
Glycosylation is one of the most common and complex post-translational protein modifications. In eukaryotes, glycan biosynthesis takes place in the ER-Golgi pathway. Glycosylation is an enzymatic process involving glycosyltransferases and glycosidases. These enzymes are responsible for the assembly, processing, and turnover of glycans. The biological function of glycans ranges from relatively subtle to being crucial for the development, growth, maintenance, or survival of an organism. Glycans can be divided into three general categories: structural and modulatory properties; specific recognition by other molecules; and molecular mimicry of host glycans. The appropriate glycan composition and structure are essential for cellular biological processes (Varki et al. 2022).
There are several documented mechanisms of protein glycosylation, e.g., N-glycosylation and O-glycosylation. At the level of the individual protein, glycosylation can have a profound effect on folding, conformation, stability, solubility, function, activity, subcellular localisation, protection from proteases, and cell-cell interactions. Interestingly, the same glycan can fulfill distinct functions depending on the tissue type, developmental stage, or environmental conditions. Therefore, any change in glycan profile, turnover, degradation, or recognition may lead to the development of disease. Altered glycosylation contributes to the development of neurodegenerative, autoimmune, and metabolic diseases, as well as cancers, by impairing immune regulation, protein function, and cellular signaling.
Additionally, glycome plays a key role in bacterial inflammation by modulating host-pathogen interactions, immune recognition, and epithelial responses. Microbial factors in the vaginal environment, including bacteria, yeast, and viruses, can influence glycan profiles and contribute to changes in glycan profiles associated with infection. Knowledge of the glycosylation profile related to a given disease can improve diagnosis and therapies for that disease (Eichler et al. 2019; Paton et al. 2021; Varki et al. 2022; Cheng et al. 2025; Lei et al. 2025).
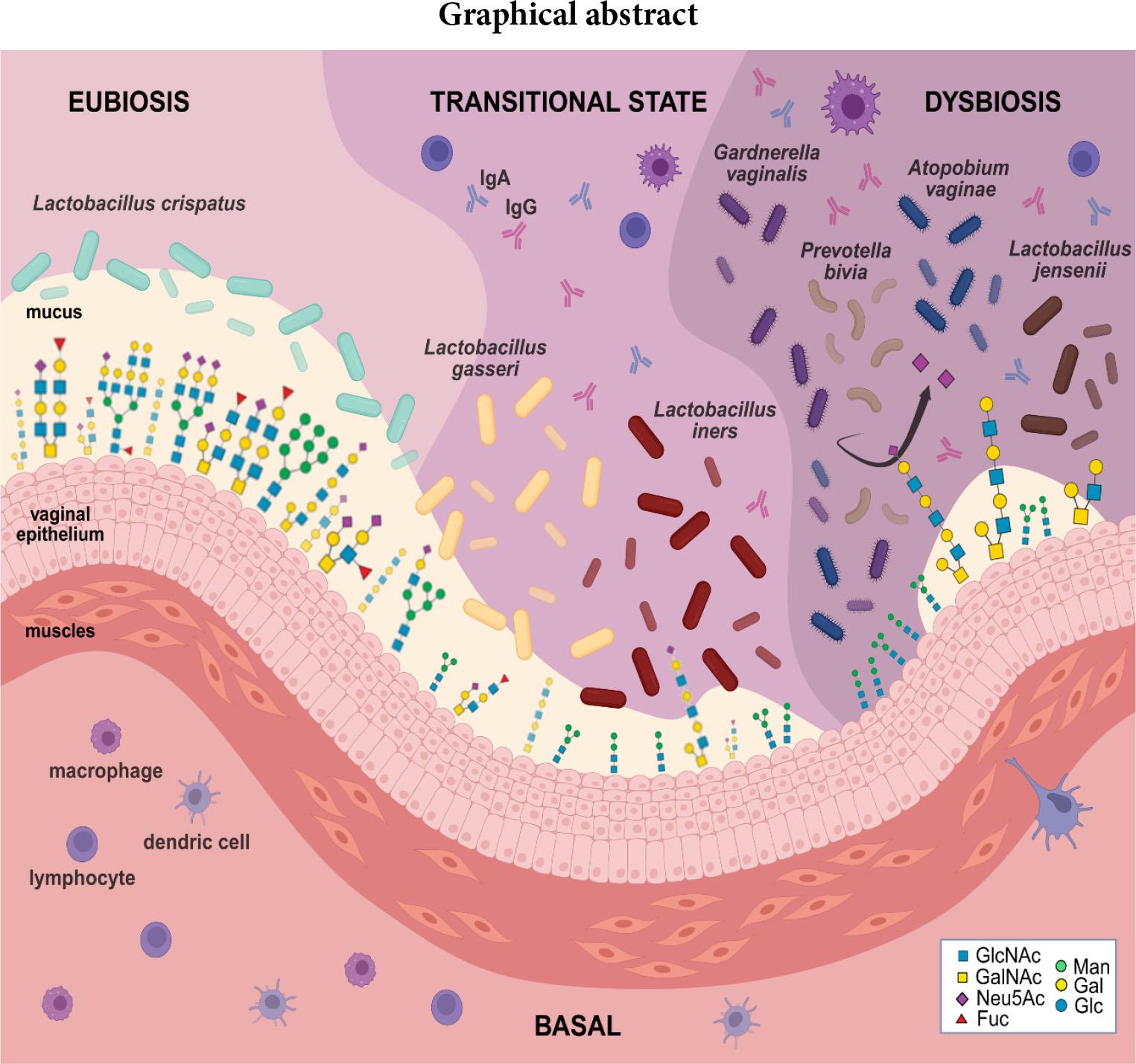
Vaginal microbiota: shift from eubiosis to dysbiosis. Legend: Sugar residues forming the structures of N- and O-glycans: GlcNAc – N-acetylglucosamine; GalNAc – N-acetylgalactosamine; Neu5Ac – N-acetylneuraminic acid; Fuc – fucose; Man – mannose; Gal – galactose; Glc – glucose.
Cervicovaginal mucus (CVM) is a viscoelastic product formed by the epithelial cells of the uterus, cervix, and vagina. CVM provides vaginal lubrication and acts as a natural barrier against microorganisms, protecting against physical and chemical damage and infection. CVM consists predominantly of water (> 95% by weight), gel-forming mucins (2–5%), and a complex mixture of immunoglobulins (secretory IgA (sIgA) and IgG), plasma proteins, antimicrobial peptides, sterols, lipids, carbohydrates, cytokines, lactoferrin, and inorganic ions (about 1%). Mucins are high-molecular-weight glycoproteins predominantly modified via O-linked glycosylation, with a minor contribution from N-linked glycans (Dong et al. 2023). The density, clustering, and variety of linear or branched O-GalNAc glycans on mucins determine their chemical, physical and biological properties. Cervicovaginal fluid (CVF) is a heterogeneous mixture of antimicrobial peptides and glycoproteins, including sIgA and IgG antibodies, cathepsin G, lysozyme, and lactoferrin, as well as cellular components such as epithelial cells and leukocytes (Fig. 1) (Lacroix et al. 2020).
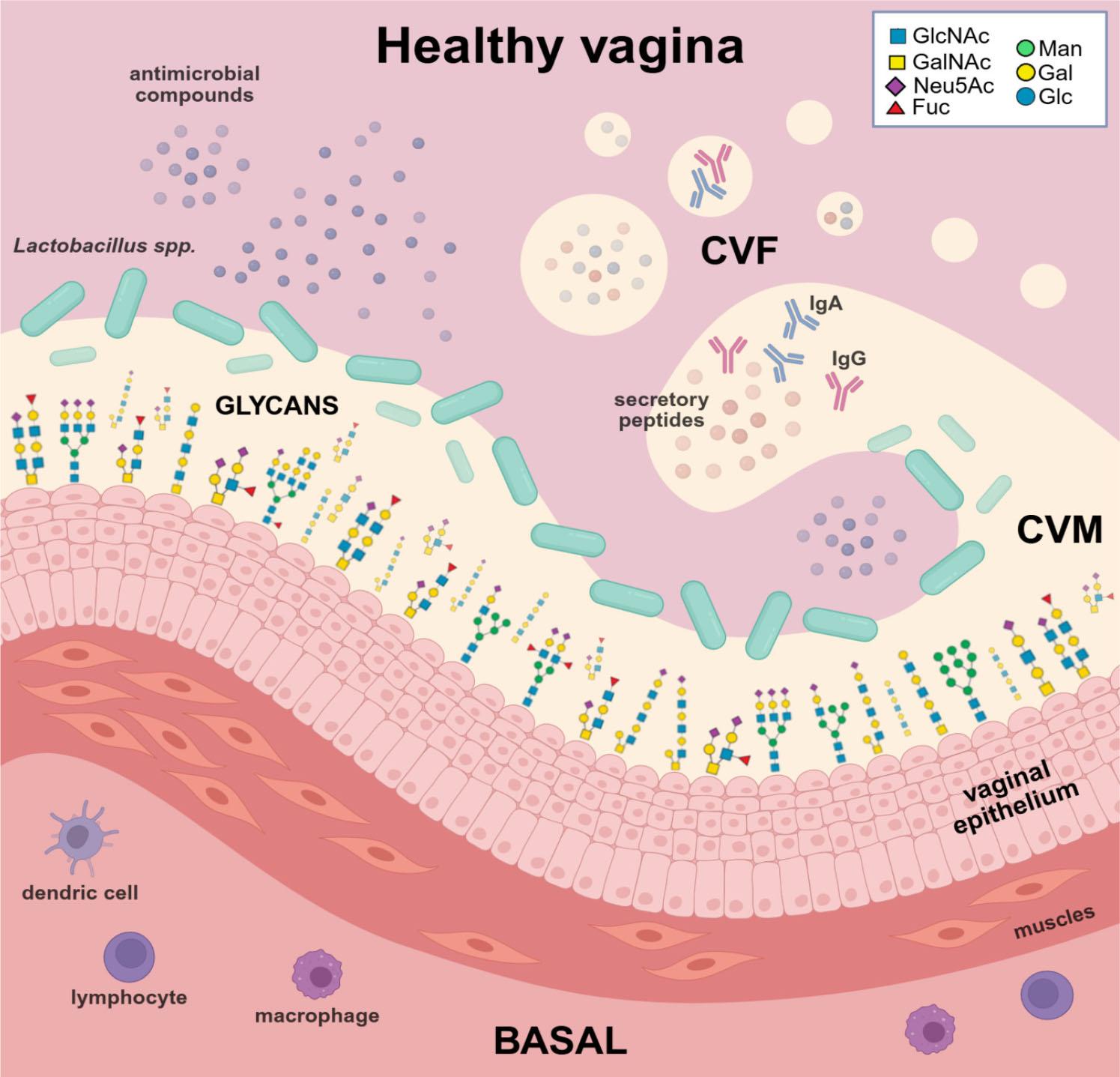
The healthy vaginal epithelium coated by a protective mucus layer (CVM) and naturally secreted fluid (CVF), maintaining hydration and microbial balance. The figure was created in BioRender.com (https://BioRender.com/npsbi58) (Bodaszewska-Lubas). Legend: Sugar residues forming the structures of N- and O-glycans: GlcNAc – N-acetylglucosamine; GalNAc – N-acetylgalactosamine; Neu5Ac – N-acetylneuraminic acid; Fuc – fucose; Man – mannose; Gal – galactose; Glc – glucose.
Glycans present in cervicovaginal fluid (CVF) originate from diverse cellular and secretory sources, including exfoliated epithelial cells from the cervical and vaginal mucosa, secreted mucins produced by these epithelial cells, immunoglobulins such as sIgA and IgG, and infiltrating immune cells (Wu et al. 2022; 2024). Mucins in mucus secretions, including endocervical and vaginal secretions, are either large, polymeric (gel-forming) or smaller, non-gel-forming, and monomeric. O-glycans are hydrophilic, negatively charged, and promote the binding of water and salts, making them major contributors to the viscosity and adhesiveness of mucus. The female genital tract contains approximately 20 gel-forming mucins, the major ones being MUC2, MUC5AC, MUC5B, and MUC6 (Lacroix et al. 2020; Chen et al. 2024). In summary, vaginal epithelial cells are covered by a glycan layer consisting of diverse glycans that function as molecules for cellular recognition, adherence, and signaling, and as mediators of host-microbe interactions (Wu et al. 2022; Agarwal et al. 2023).
Pathogens involved in vaginal dysbiosis express a variety of virulence factors, e.g., adherence factors (pili or fimbriae), toxins (exotoxins and endotoxins, e.g., lipopolysaccharide of Gram-negative bacteria, LPS), and enzymes (IgA proteases, tissue-degrading enzymes or glycan-degrading enzymes) (Brooks et al. 2013; Yamini and Nestorovich 2017; Ingelfinger 2017; Mostowy and Holt 2018; Chen et al. 2024). Sialidases (neuraminidases) play a significant role in pathogenicity by cleaving terminal sialic acid residues from glycoconjugates of the CVM and CVF, thereby damaging their physical and immunological properties. Additionally, bacterial sialidases hydrolyze sialic acid residues from glycoconjugates, epithelial receptors, immunoglobulins (especially sIgA), and cytokines, thereby inhibiting their immune reactivity (Lewis et al. 2012; Bonnardel et al. 2021; Chen et al. 2024). Alterations in the glycan profile may result from the activity of pathogen-derived glycosidic enzymes, which disrupt vaginal mucus and the epithelium (Pérez-Ibave et al. 2025).
In healthy women, the vaginal epithelium is predominantly colonised by lactic acid bacteria (LAB). The most common species in the vagina are Lactobacillus crispatus, followed by Lactobacillus gasseri, Lacticaseibacillus rhamnosus, and Lactobacillus jensenii, Lactiplantibacillus plantarum and Lactobacillus iners. LAB defend against harmful bacteria by producing immunomodulatory and antimicrobial compounds such as lactic acid, hydrogen peroxide, various bacteriocins, and an arginine deaminase enzyme (Bodaszewska-Lubas et al. 2012; Dobrut et al. 2018; Segui-Perez et al. 2024; Pérez-Ibave et al. 2025). Interestingly, significant overlap of LAB strains is observed in rectal and vaginal samples from the same woman (Krog et al. 2022). Five Community State Types (CSTs) were distinguished based on vaginal microbiota status (Lee et al. 2020) and of the four vaginal CSTs – I, II, III and V – LAB species predominate, with each CST characterized by a distinct dominant species: L. crispatus (CST I), L. gasseri (CST II), L. iners (CST III), and L. jensenii (CST V). CST IV is a diverse group characterized by higher proportions of strictly anaerobic bacteria, including Prevotella, Dialister, Atopobium, Gardnerella, Megasphaera, Peptoniphilus, Sneathia, Eggerthella, Aerococcus, Finegoldia, and Mobiluncus (Ma and Li 2017). Depending on the vaginal pathogen, vaginitis can be categorized into several types. The most common types are aerobic vaginitis (AV), bacterial vaginosis (BV), trichomoniasis vaginitis (TV), and vulvovaginal candidiasis (VVC), as well as mixed vaginitis, which is caused by two or more coexisting microorganisms. Biofilm formation may be a crucial infection mechanism in mixed vaginitis or bacterial vaginosis (Shang et al. 2024). A shift in vaginal microbiota from eubiosis to dysbiosis is connected with the colonization of the vaginal epithelium by pathogens. Microorganisms express a multitude of virulence factors and antigens, causing progressive mucus and endothelial damage and inhibiting or skewing immune system activity, for example, via antigen mimicry. In most cases, different types of pathogens and host glycans or mechanisms are involved, such as changes to, degradation of, or use of glycan structures or single sugar residues. Analyzing and understanding changes in glycan alterations will allow us to conclude putative glycan biomarkers of genital tract disturbances or new treatment targets (Pérez-Ibave et. al. 2025).
N-glycan biosynthesis begins with a precursor structure from which high-mannose, hybrid, and complex glycans are synthesized. The antennae of complex and hybrid N-glycans often terminate with specific glycan epitopes such as Lewis antigens, ABO blood group antigens, and poly-N-acetyllactosamine (polyLacNAc) chains. These important structures are recognized by lectins (glycan-binding proteins, GBPs) (Varki et al. 2022). The presence of rich, extensive N-glycans has been demonstrated in CVF collected from pregnant and non-pregnant women using a detailed MALDI-MS method. Analysis showed the presence of low-mass-range paucimannose N-glycans, high-mannose and biantennary complex N-glycans with fucosylation and/or sialylation (mid-mass-range N-glycans), and glycans in the higher-mass range containing polyLacNAc units decorated with a different number of fucose and sialic acid residues. The CVF N-glycan profile depends on many factors, including the microbial composition of the vagina, which is classified into CSTs (Wu et al. 2022; 2024). The CST I, CST III, and CST IV statuses of examined vaginal samples were compared. In the CST I-A state, characterized by the abundance of L. crispatus, the highest levels of sialylated and bisected glycans were detected, but lower levels of high-mannose glycans. Higher levels of paucimannose glycans were observed in CST III-A microbiota communities, which are dominated by L. iners. In CST IV-B samples, which are characterized by the dominance of Gardnerella vaginalis and Atopobium vaginae, higher levels of paucimannose glycans and non- or mono-fucosylated N-glycans with two or three LacNAc units were detected, as well as lower levels of bi-, tri- or tet-ra-fucosylated N-glycans. These results suggest that a higher occurrence of LAB is associated with a higher percentage of sialylated glycans, as well as bi-, tri-, and tetra-fucosylated glycans, and lower levels of non- and mono-fucosylated glycans (Wu et al. 2022). Changes in the vaginal N-glycome profile in CST IV compared to CST I may result from the different mechanisms of action of harmful bacteria producing enzymes. Sialic acid-decorated glycans are a prominent feature of cervicovaginal mucus (CVM) and cervicovaginal fluid (CVF), where they contribute to the viscoelastic properties of mucus and play key roles in immune defense, microbial recognition, and reproductive health (Moncla et al. 2016). The negatively charged mucins, which are conferred by sialic acid residues, maintain a rigid structure and trap pathogens, thereby preventing them from reaching the vaginal epithelium. Desialylation of glycans in the vagina during bacterial infection by sialidases alters their structure, charge, and function, exposing them to various hydrolytic enzymes and rendering them susceptible to proteolysis, thereby hampering the immune response against bacteria (Lewis et al. 2012; Bonnardel et al. 2021; Chen et al. 2024).
The changes in N-glycan profiles observed in the vaginal microbiota result from the activity of various hydrolytic enzymes, such as sialidases, sulfatases, and glycosidases, as well as proteolytic enzymes that degrade host glycans. Conversely, the sugar composition of N-glycans determines the bacterial vaginal community. Pathogens modify N-glycan decoration and composition for their own use, facilitating processes such as adhesion, colonization, invasion, and evasion (Pérez-Ibave et al. 2025).
The mechanism of adhesion of harmful microbes is the first step in the infection process. S-layers are crystalline, two-dimensional (2D) lattices that self-assemble and enclose microbial cell envelopes. They are created by glycoprotein and protein monolayers. S-layer proteins (SLPs) can be single proteins or glycosylated proteins with covalently bound carbohydrate chains. Certain S-layer functions may be supplemented by a novel subgroup of proteins known as S-layer-associated proteins (SLAPs). S-layers have been identified in various bacterial species, including the genus Lactobacillus, and are often linked to the probiotic properties of lactobacilli strains, such as enhanced adherence to host epithelial cells and modulation of the immune response. SLPs, through interaction with immune receptors, promote anti-inflammatory signaling and enhance mucosal barrier integrity. They can stimulate the production of protective cytokines (e.g., IL-10) while suppressing pro-inflammatory mediators, helping to prevent overactivation of the immune system (Klotz et al. 2020a; 2020b; Adapen et al. 2022). When assessing the adherence of commensal LAB strains and BV-involved strains to the vaginal cell line VK2/E6E7 and the endocervical cell line End1/E6E7, the strongest adherence was demonstrated by L. crispatus, followed by intermediate adherence by G. vaginalis and Prevotella timonensis, and the weakest by Prevotella bivia (Segui-Perez et al. 2024).
SLPs (surface-located proteins) isolated from different vaginal LAB species, e.g., SLPs isolated from Lactobacillus johnsonii or SLPs purified from L. crispatus strains, possess the ability to interact with the receptor DC-SIGN (C-type lectin receptor, CLR). CLRs generally recognize pathogen-associated molecular patterns (PAMPs) to promote innate immune responses and affect adaptive immune responses. Surprisingly, bacterial SLPs were detected in human vaginal secretions, either released freely into the environment or incorporated into extracellular vesicles. SLPs could be detected in some cervical vaginal fluid (CVF) samples from pregnant women with CST I and CST III, as well as in LAB-depleted samples with high-diversity compositions enriched for bacteria associated with bacterial vaginosis (BV). Among L. crispatus-dominant samples, the detection of SLPs in the CVF was associated with lower levels of IL-1β and IL-8. Malamud et al. (2020) analyzed the glycosylation profiles of three SLPs: SLP-8348, SLP-8321, and SLP-5818, expressed by the probiotic strain Lactobacillus kefiri CIDCA 8348 (SLP-8348). The significant component in the O-glycans of the three analyzed SLPs was glucose; however, differences in the length of the hexose chains were observed. N-glycosylation was detected only in SLP-5818, which had two glycosylation sites (Malamud et al. 2020). Conversely, SLP detection was associated with increased IL-6 levels in samples dominated by L. iners. Transitional L. iners provides reduced protection against vaginal dysbiosis (Decout et al. 2024). Bacterial vaginal infections significantly alter the local immune response by disrupting the LAB-dominated microbiota. Dysbiosis leads to the activation of pattern recognition receptors (PRRs), including Toll-like receptors (TLRs) and nucleotide-binding oligomerization domain-like receptors (NLRs). Elevated microbial diversity triggering the release of proinflammatory cytokines such as IL-1β, IL-6, and TNF-α. These cytokines recruit neutrophils and macrophages, contributing to mucosal inflammation and epithelial barrier disruption. Beyond Gardnerella, other bacterial species implicated in vaginal infections – such as A. vaginae, Mobiluncus spp., and Prevotella spp. – elicit distinct immune responses, often characterized by elevated levels of matrix metalloproteinases (MMPs), reactive oxygen species (ROS), and antimicrobial peptides, which further compromise epithelial integrity and promote chronic inflammation. In summary, SLPs of LAB support vaginal mucosal homeostasis by shielding proinflammatory ligands and promoting anti-inflammatory signaling, whereas L. iners and G. vaginalis may engage both pro- and anti-inflammatory pathways, potentially leading to suboptimal immune responses and contributing to dysbiosis (Adapen et al. 2022; Dong et al. 2023).
Streptococcus agalactiae (group B streptococcus, GBS) is present in the vagina or rectum of around 30–40% of pregnant women, and epidemiological data confirm the association between GBS colonization and infection in term and preterm infants. Furthermore, GBS has been identified as a commensal bacterium in adults and non-pregnant women (Bodaszewska-Lubas et al. 2012; Maidment et al. 2024). The mechanism by which GBS strains adhere to HT-29 colorectal adenocarcinoma cells and A-431 human epidermoid vulvovaginal cells may depend on alpha-like protein genes and GBS serotypes, respectively (Bodaszewska-Lubas et al. 2013). Other GBS adhesion surface antigens identified, such as the serine-rich repeat protein (Srr) and pili, were highly glycosylated. A mouse model of GBS vaginal colonization showed that wild-type (WT) GBS strains exhibited a significant survival advantage over ΔpilA or Δsrr-1 mutants in the vaginal tract (Sheen et al. 2011). Pellegrini et al. (2024) characterized the molecular mechanism by which the Srr2 of GBS interacts with fibronectin, a multi-domain glycoprotein found in body fluids, the extracellular matrix, and on cell surfaces. Fibronectin was identified as a novel ligand for the Srr2 adhesin of GBS, promoting the adhesion of GBS to human cervicovaginal epithelial cells. In the presence of fibronectin, the ability of a Δsrr-2 mutant to adhere to human cervicovaginal epithelial cells was significantly lower than that of the wild-type strain (Cinar et al. 2023; Pellegrini et al. 2024).
Researchers have identified and characterized the Neisseria gonorrhoeae host-glycan binding profile (glycointeractome) as another mechanism of bacterial infection. The results revealed numerous host-pathogen interactions, including high-affinity binding to mannosyl glycans. The most abundant mannose-binding gonococcal proteins identified were three opacity proteins (NGO1073a, NGO1040a, and NGO1513) (Semchenko et al. 2019). The researchers then analyzed the role of the neisserial heparin-binding antigen (NHBA) in epithelial cell adherence. They confirmed the role of NHBA as a binding structure with the highest calculated affinity for heparin, preferentially binding to glycans with a specific sulphation configuration, such as chondroitin sulphate C or heparan sulphate. A gonococcal NHBA mutant displays reduced survival in human serum and reduced adherence to human cervical and urethral epithelial cells relative to the wild-type strain. These results suggest that the gonococcal NHBA protein plays a role in bacterial colonization and survival (Semchenko et al. 2020).
Changes in the N-glycan profile of the vagina depend on the microbiota community and result from the activity of various bacterial mechanisms, e.g., hydrolytic enzymes such as sialidases (neuraminidases), sulfatases, and glycosidases, as well as proteolytic enzymes that degrade host glycans. Consistently, the sugar composition of glycans influences the vaginal bacterial community. Pathogens modify glycan composition and decoration for their own use, facilitating processes such as adhesion, colonization, invasion, and evasion. Pathogen sialidases cause lower sialylation of CVM and CVF glycans in the vagina during infection. The function of glycan-decorated sialic acid is to determine the negative charge, tertiary structure, and function of glycans. Negatively charged mucins maintain a rigid structure and trap pathogens, preventing them from reaching the vaginal epithelium. Bacterial cleavage of terminal sialic acid residues from glycoconjugates on the CVF or the vaginal epithelium surface alters their physical and immunological properties, thereby abolishing their functions. Additionally, bacterial sialidases cleave sugar residues from epithelial receptors, immunoglobulins, and cytokines. The remaining uncovered carbohydrate residues are susceptible to various pathogen glycosidases and are more susceptible to proteolytic degradation, which hampers the immune response against bacteria. In summary, sialic acid residues protect the oligosaccharide and protein structure of mucin molecules and N-glycans in the vagina from enzymatic degradation. Additionally, bacteria use hydrolyzed sialic acid from glycoproteins as a carbon source for nutrition and colonization (Lewis et al. 2012; Bonnardel et al. 2021; Chen et al. 2024).
The overgrowth of anaerobic bacteria associated with bacterial vaginosis (BV) is linked to higher levels of bacterial enzymes such as glycosidases, proteases, prolidases, and sulphatases, which play a role in the development of BV. Bacterial vaginosis is a complex polymicrobial disorder characterized by a depletion of LAB and an overgrowth of mixed anaerobic and facultative bacteria, including G. vaginalis, Prevotella spp., A. vaginae, Bacteroides spp., Mobiluncus spp., Gram-positive cocci, and genital mycoplasmas. BV frequently accompanies other vaginal infections, especially those caused by Trichomonas vaginalis, Chlamydia trachomatis, and N. gonorrhoeae. BV is associated with an increased susceptibility to sexually transmitted infections (STIs), including HIV (human immunodeficiency virus) and HPV (human papillomavirus). G. vaginalis is the best characterized BV-associated bacterium. It requires the presence of other bacteria during vaginal infection; for example, co-existing P. bivia produces ammonia, which stimulates the growth of G. vaginalis and expresses sialidases that digest the vaginal mucus layer, facilitating bacterial adhesion. P. bivia and P. timonensis exhibit sialidase activity that targets the vaginal mucus layer, leading to increased adhesion of other BV-associated bacteria. Additionally, a slightly acidic vaginal pH is optimal for bacterial sialidase activity. Another bacterium often found alongside P. bivia in BV samples is P. timonensis (Segui-Perez et al. 2024). Interestingly, sialidase expression is a typical feature of every strain of P. bivia, but only some G. vaginalis strains. In turn, G. vaginalis strains exhibit lower heterogeneity in sialidase expression than P. bivia at higher concentrations. Based on quantitative polymerase chain reaction (qPCR) targeting clade-specific genes, Gardnerella is divided into four clades. These clades differ in sialidase activity: clade 2 has the highest activity, followed by clades 1, 3, and 4 (Qin and Xiao 2022).
Gilbert et al. (2019) demonstrated that mice infected with G. vaginalis exhibited approximately 4 times higher levels of vaginal sialidase activity. In contrast, those infected with P. bivia exhibited approximately two times the activity observed in mock-infected mice (Gilbert et al. 2019). Sialidases are considered a virulence factor in BV, clipping the negatively charged sialic acid residues from the terminal end of mucin oligosaccharides. BV may cause the mucin gel layer in the vagina to thin and the CVF barrier to lose its protective capacity against viral infections such as HIV or HPV (Wessman et al. 2017). Bacterial genome sequencing detected predicted carbohydrate-active enzyme genes in pathogenic bacterial strains (G. vaginalis, P. timonensis, and P. bivia). P. timonensis possessed many predicted α/β-galactosidases, α/β-N-acetylgalactosaminidases, and α/β-N-acetylglucosaminidases. Both P. timonensis and P. bivia strains possess highly active sialidases and fucosidases. Sialidase plays a central role in immunosuppression, relapse, and recurrence in BV, although this is supported by other hydrolytic enzymes, vaginolysins, and immunomodulatory metabolites (Amabebe and Anumba 2022).
The results of the enzymatic degradation of mucin O-glycans on the surface of the vaginal cell line VK2/E6E7 showed different results depending on the bacterial strain (G. vaginalis, P. timonensis, P. bivia and L. crispatus). After the vaginal cells were treated with bacteria, staining with lectins against fucoses (UEA-1), α-2,3-linked sialic acids (MAL-II) and α-2,6-linked sialic acids (SNA) was used to visualize the different mucin glycan structures. This demonstrated that, in controls, all glycan structures were present on vaginal epithelial surfaces, but these changed after bacterial treatment. Of the analyzed strains, P. timonensis was found to efficiently remove sialic acid and fucose residues from the vaginal epithelium. G. vaginalis significantly reduced UEA-1 and, to a lesser extent, MAL-II staining compared to untreated vaginal epithelium (Segui-Perez et al. 2024). These data suggest that bacteria producing sialidases, resulting from stepwise exodeglycosylation and enhanced proteolysis, influence the function of the antimicrobial mucosa. Due to the elevated activity of sialidases in cervical vaginal fluid (CVF) during bacterial vaginosis (BV), these enzymes are a promising biomarker of BV and the basis of analysis for many BV diagnostic tests. They are also potential targets for BV treatment (Chen et al. 2024). Additionally, inhibiting sialidase activity in whole G. vaginalis cells may be a goal of BV treatment. In a preliminary study, the authors indicated that Zanamivir, an anti-influenza drug, inhibited G. vaginalis sialidases and influenced host–pathogen interactions in vitro. It is worth remembering that other BVABs (bacterial vaginosis-associated bacteria), viruses, and even the human body itself can produce sialidases (Govinden et al. 2018; Qin and Xiao 2022).
PAMPs are specific, highly conserved molecular structures, such as lipids, proteins, and nucleic acids, that are characteristic of certain types of pathogens. PAMPs are recognized by innate immune cells via pattern recognition receptors (PRRs), most of which are TLRs, NLRs, retinoic acid-inducible gene I-like receptors (RLRs), and CLRs. CLRs are predominantly expressed on dendritic cells, macrophages, monocytes, neutrophils, and natural killer (NK) cells, and they exhibit organ-specific expression patterns across tissues such as the lung, liver, heart, kidney, and brain, where they contribute to immune surveillance, homeostasis, and inflammation. CLRs recognize carbohydrates through a carbohydrate recognition domain (CRD), known as a C-type lectin-like domain (CTLD), in a calcium-dependent manner, and mediate their internalization into cytoplasmic vesicles for subsequent degradation and elimination (Lech et al. 2012; Kerscher et al. 2013; Scur et al. 2023). Depending on their location within the cell, CLRs are categorized as either transmembrane or secretory receptors. The primary representative of the latter is the collagen lectin family. This family mainly includes surfactant proteins (SPs) and mannose-binding lectins (MBLs). MBL recognizes terminal microbial glycans such as mannose, fucose, and N-acetylglucosamine, enabling binding to structures like LPS of Gram-negative bacteria, fungal mannans, and viral envelope glycoproteins, without affecting the glycosylation process itself. This interaction activates the lectin complement pathway, contributing to pathogen clearance (Man-Kupisinska et al. 2018; Doulami et al. 2021; Dobó et al. 2024). For example, MBL binds with high affinity to the mannan fraction of the mannan-rich Candida albicans cell wall (Consuegra-Asprilla et al. 2024). A membrane CLR is the single-chain transmembrane molecule known as the mannose receptor (MR). The extracellular segment of MRs consists of two parts. One is the proximal membrane end, which is responsible for endocytosis and ligand transport. The second extracellular segment of MR is the distal membrane end of the cysteine-rich lectin domain, which recognizes sulphated carbohydrate conjugates. The MR ligand is a mannan-rich structure expressed by pathogens (Li and Wu 2021). The human MR (hMR) binds HIV on vaginal epithelial cells without using the conventional CD4 receptor. This binding induces the production of matrix metalloproteinase 9 (MMP9), which degrades extracellular matrix proteins, thereby increasing the risk of HIV entering the vaginal epithelium and being transmitted to distal cells (Jadhav et al. 2013). Dectin-1 and Dectin-2 are typical representatives of the CLR family. Dectin-1 is a type II transmembrane protein that is expressed e.g., in monocytes, macrophages, dendritic cells (DCs) or neutrophils. It has a signal transduction function via an immunoreceptor tyrosine-based activation motif (ITAM). Dectin-1 can recognize a variety of fungi, including the yeast C. albicans. Dectin-1’s ligand is β-1,3-glucan, which activates downstream signals via tyrosine kinase-dependent and independent pathways upon recognition and binding (Li and Wu 2021).
Unlike Dectin-1, Dectin-2 lacks an ITAM sequence and lacks signal transduction function. It mainly recognizes α-mannan in the fungal cell wall and can also identify lipoglycans from bacterial species (Li and Wu 2021).
Siglecs (sialic acid-binding immunoglobulin-type lectins) are a family of immune-regulatory receptors found on immune cells. The N-terminal V-set Ig-like domain of Siglecs mediates the identification of sialic acid residues on microbial glycoconjugates. Siglecs are divided into two subgroups: one that includes all CD33-related Siglecs and one that consists of Siglec-1 (sialoadhesin), Siglec-2 (CD22), Siglec-4 (myelin-associated glycoprotein, MAG), and Siglec-15. Interactions between Siglecs and their ligands may activate or inhibit the immune response (Bornhöfft et al. 2018). An example of a Siglec with inhibitory activity against immune cells is CD33 (Siglec-3), which down-regulates TLR4-mediated signaling by binding to CD14. The CD14/CD33 interaction influences the presentation of LPS from CD14 to TLR4, thereby downregulating the LPS-NF-κB pathway. Interestingly, SIGLEC-mediated negative regulation of TLR4 signaling has been observed in BV, thereby attenuating the immune system’s pro-inflammatory response. Sialic acid residues have been identified on the surface of bacterial capsules and C. albicans cells (Amabebe and Anumba 2022). Stimulation of human peripheral blood mononuclear cells (PBMCs) by C. albicans induced SIGLEC15 expression and a polymorphism in the SIGLEC15 gene among the PBMCs. SIGLEC15 expression increased after 4 and 24 hours of stimulation, but only the 24-hour time point remained significant. SIGLEC15 expression was higher in PBMCs and neutrophils than in CD4+ and CD14+ cells. Moderately high SIGLEC15 expression was observed in a vaginal epithelial cell line stimulated with different C. albicans morphotypes. Researchers observed minor differences in cytokine production by PBMCs after 24 hours of stimulation with neuraminidase-treated or untreated C. albicans cells. In contrast, stimulation of T cells for seven days with the C. albicans variant resulted in significant differences in IL-17A and IFN-γ. Intravaginal infection of C57BL/6 mice with C. albicans resulted in a significant increase in SIGLEC15 expression, and SIGLEC15 mRNA silencing led to increased fungal burden and polymorphonuclear leukocyte infiltration (Jaeger et al. 2019).
Specific pathogens exhibit a wide diversity of capsular structures, reflected in the different compositions and linkages of repeating sugar residues. The strategy of capsular molecular mimicry enables pathogens to evade the immune system, while antigenic diversity among capsule types presents a moving target (Fig. 2 A-B) (Varki et al. 2022).
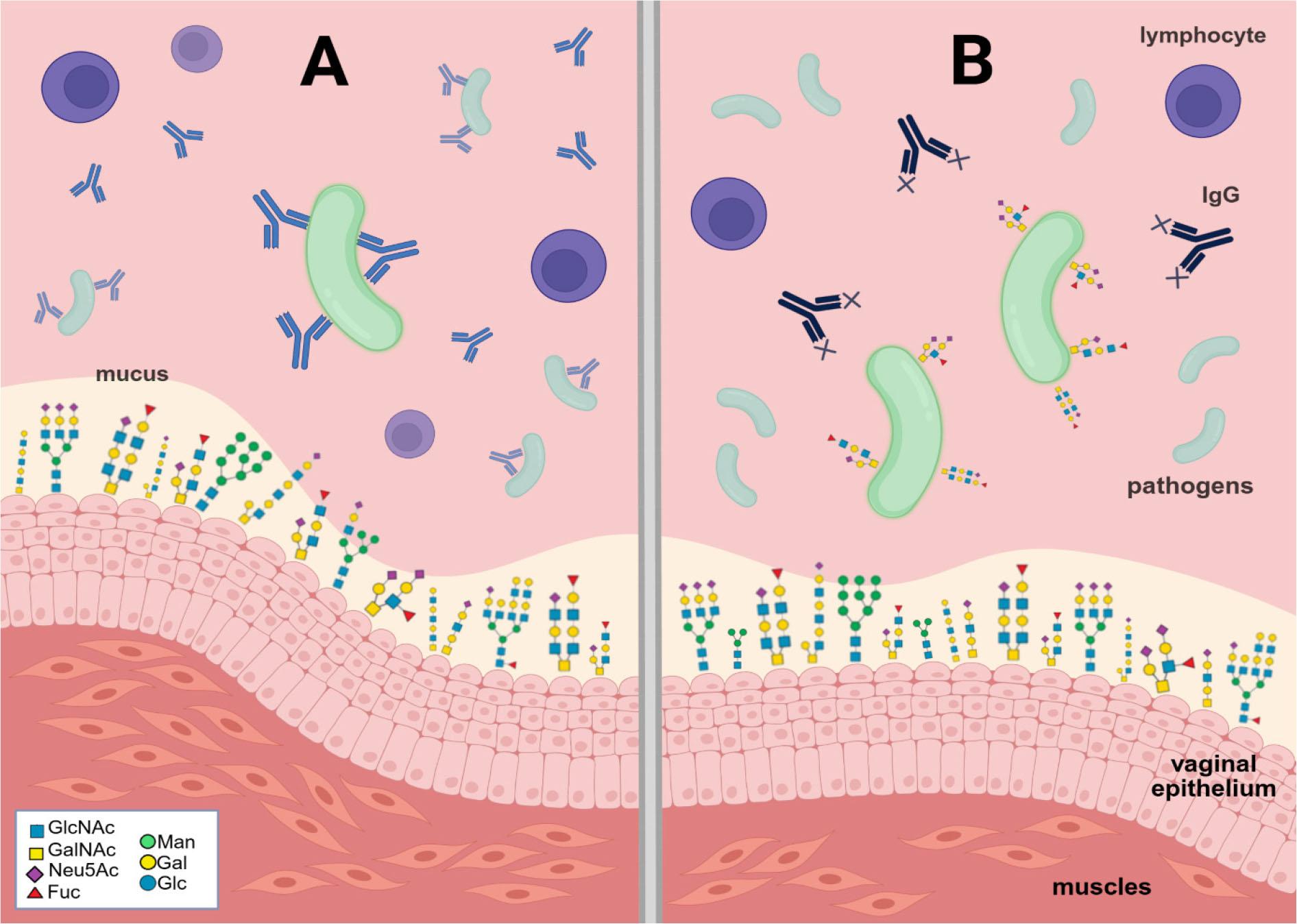
Bacterial evasion. A) Bacteria without antigen mimicry feature are actively inactivated by immune system. B)Antigen mimicry. The figure was created in BioRender.com (https://BioRender.com/zwlqbxx) (Bodaszewska-Lubas). Legend: Sugar residues forming the structures of N- and O-glycans: GlcNAc – N-acetylglucosamine; GalNAc – N-acetylgalactosamine; Neu5Ac – N-acetylneuraminic acid; Fuc – fucose; Man – mannose; Gal – galactose; Glc – glucose.
GBS strains are divided into nine serotypes (Ia, Ib, II-VIII) based on the sugar profile of capsule polysaccharides (Bodaszewska-Lubas et al. 2013). GBS involves sialic acid residues in a negative signaling mechanism known as sialoglycan mimicry. The capsular polysaccharides that determine the serotype of GBS present terminal Siaα2-3Galβ1-4GlcNAc units that are recognized as self-glycans by neutrophils (Bonnardel et al. 2021). Substantial evidence suggests that GBS may synthesize sialic acid de novo to produce a fully sialylated cell capsule. However, exogenous sialic acid is crucial for colonizing host niches and is used by bacterial cells as a carbon and energy source. GBS cannot produce sialidases to obtain sialic acid residues from mucosal glycoproteins and instead uses the products of sialidases from other bacteria that colonize the vaginal mucosa. Pezzicoli et al. (2012) suggested that GBS expresses the putative sialic acid-binding unit SAL0039, which may play a key role in sequestering this sugar from the environment and making it accessible for transport. Researchers created a mouse model of intravaginal GBS infection in the presence or absence of sialic acid. Vaginal colonization by the wild-type strain was poor (in 4 out of 10 animals), but adding sialic acid increased the number of infected animals to 9 out of 10. The importance of SAL0039-mediated transport of sialic acid during the colonization of mucosal surfaces, and the independence of bacterial adherence to cell monolayers from SAL0039, were also confirmed. Furthermore, GBS secretes hyaluronidases that degrade cervical hyaluronic acid into disaccharides. In this way, the bacteria dampen host immune activation by inhibiting Toll-like receptors, thereby potentially contributing to preterm birth via ascending infection (Pezzicoli et al. 2012).
Bacterial vaginosis (BV) is associated with a higher risk of infection with N. gonorrhoeae, the bacterium that causes gonorrhoea. The cytolytic activity of vaginolysin, produced by Gardnerella, causes the destruction of HeLa cells and the release of their contents. N. gonorrhoeae captures lysed intracellular metabolites and evaded host immunity via glycan molecular mimicry. An example of this mimicry is the capping of its lipooligosaccharide (LOS) with α-2,3-linked sialic acid, which is obtained from the surrounding environment. A higher level of bacterial sialylation is associated with increased binding of the complement regulatory protein factor H and greater resistance to complement attack (Morrill et al. 2023). The vaginal immunological response is modulated by bacteria associated with BV. IL-1β, a proinflammatory cytokine, can activate innate and adaptive immunity and appears to be a crucial cytokine in reproductive tract infection-related lesions. Its consistently elevated concentration has been described among BV- and HIV-infected women. Positive correlations were observed between IL-1β and IL-8, as well as a consistent correlation with the number of vaginal neutrophils, in both healthy individuals and those with BV infection. In cases of BV and Trichomonas vaginalis co-infection, much higher concentrations of the proinflammatory cytokines IL-1β and IL-8, as well as a higher number of neutrophils, were observed. Additionally, co-colonization with yeast does not affect IL-1β concentration but significantly increases IL-8 levels and can affect the number of neutrophils that arise. Furthermore, levels of sialidase, prolidase, and antibodies against G. vaginalis did not change during yeast co-infection. Vaginal Prevotella species have been associated with increased cytokine concentrations in CVF. P. timonensis induces a strong pro-inflammatory response through the activation of dendritic cells and increases HIV-1 uptake and storage by Langerhans cells (Cauci et al. 2007). The inflammatory response of the vaginal cell line VK2/E6E7 and the endocervical cell line End1/E6E7 was determined after incubation with the commensal strain L. crispatus and the pathogenic strains G. vaginalis, P. timonensis, and P. bivia. Only G. vaginalis significantly increased IL-1β, IL-8, and CCL20 mRNA in both cell lines compared to the control strain L. crispatus (Segui-Perez et al. 2024). Furthermore, sialylation plays a role in the immune response by altering immunoglobulin function and regulating inflammation (Chen et al. 2024). Women with BV, but not those with a LAB-dominated vaginal microbiota, have been found to exhibit sialidase activity in their vaginal fluids (Gilbert et al. 2019). Lewis et al. (2012) examined clinical BV specimens in combination with sIgA as a substrate for BV-associated glycosidases and proteases. The BV samples hydrolyzed sialic acid from sIgA, but not in the presence of dehydro-deoxysialic acid, which was used as a sialidase inhibitor. Terminal sialic acid residues on sIgA protect underlying carbohydrate residues from exposure and hydrolysis by exoglycosidases (galactosidase and hexosaminidase). A study among Danish pregnant women found that high levels of sIgA against the G. vaginalis hemolytic exotoxin (anti-Gvh IgA) protect against adverse pregnancy outcomes (Cauci et al. 2007).
Glycobiology is a new and fascinating field of science that is still developing. It continues to provide important insights into cellular mechanisms in health and disease. In turn, the glycan profile of the female genital tract is closely connected with the microbiota colonizing the vagina. The composition of the microbiota and the glycan profile have a significant influence on the health of the female genital tract, as well as on fertilization, the course of pregnancy, and term childbirth. Any disturbance to the microbiota or the glycan profile negatively impacts these processes. The mechanisms of pathogen virulence alter glycans in the CVM and CVF, creating an abundance of modified glycans that can be used for diagnosis as putative biomarkers of vaginal dysbiosis, infections, infertility, and preterm birth. Conversely, pathogen virulence factors may be targets for treating infections in cases of antibiotic resistance.