%) | granulate | finely striate-perforate, diameter of perforation varies | 14 (Runemark, 2006) |
| R. takhtadjanii Mulk. | 12.9–15.6 / 14.2 ± 0.8 (5.6%) | 14.1–16.8 / 15.4 ± 0.9 (6.1%) | 6/8–8/6 (subspheroidal) | 5.3–6.9 / 6.1 ± 0.4 (6.6%) | 9.8–11.5 / 10.7 ± 0.4 (4.0%) | finely reticulate | – | – |
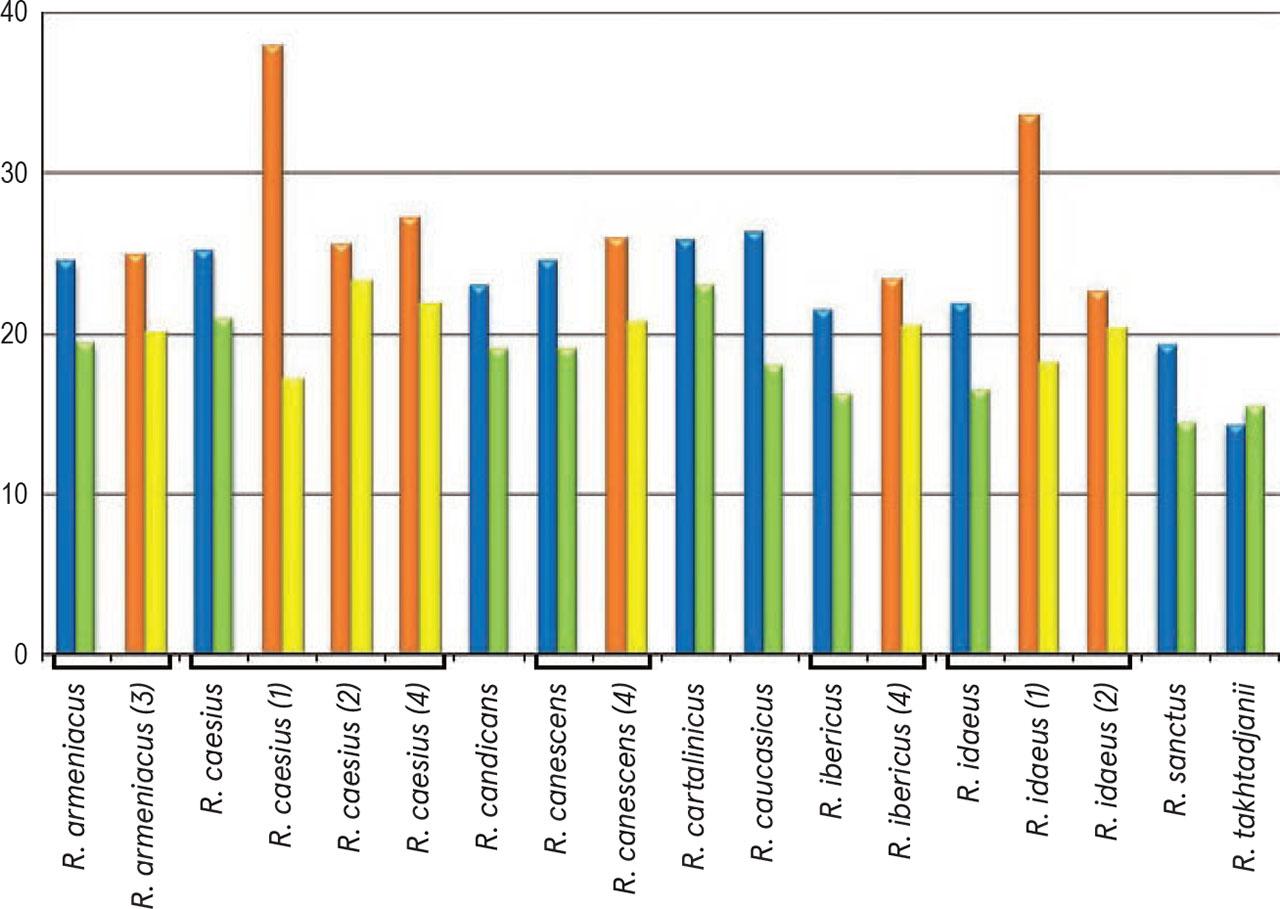
Average sizes of pollen grains (P × E) in some species of the genus Rubus L.
Our data: blue – polar axis (P), green – equatorial diameter (E). Data from literature: orange – polar axis (P), yellow – equatorial diameter (E). (1) – Li et al. (2001), (2) – Lechnovich et al. (2020), (3) – Tomlik-Wyremblewska (1995), (4) – Monasterio-Huelin & Pardo (1995).
Slika 1: Povprečna velikost pelodnih zrn (P × E) nekaterih vrst rodu Rubus L.
Naši podatki: modra – polarna (vertikalna) os (P), zelena – ekvatorialna (horizontalna) os (E). Podatki iz literature: oranžna – polarna (vertikalna) os (P), rumena – ekvatorialna (horizontalna) os (E). (1) – Li et al. (2001), (2) – Lechnovich et al. (2020), (3) – Tomlik-Wyremblewska (1995), (4) – Monasterio-Huelin & Pardo (1995).
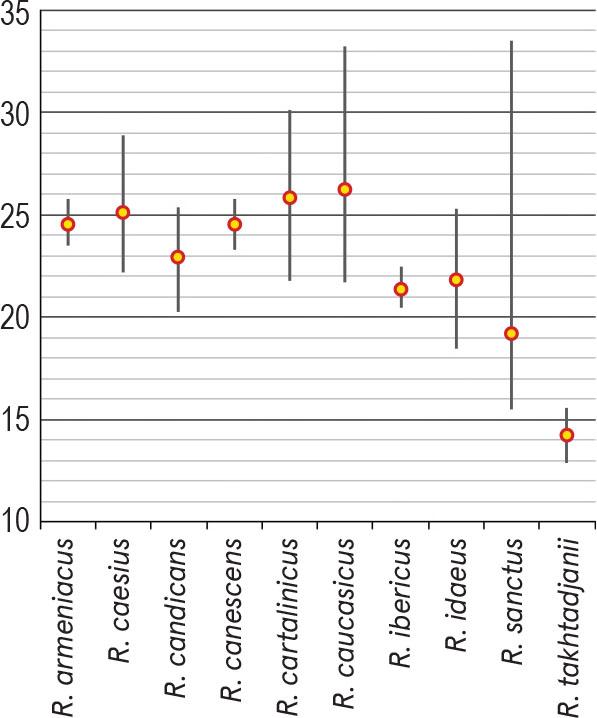
Comparison of the average values of pollen polar axis (P) of investigated Rubus species using Simpson and Roe test.
Slika 2: Primerjava povprečnih vrednosti polarne (vertikalne) osi (P) preučevanih vrst rodu Rubus s Simpsonovim in Roevim testom.
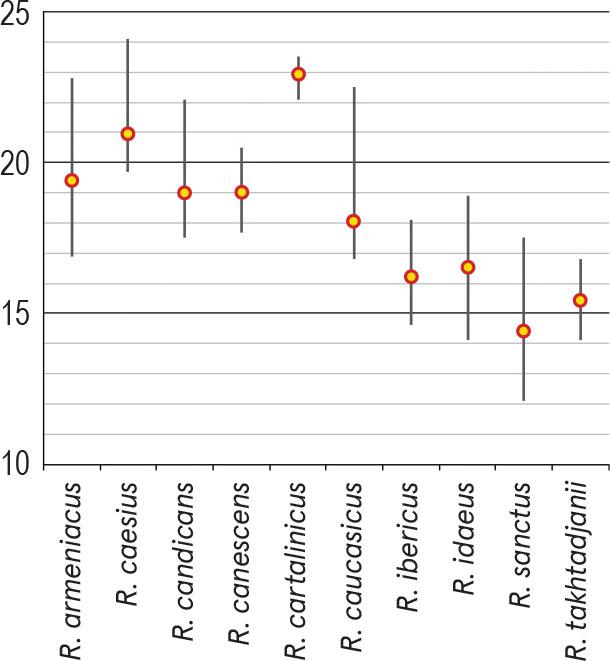
Comparison of the average values of pollen equatorial diameter (E) of investigated Rubus species using Simpson and Roe test.
Slika 3: Primerjava povprečnih vrednosti premera ekvatorialnega premera pelodnega zrna (E) preučevanih vrst rodu Rubus s Simpsonovim in Roevim testom.
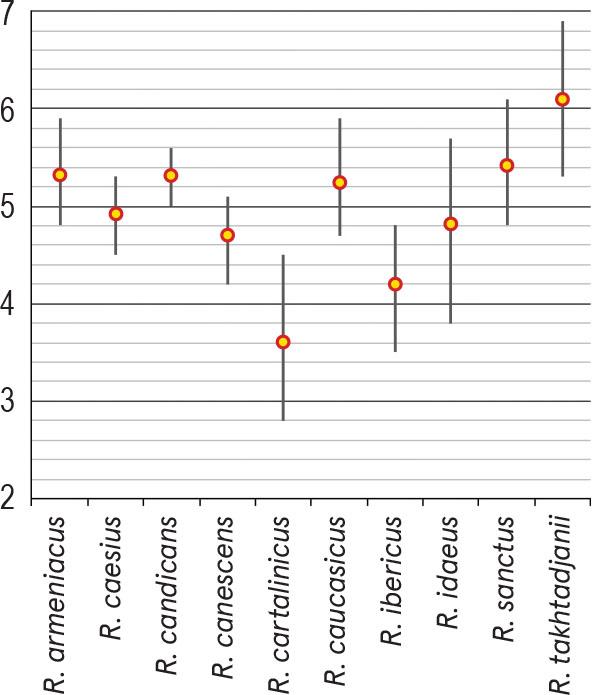
Comparison of the average values of pollen apocolpium diameter of investigated Rubus species using Simpson and Roe test.
Slika 4: Primerjava povprečnih vrednosti premera apokolpija (brazde) preučevanih vrst rodu Rubus s Simpsonovim in Roevim testom.
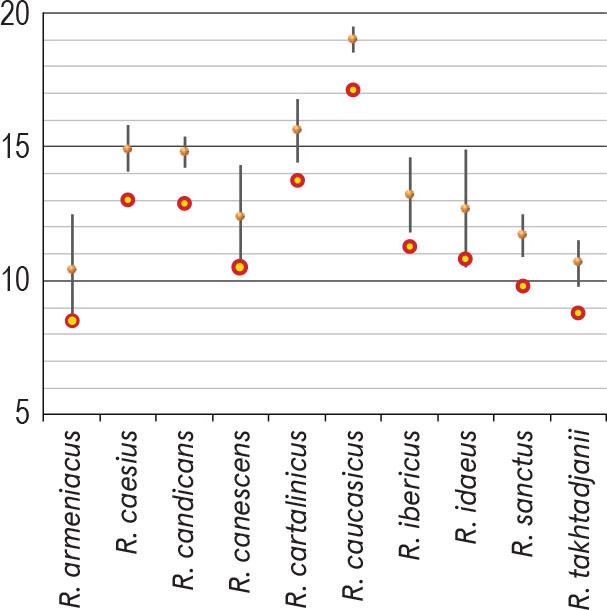
Comparison of the average values of pollen mesocolpium width of investigated Rubus species using Simpson and Roe test.
Slika 5: Primerjava povprečnih vrednosti premera mezokolpija (brazde) preučevanih vrst rodu Rubus s Simpsonovim in Roevim testom.
The classification of the shape and size of pollen grains was given according to Erdtman (1952). The morphological terminology used in our study mainly follows Erdtman (1952), Kuprianova & Alyoshina (1967), Punt et al. (2007), Halbritter et al. (2018), and Hayrapetyan & Bruch (2020).
Karyological studies the genus Rubus in Armenia have not been conducted, and the chromosome numbers for some species, as shown in Table 2, have been obtained from literature sources.
Pollen grains are 3-zonocolp-orate, sometimes 4-zonocolp-orate (Figure 6b), often prolate or subprolate (the shape of pollen grains can vary even within the same sample), sometimes prolate spheroidal in R. cartalinicus and subspheroidal in R. takhtadjanii (Figure 1, Table 2); outline in polar view is rounded-3(4)-angular or rounded-3(4)-lobed; polar axis (P) varies from 12.9 μm (in R. takhtadjanii) to 33.5 μm (in R. sanctus) (Table 2, Figure 2), equatorial diameter (E) from 12.1 μm (in R. sanctus) to 24.1 μm (in R. caesius) (Table 2, Figure 3). Colpi are long or very long (colpus length 4/5–1 of polar axis), occasionally anastomose towards the poles (i.e., synaperturate) (Figure 9b; Figure 10b, c), often geniculate (i.e., with bulge in the equatorial exine of the colpus) (Figure 6f; Figure 8e, g; Figure 9d), from rather narrow to narrow, sometimes almost slit-like (rarely wide), usually with evenly thickened edges (Figure 6d, e; Figure 9e; Figure 10e) and with rounded or pointed ends; colpus membrane ornamentation from psilate to irregularly verrucate (Figure 10a); apocolpium diameter from 2.8 μm (in R. cartalinicus) to 6.9 μm (in R. takhtadjanii) (Table 2, Figure 4), mesocolpium width from 8.4 μm (in R. armeniacus) to 19.5 μm (in R. caucasicus) (Table 2, Figure 5). Ora are spheroidal or oblate spheroidal (Figure 7c; Figure 9c), due to the presence of the geniculum or the convergence of the colpi at the equator, sometimes weakly expressed, rarely with uneven edges and indistinct ends. Exine thickness is 1.2–1.3 μm, columellae separate, with spherical or claviform heads (Figure 6g; Figure 8c). Exine ornamentation is granulate (Figure 7f), reticulate (Figure 6h) or striate-reticulate (Figure 9f) (LM); in the vast majority of the species ornamentation is finely striate-perforate (Figure 8h; Figure 9h; Figure 10g; Figure 11e), sometimes (R. armeniacus and R. caesius) in combination with finely striate-microreticulate one (Figure 6j; Figure 7h) (SEM).
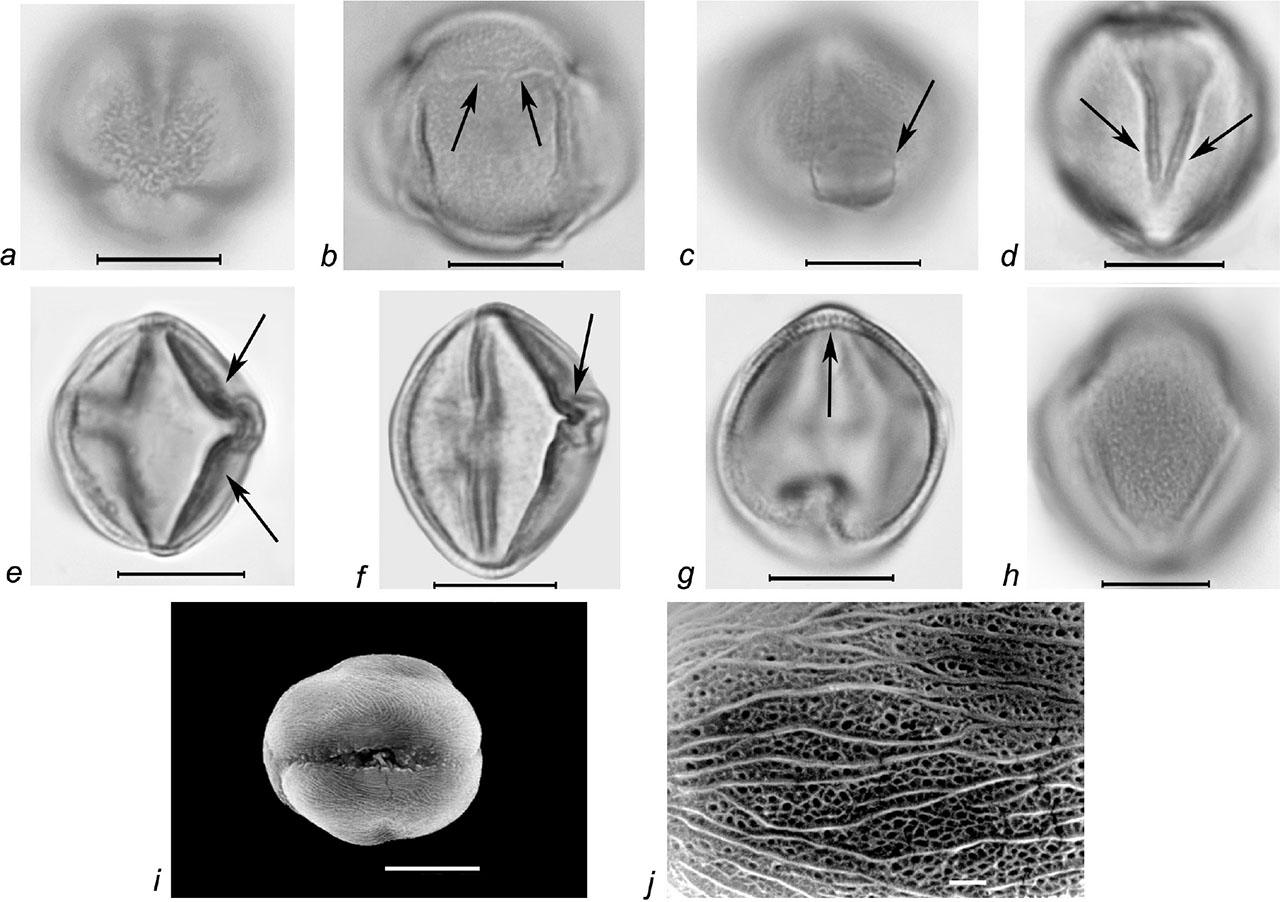
Pollen grains of Rubus armeniacus Focke. a–h – LM micrographs: a – 3-zonocolp-orate pollen grain, b – 4-zonocolp-orate pollen grain with additional small slit-like apertures at the pole (marked with arrows), c – spheroidal os (marked with arrow), d, e – thickened edges of colpi (marked with arrows), f – geniculum (marked with arrow), g – exine, columellae layer (marked with arrow), h – finely reticulate exine ornamentation; i, j – SEM micrographs: i – pollen grain in equatorial view, colpus, j – striate-perforate exine ornamentation in combination with finely striate-microreticulate one; scale bars: a–i – 10 μm, j – 1 μm.
Slika 6: Pelodna zrna vrste Rubus armeniacus Focke. a–h – LM mikrografi: a – 3-zonokolporatna pelodna zrna, b – 4 zonokolporatna pelodna zrna z dodatno odprtino na polih (označeno s puščico), c – sferoidna os (označeno s puščico), d, e – odebeljen del brazde (označeno s puščico), f – genikulum (označeno s puščico), g – eksina, kolumelna plast (označeno s puščico), h – fino mrežasta ornamentacija eksine, i, j – SEM mikrograf: i – pelodno zrno v ekvatorialnem pogledu, brazda, j – črtasto-luknjičasta ornamentacija eksine v kombinaciji s fino črtastomikroretikuliranim okrasjem; merilce: a–i – 10 μm, j – 1 μm.
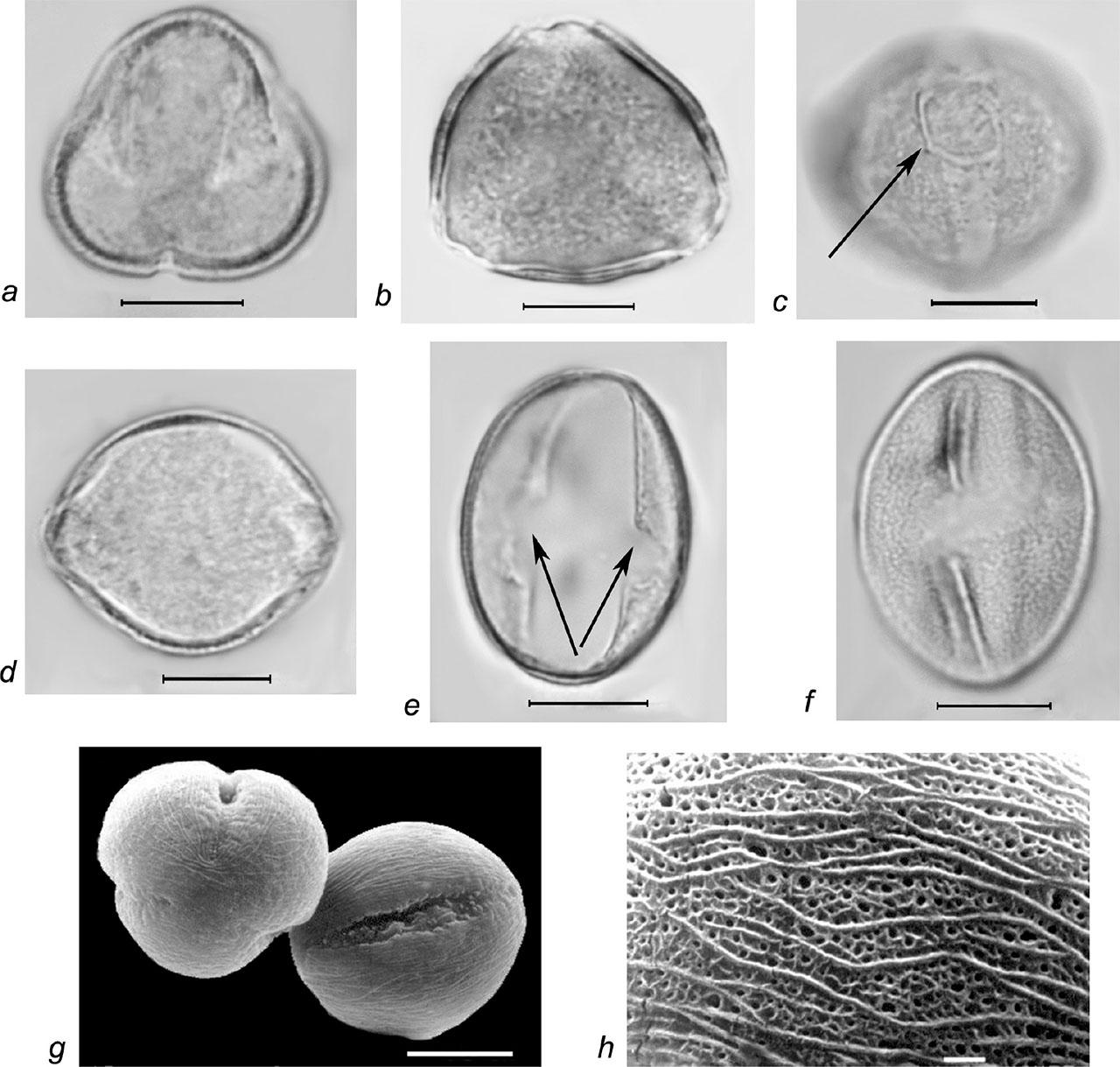
Pollen grains of Rubus caesius L. a–f – LM micrographs: a–b – pollen grains in polar view (a – colpi, b – exine, columellae layer), c – spheroidal os (marked with arrow), d–f – pollen grains in equatorial view (e – os area, marked with arrows), f – granulate exine ornamentation; g–h – SEM micrographs: g – pollen grains in polar and equatorial view, h – striate-perforate exine ornamentation in combination with finely striate-microreticulate one; scale bars: a–g – 10 μm, h – 1 μm.
Slika 7: Pelodna zrna vrste Rubus caesius L. a–f – LM mikrografi: a–b – pelodno zrno v polarnem pogledu (a – brazda, b – eksina, kolumelna plast), c – sferoidna os (označena s puščico), d–f – pelodno zrno v ekvatorijalnem pogledu (e – osno območje, označeno s puščico), f – zrnata ornamentacija eksine, g–h – SEM mikrograf: g – pelodno zrno v polarnem in ekvatorialnem pogledu, h – črtasto-luknjičasta ornamentacija eksine v kombinaciji s fino črtasto-mikroretikulirano ornamentacijo; merilce: a–g – 10 μm, h – 1 μm.
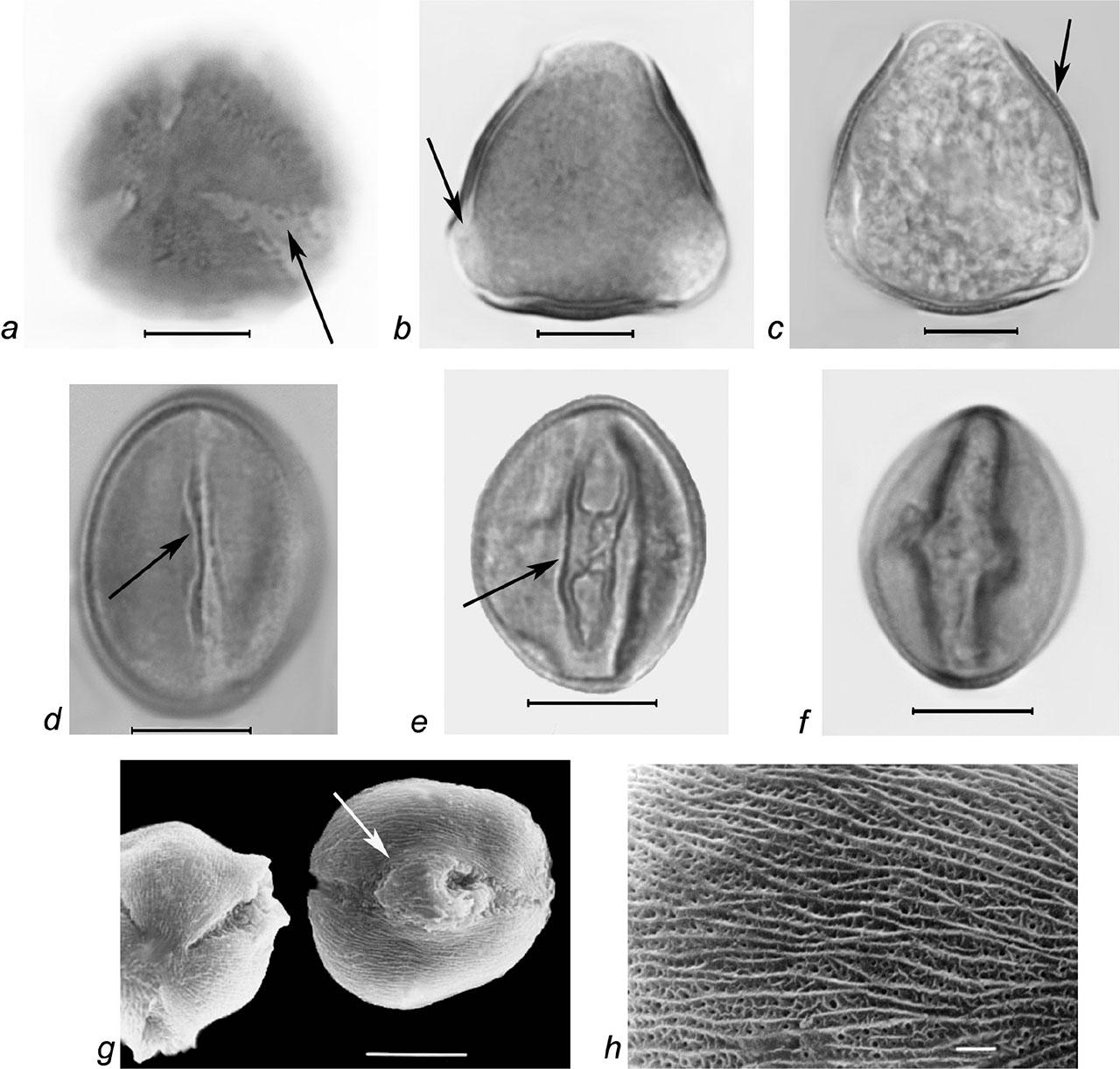
Pollen grains of Rubus candicans Weihe ex Rchb. a–f – LM micrographs: a–c – pollen grains in polar view (a – colpus membrane ornamentation, b – os area, c – exine, columellae layer, marked with arrows), d–f – pollen grains in equatorial view (d – colpus without genuculum, e – colpus with genuculum, marked with arrows, f – thickened edges of colpi); g–h – SEM micrographs: g – pollen grains in polar and equatorial view (colpus with genuculum, marked with arrow), h – finely striate-perforate exine ornamentation (SEM); scale bars: a–g – 10 μm, h – 1 μm.
Slika 8: Pelodna zrna vrste Rubus candicans Weihe ex Rchb. a–f – LM mikrografi: a–c – pelodna zrna v polarnem pogledu (a – ornamentacija membranebrazde, b – območje osi, c – eksina, kolumelna plast, označeno s puščico), d–f – pelodno zrno v ekvatorialnem pogledu (d – brazda brez genikuluma, e – brazda z genikulumom, označeno s puščico, f – odebeljen del brazde); g–h – SEM mikrograf: g – pelodna zrna v polarnem in ekvatorialnem pogledu (brazda z genikulumom, označeno s puščico), h – fino črtasto-luknjasta ornamentacija eksine (SEM); merilce: a–g – 10μm, h – 1 μm.
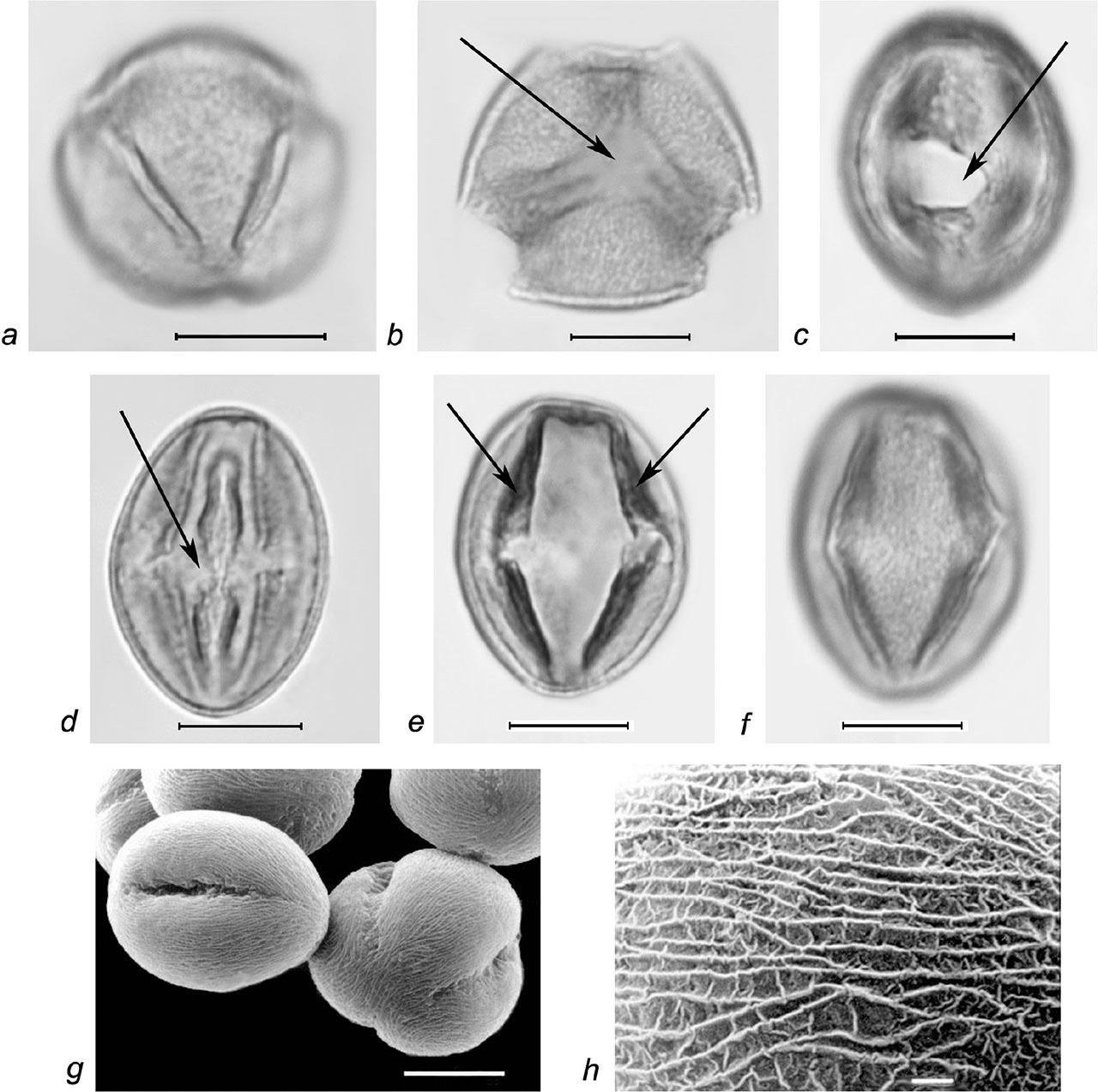
Pollen grains of Rubus canescens DC. a–f – LM micrographs: a – pollen grains in semipolar view, b – synaperturate pollen grain in polar view (synaperture marked with arrow), c–f – pollen grains in equatorial view (c – os, d – genuculum, e – thickened edges of colpi, marked with arrows, f – striate-reticulate exine ornamentation); g–h – SEM micrographs: g – pollen grains in semipolar and equatorial view, h – finely striateperforate exine ornamentation (SEM); scale bars: a–g – 10 μm, h – 1 μm.
Slika 9: Pelodna zrna vrste Rubus canescens DC. a–f – LM mikrografi: a – pelodno zrno v semipolarnem pogledu, b – po daljšnici anastamozna odprtina (označeno s puščico), c–f – pelodno zrno v ekvatorialnem pogledu (c – os, d – genikulum, e – odebeljeni rob brazde, označen s puščico, f – črtasto-mrežasta ornamentacija eksine; g–h SEM mikrograf: g – pelodno zrno v semipolarnem in ekvatorialnem pogledu, h – fino črtasto-luknjasta ornamentacija eksine (SEM); scale bars: a–g – 10 μm, h – 1 μm.
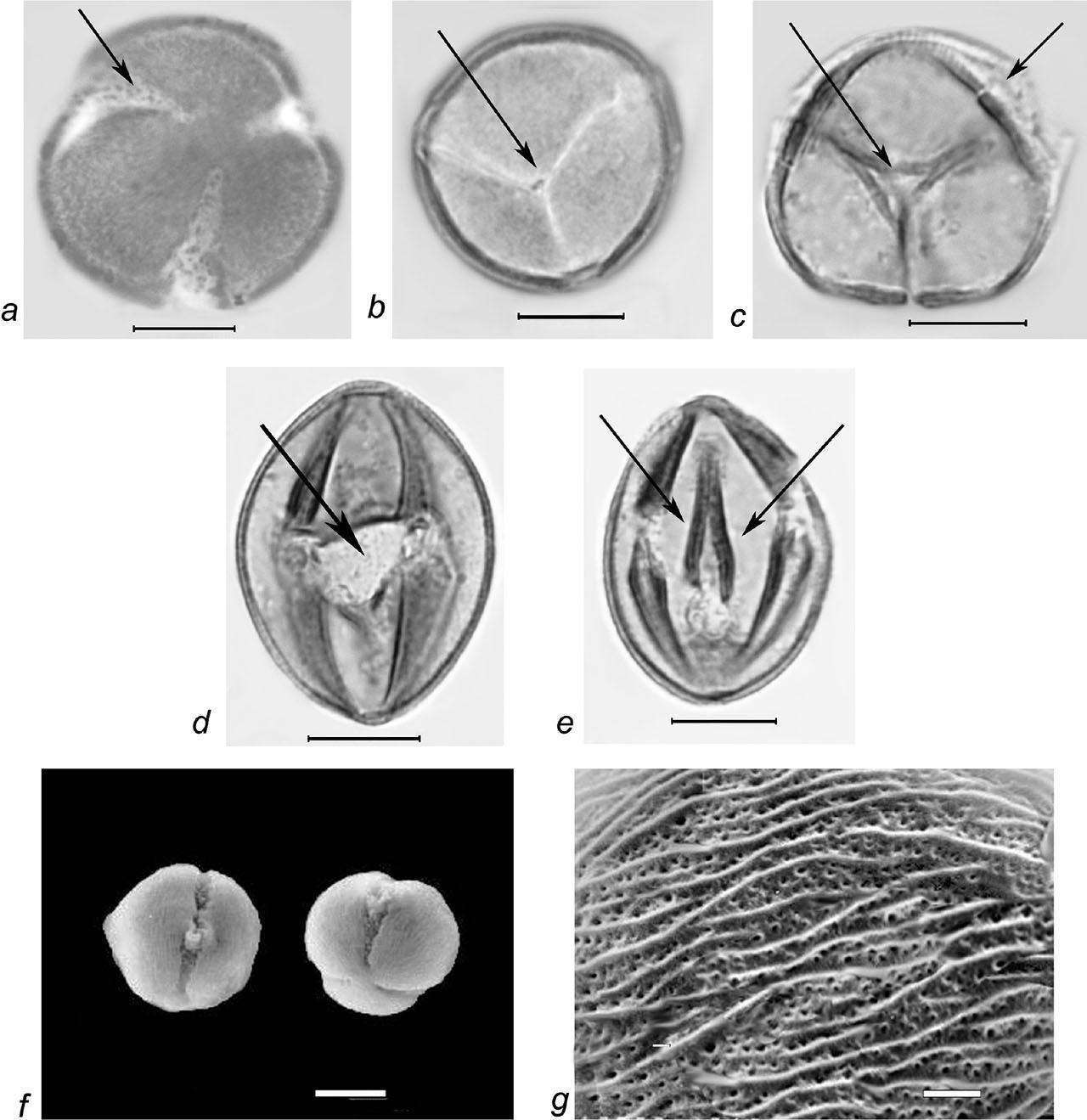
Pollen grains of Rubus cartalinicus Juz. a–e – LM micrographs: a–c – pollen grains in polar view (a – pollen and colpus membrane ornamentation, b–c – synaperturate pollen grains and os area, marked with arrows), d–e – pollen grains in equatorial view (d – os, e – thickened edges of colpi); f–g – SEM micrographs: f – pollen grains in equatorial and semipolar view, g – finely striate-perforate exine ornamentation; scale bars: a–f – 10 μm, g – 1 μm.
Slika 10: Pelodna zrna vrste Rubus cartalinicus Juz. a–e – LM mikrografi: a–c – pelodno zrno v polarnem pogledu (a – pelod in ornamentacija membrane brazde, b–c pelodno zrno z anastamozno odprtino, označeno s puščico), d–e – pelodno zrno v ekvatorialnem pogledu (d – os, e – odebeljen del brazde); f–g – SEM mikrograf: f – pelodno zrno v ekvatorialnem in semipolarnem pogledu, g – fino črtasto-luknjasta ornamentacija eksine; merilce: a–f – 10 μm, g – 1 μm.
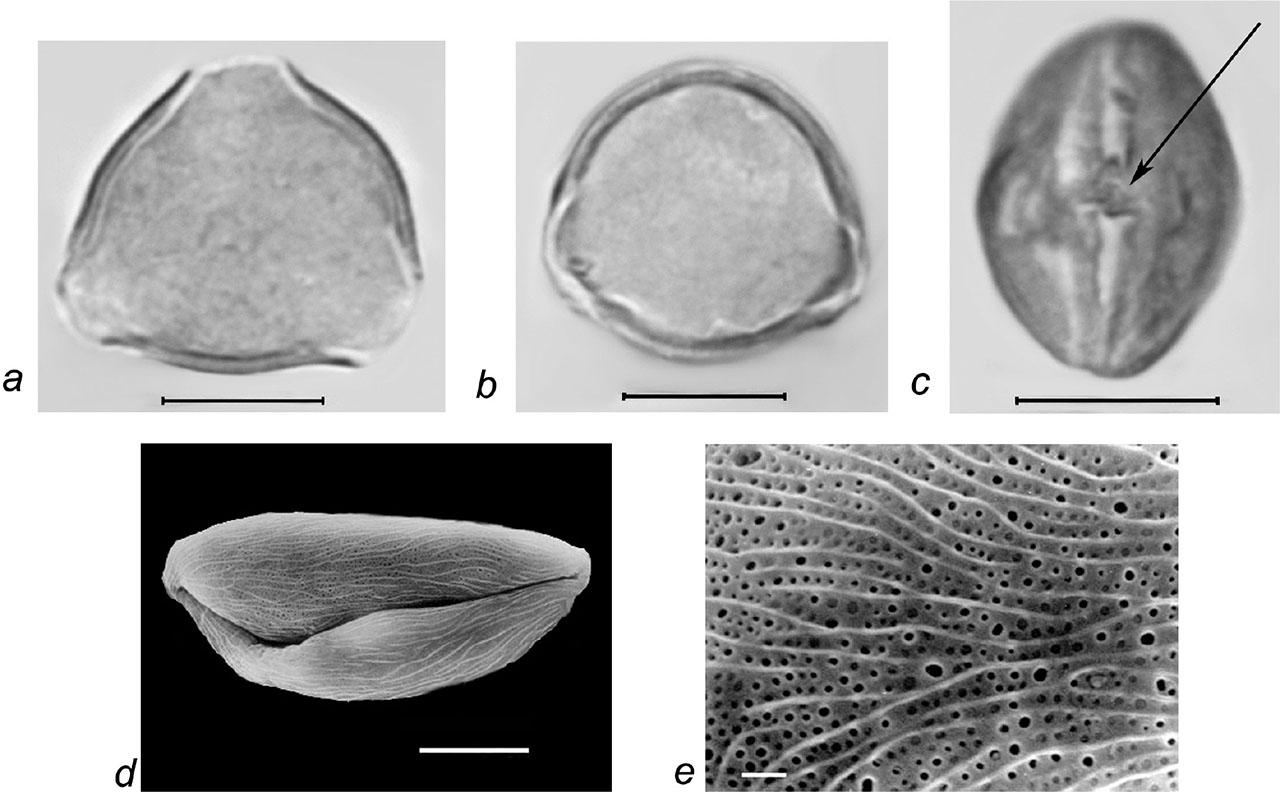
Pollen grains of Rubus sanctus Schreb. a–c – LM micrographs: a – pollen grain in polar view, b–c – pollen grains in equatorial view (c– colpus with genuculum, marked with arrow); d–e – SEM micrographs: d – pollen grain in equatorial view, e – finely striate-perforate exine ornamentation; scale bars: a–d – 10 μm, e – 1 μm.
Slika 11: Pelodna zrna vrste Rubus sanctus Schreb. a–c – LM mikrografi: a – pelodno zrno v polarnem pogledu, b–c – pelodno zrno v ekvatorialnem pogledu (c– brazda z genikulumom, označeno s puščico); d–e – SEM mikrograf: d – pelodno zrno v ekvatorialnem pogledu, e – fino črtastoluknjasta ornamentacija eksine; merilce: a–d – 10 μm, e – 1 μm.
The striae are long, often dichotomously branched; in the species R. canescens, the individual striae are connected by thin transverse bridges (Figure 9h). The diameter of the perforations is 0.1–0.7 μm; the strongest variability of this character was found in the species R. sanctus (Figure 11e).
Our studies revealed the presence of predominantly 3(4)-zonocolporate pollen apertures in representatives of the genus Rubus from the Armenian flora, where endoapertures are formed due to thinning or complete absence of the internal layers of the exine. Additionally, in some 4-zonocolporate pollen grains of the species R. armeniacus, additional slit-like or colpus-like areas were observed at one or both poles (Figure 6b), indicating a possible hybridogenic nature of this species. It should also be noted that such spheroidal or oblate spheroidal endoapertures, often invisible beyond the colpi, have been considered by several authors as pores (e.g., Avetisyan & Manukyan, 1958; Kuprianova, 1978; Kosenko et al., 1984; Tomlik-Wyremblewska, 1995). On the other hand, pollen studies of five West-European Rubus species (Reitsma, 1966), as well 18 species from the Iberian Peninsula (Monasterio-Huelin & Pardo, 1995) revealed lalongate endoapertures formed due to the disappearance or thinning of the nexine, thereby confirming our standpoint. Some authors characterized these areas as endopores with irregular margins and a fastigium (Wronska-Pilarek et al., 2006; Wronska-Pilarek et al., 2012).
A characteristic feature of pollen grains of the genus Rubus is the presence of a geniculum, i.e., a bulge in the equatorial exine of the colpus, usually formed due to the separation of the sexine from the nexine and the rupture of the latter (Potonié, 1934; Punt et al., 2007). The presence of a similar bulge in some species of the genus Rubus, referred to as “fastigium”, was noted by Reitsma for the first time (1966). It is possible that the presence of the geniculum covering the ora led several authors to characterize Rubus pollen as tricolpate (e.g., Demchenko, 1967; Eide, 1981b). Some authors noted the presence of an equatorial bridge, with margins constricted at the equator (Tomlik-Wyremblewska, 1995).
The general outline of pollen grains in the investigated species was mainly prolate (P/E = 8/6–8/4) or subprolate (P/E = 8/7–8/6), prolate spheroidal (P/E = 8/8–8/7) in R. cartalinicus, and subspheroidal (6/8–8/6) in R. takhtadjanii (Table 2, Figure 1).
In most cases, pollen grains were small. However, for R. caesius, R. cartalinicus Juz., R. caucasicus Focke, R. sanctus both small and medium-sized pollen were noted, with Pmax= 33.5 μm observed in R. sanctus (Table 2). The smallest pollen grains (according to polar axis length) were noted in the rare endemic species R. takhtadjanii (on average 14.2 μm), and the largest ones in R. caucasicus (on average 26.3 μm). According to available literature data, the largest pollen grains in the genus Rubus (P × E = 42.5 × 35.8 μm) were found in the species R. alceifolius Poir. (Kosenko et al., 1984).
In general, according to Monasterio-Huelin & Pardo (1995, p. 231), “the comparative analysis of the mean values of P and E is not significant in distinguishing pollen types”.
The thickness of the exine layer in the species studied ranged from 1.2 to 1.3 μm, resulting in a certain percentage of destroyed pollen grains during acetolysis treatment According to literary sources, the maximum exine thickness was noted in the species R. nutkanus Moc. (2.5 μm) (Demchenko, 1967).
Examination of the surface of pollen grains using SEM revealed finely striate-perforate exine ornamentation, sometimes (in R. armeniacus, R. caesius) combined with finely striate-microreticulate one (Figure 6j, Figure 7h). A similar type of sculpture is also noted for the pollen of approximately twenty species of the genus Rubus in the Iberian Peninsula (Monasterio-Huelin & Pardo, 1995). The exception is R. genevierii Boreau, “… whose ornamentation is striate-gemmate-perforate and with a striated margin” (Monasterio-Huelin & Pardo, 1995, p. 229). Pollen studies of Rubus representatives have also revealed some other different types of ornamentation besides the perforate-stiate one, including rugulate, rugulate-striate, striate-scabrate, stiate-gemmate, stiate-reticulate, rugulate, reticulate, clavate-bacculate (Gonzalez Romano & Candau, 1989; Ueda, 1992; Tomlik-Wyremblewska, 1995, 2000, 2004; Zhou et al., 1999; etc.).
A wide-ranging research of the pollen morphology of 155 species and 13 varieties representing all 12 subgenera of Rubus revealed six types of exine ornamentation (I – perforate, II – microreticulate, III – striate-perforate, IV – rugulate-perforate, V – echinate, VI – striate-micro-reticulate) and three subtypes (IA– scabrate-perforate, IB – perforations and discontinuous muri; IIIA – scabrate-striate-perforate) (Xiong et al., 2019).
It is possible that the pronounced polyploidy has played an undoubted role in the display of pollen polymorphism within the genus Rubus. For example, in contrast to the striate or reticulate sculpture, sometimes a vermiculate sculpture is characteristic of the pollen of many species (Ghosh & Saha, 2017; Valdés et al., 1987; Wronska-Pilarek et al., 2012); the systematically and ecologically isolated dioecious octoploid glacial relict R. chamaemorus L. (2n = 8× = 56) (Thiem, 2003) exhibits spinose (Erdtman, 1952; Demchenko, 1967; Faegri & Iversen, 1989), verrucate, gemmate (Reitsma, 1966; Hebda & Chinnappa, 1990, 1994) or perforate-striate (Eide, 1981a) pollen exine ornamentations.
Disagreements also arise when characterizing the type of apertures in the aforementioned species, namely, 3-colpor(oid)ate (Erdtman, 1952) vs tricolpate (Demchenko, 1967). The latter author also noted clear differences in pollen characteristics between R. chamaemorus and two closely related species, R. trifidus Thunb. and R. nutkanus (synonym of R. parviflorus Nutt.), both in terms of aperture type (tricolpate vs tricolporate) and exine ornamentation type (spinose vs reticulate-foveolate and sinuously reticulate, respectively). Additionally, tricolpate pollen grains (Eide, 1981b) and tuberculate sculpture for the species R. arcticus L. (Surova, 1975), as well as micro-verrucate exine ornamentation for R. parviflorus (Hebda & Chinnappa, 1990) were reported. In both R. idaeus (Eide, 1981b) and R. sanctus (our studies), a significant diversity in the total size of pollen grains within the same sample (almost twice) was observed. A considerable variability in pollen grain sizes between the different samples of species R. caesius and R. idaeus is shown in Figure 1. On the other hand, a study of pollen morphology and pollen variability in R. gracilis from 13 natural Polish localities “… revealed no differences among the grains from the individual localities, likely the result of apomixis” (Wronska-Pilarek et al., 2006, p. 69).
Pollen morphology of 10 Armenian species of the genus Rubus was studied using both light microscopy (LM) and scanning electron microscopy (SEM), including the species R. candicans, R. cartalinicus and R. takhtadjanii investigated for the first time.
Analysis of the data obtained revealed significant palynomorphological uniformity both in the type of apertures (mainly 3(4)-zonocolporate) and the type of exine ornamentation (finely striate-perforate), in combination with finely striate-microreticulate one in R. armeniacus and R. caesius. On the other hand, the obtained data confirmed the observations of Wronska-Pilarek et al. (2012) and Xiong at al. (2019) regarding the significant variability of certain pollen features within individual species of the genus Rubus (such as pollen shape, size, etc.), and especially within the subgenus Rubus (= Eubatus), to which most of the Armenian species of the genus Rubus belong. For example, in R. sanctus a significant variability in the length of the polar axis (P) was revealed, ranging from 15.5 to 33.5 μm.
In this context, our data support the previously held view that pollen grain morphology within the genus Rubus can only be used as an auxiliary feature for the diagnosis of individual species.