
Streptococcus agalactiae (also known as group B Streptococcus, GBS) is commonly found as part of the gastrointestinal and urogenital microbiota of healthy adults. GBS is associated with an increased risk of neonatal sepsis and meningitis, preterm labor, stillbirth, and pregnancy-related infections (Pangerl et al. 2021; Gergova et al. 2024). GBS is an opportunistic pathogen that causes various invasive infections in different locations (Hayes et al. 2020). It primarily affects elderly patients and non-pregnant immunocompromised adults and causes urinary tract infections, bloodstream infections, osteomyelitis, meningitis, endocarditis, pneumonia, and skin/soft tissue infections (Raabe and Shane 2019; Liu et al. 2023). Currently, most cases of invasive GBS disease occur in adults over 65 years old who have concomitant chronic non-infectious diseases such as diabetes, cirrhosis, malignancies, and decubitus ulcers (Stewart et al. 2020). S. agalactiae also causes mastitis and other infections in various animals (Oliveira et al. 2024).
The ability of S. agalactiae to cause infection is associated with the possibility of colonization, crossing of tissue barriers, evading host immune response, biofilm formation, and expression of virulence and pathogenicity factors (Silvestre et al. 2020). Capsular polysaccharide (CPS) is crucial for bacterial survival (Shabayek et al. 2018). GBS is classified based on capsular polysaccharides into ten serotypes (Ia, Ib, II–IX). Serotype III is considered the most virulent and common cause of late-onset meningitis in neonates, followed by serotype Ia, while Ia, V, and III are, according to some data, predominant in invasive infections in adult patients (Shabayek et al. 2018; Ali et al. 2020; Miselli et al 2022). The importance of classification into serotypes and their geographical distribution is related to developing vaccines against GBS in certain regions (Carreras-Abad et al. 2020; Campisi et al. 2021).
In recent years, the number of multidrug-resistant (MDR) GBS isolates has increased, including cases of isolates with significantly higher MICs than the susceptibility testing breakpoints to penicillins, which are the first line of choice for prophylaxis and prevention of neonatal infections, as well as invasive GBS disease in adults (Raabe and Shane 2019; Mudzana et al. 2021; Zhang et al. 2021). Second-line medications are macrolides and lincosamides. Clindamycin, a member of the lincosamides group, is often used to treat severe staphylococcal and streptococcal infections (Liu et al. 2023). Recent studies have shown that resistance to these antibiotics is becoming increasingly common (Plainvert et al. 2020; Awwad et al. 2021; Jin et al. 2022; Lohrmann et al. 2021; Alemán et al. 2022). The crossresistance phenotype to macrolides, lincosamides, and streptogramins is called MLSb. According to antibiotic non-susceptibility to clindamycin and erythromycin, there are four phenotypes: constitutive phenotype, concurrent erythromycin and clindamycin resistance profile (cMLSb), inducible phenotype, erythromycin resistance induces cross clindamycin resistance (iMLSb), M-phenotype (erythromycin resistance and clindamycin susceptible), and L-phenotype – a rare lincosamide resistance/macrolide susceptibility (LR/MS) phenotype (Hayes et al. 2020; Liu et al. 2023). Due to the potentially severe side effects in pregnant women and children, as well as the high rate of resistance observed, tetracyclines have limited use in human infections caused by S. agalactiae, but are still widely used in animals (Haenni et al. 2018; Gizachew et al. 2019; Khademi and Sahebkar 2020; Kekic et al. 2021; Mudzana et al. 2021; Gergova et al. 2024). In a previous study of Bulgarian S. agalactiae strains isolated between 2018 and 2019, their resistance was found to be 58.9% to erythromycin, 15.9% to clindamycin, and 94.6% to tetracycline (Gergova et al. 2021). Fluoroquinolones have good efficacy and elicit low bacterial resistance and are important for treating streptococcal infections in adults. However, recent studies have shown that Africa, Asia, North and South America are experiencing increasing rates of S. agalactiae resistant to quinolones (do Nascimento et al. 2019; Francois Watkins et al. 2019; Arias et al. 2022; Liu et al. 2023; Wang et al. 2023; Zakerifar et al. 2023). Moreover, levofloxacin nonsusceptibility GBS isolates have also been observed in Europe in recent years (Genovese et al. 2020, Gergova et al. 2021; Kekic et al. 2021; Lusta et al. 2023).
The study’s objective is to assess the prevalence of genes determining antibiotic resistance and capsule serotypes of Bulgarian clinical isolates of S. agalactiae from 2021 to 2024.
A total of 257 (n = 257) GBS isolates were obtained from inpatients and outpatients aged 17–88 years old during routine diagnostics. The specimens were collected from two University Hospitals in Sofia and one University Hospital in Pleven, Bulgaria, between September 2021 and May 2024. They are divided into two groups according to their source. The first significant group (1st group) consisted of vaginal samples – 191 (74.3%) collected from pregnant and non-pregnant women with confirmed genital infections with clinical manifestation and microscopic data such as either a coinfection pathogen in bacterial vaginosis or as the primary pathogen in aerobic vaginitis. Specimens with extra-vaginal localization represent the second group (2nd group) – 66 (25.7%), which are divided based on the sites of the collection as 2A) invasive specimens from normally sterile sites (36) such as blood cultures (4), soft tissue wounds aspirates (25), ear fluid aspirate (1), tracheal aspirates (6), 2B) non-invasive samples (30) taken from urine (20), sperm fluids (9), and eye discharge (1).
GBS was presumptively identified by colony and Gram staining morphology, positive tests of CAMP factor, negative tests of catalase and pyrrolidonyl arylamidase (PYR), lack of susceptibility to bacitracin, and positive latex-agglutination test for Lancefield B serological group (PathoDxtra Strep Grouping Kit, Oxoid™, Thermo Fisher Scientific, Inc., USA). If necessary, subsequent detailed biochemical identification was done with BD BBL™ Crystal™ Gram-Positive (GP) (Becton, Dickinson and Company, USA). GBS strains were stored in skim milk at –70°С and were sub-cultured three times on BD BBL™ Columbia Agar (Becton, Dickinson and Company, USA) with 5.0% sheep blood for 18–24h at 35°C in 5.0% CO2 atmosphere before the antibiotic susceptibility testing and other tests. Streptococcus pneumoniae ATCC® 49619™ and S. agalactiae ATCC® 27956™ were used as control strains according to EUCAST (2024) guidelines and in the molecular genetic experiments.
Pure streptococcal cultures were used for genomic DNA extraction, which was performed using an extraction kit (DNA-Sorb-A DNA extraction kit, (Sacace Biotechnologies Srl, Italy) in accordance with the manufacturer’s instructions. All DNA extractions were stored at –70°С until testing.
Antimicrobial susceptibility testing was performed using the Kirby-Bauer disk diffusion method and minimal inhibitory concentration (MIC). MLSb phenotype was detected with the disk-diffusion susceptibility method. Two antibiotic disks were used to determine antimicrobial susceptibility: erythromycin (15 μg) and clindamycin (2 μg). We determined the MIC values of erythromycin, clindamycin, and tetracycline for all isolates by E-tests (obtained from Laboratories Pvt. Ltd., India). The MICs of penicillin G and vancomycin were determined using a broth microdilution test (MIKROLATEST® MIC, Erba Lachema s.r.o., Czech Republic). We determined the MIC90 values for all the tested GBS strains as the lowest concentration of the expected antimicrobial agent at which 90.0% of the isolates were inhibited. For interpretations of the results of antibiotic testing, EUCAST recommendations were used (EUCAST 2024).
All collected GBS strains were confirmed by PCR using forward and reverse primers STRA-AgI and II, which target the 16S to 23S rRNA intergenic spacer region (Delannoy et al. 2013). PCR was used to determine the macrolide, lincosamide, and tetracycline resistance genes. The respective primer sequences are shown in Table I. The GBS capsular serotypes were detected by PCR using primers previously described by Poyart et al. (2007). We used prime Taq premix 2× (Genetbio Co., Ltd., Republic of Korea). The reaction conditions for conventional PCR were initial denaturation at 95°C for 5 min, followed by 30–35 cycles consisting of denaturation at 95°C for 30 sec, annealing for 30 sec, and elongation at 72°C for 1 min; final elongation at 72°C for 5–10 min. The result was read by gel electrophoresis (2.0% agarose) and GelRed® Nucleic Acid Gel Stain (Biotium, Inc., USA).
| Primer sequence (5’→3’) | Product size (bp) | Annealing temperature (°C) | Reference | |
|---|---|---|---|---|
| ermB | F: TGGTATTCCAAATGCGTAATG | 745 | 59 | Malhotra-Kumar et al. 2005 |
| ermA/TR | F: AACTTGTGGAAATGAGTCAACGG | 375 | 59 | Pérez-Trallero et al. 2007 |
| ermC | F: AATCGTCAATTCCTGCATGT | 297 | 55 | Duran et al. 2012 |
| msrD | F: TTGGACGAAGTAACTCTG | 371 | 53 | Daly et al. 2004 |
| mefA | F: AGTATCATTAATCACTAGTGC | 346 | 53 | Poyart et al. 2003 |
| mefE | F: ATGGAAAAATACAACAATTGGAAACGA | 1,218 | 58 | Arpin et al. 1999 |
| lsaC | F: CGGAATTGGGAAATCAACAC | 327 | 56 | Douarre et al. 2015 |
| lsaE | F: TGTCAAATGGTGAGCAAACG | 496 | 56 | Douarre et al. 2015 |
| lnuB | F: ACCAAAGGAGAAGGTGACCAA | 584 | 59 | Zhou et al. 2019 |
| tetM | F: AGTGGAGCGATTACAGAA | 159 | 55 | Strommenger et al. 2003 |
| tetO | F: AACTTAGGCAATCTGGCTCAC | 515 | 55 | Zakerifar et al. 2023 |
F – forward primer, R – reverse primer
Statistical analyses were carried out with IBM SPSS Statistics for Windows v19.0 (IBM Corp., USA). A p-value ≤ 0.05 was considered statistically significant.
Of the total number of isolates (n = 257), serotypes Ia (26.5%), III (20.2%), and V (19.8%) were the most common. Twenty-two strains were non-typeable (8.6%). The distribution of predominant serotypes in vaginal samples (n = 191) was as follows: Ia (29.8%), V (21.4%), and III (16.1%). Respectively, in extra-vaginal samples among the invasive strains (n = 36), the prevailed serotype was II (27.8%), followed by III (22.2%) and Ia (19.4%). In the non-inva-sive group (n = 30), serotypes Ia (23.3%) and III (23.3%) were the most common. There was no statistical significance between the distribution of serotypes in vaginal and extra-vaginal samples (p> 0.05) (Table II).
| Serotypes | Vaginal samples (n = 191) | Extra-vaginal samples | Total number (n = 257) | p-value (vaginal/extravaginal samples) | ||
|---|---|---|---|---|---|---|
| Invasive (n = 36) | Non-invasive (n = 30) | Total extravaginal (n = 66) | ||||
| Ia | 54 (28.3%) | 7 (19.4%) | 7 (23.3%) | 14 (21.2%) | 68 (26.5%) | 0.3317 |
| Ib | 2 (1.0%) | 0 | 0 | 0 | 2 (0.8%) | 1.0000 |
| II | 29 (15.2%) | 10 (27.8%) | 4 (13.3%) | 14 (21.2%) | 43 (16.7%) | 0.2572 |
| III | 37 (19,4%) | 8 (22.2%) | 7 (23.3%) | 15 (22.7%) | 52 (20.2%) | 0.5952 |
| IV | 13 (6,8%) | 1(2.8%) | 4 (13.3%) | 5 (7.6%) | 18 (7.0%) | 0.7852 |
| V | 41 (21.5%) | 6 (16.7%) | 4 (13.3%) | 10 (15.2%) | 51 (19.8%) | 0.2897 |
| VI | 0 | 0 | 0 | 0 | 0 | |
| VII | 1 (0.5%) | 0 | 0 | 0 | 1 (0.4%) | 1.000 |
| VIII | 0 | 0 | 0 | 0 | 0 | |
| NT | 14 (7.3%) | 4 (11.1%) | 4 (13.3%) | 8 (12.1%) | 22 (8.6%) | 0.3054 |
NT – non-typeable
p-value < 0.05 is considered statistically significant
All examined isolates showed susceptible MICs to penicillin and vancomycin. Among tested strains resistance to macrolides was revealed in 155 isolates (60.3%). There were 64 (24.9%) and 229 (89.1%) strains resistant to lincosamides and tetracyclines, respectively. The nonsusceptible strains isolated from different clinical materials were as follows: from vaginal samples – resistant to erythromycin (57.6%), clindamycin (23.0%), and tetracycline (90.1%), and from extra-vaginal specimens – resistant to erythromycin (68.2%), clindamycin (30.3%), and tetracycline (86.4%). There was no statistical significance between the distribution of resistance in vaginal and extra-vaginal samples (p> 0.05) (Table III).
| Antibiotics | Vaginal samples (n = 191) | Extra-vaginal samples | Total number (n = 257) | p-value (vaginal/extra-vaginal samples) | ||
|---|---|---|---|---|---|---|
| Invasive (n = 36) | Non-invasive (n = 30) | Total extra-vaginal (n = 66) | ||||
| ERY | 110 (57.6%) | 24 (66.7%) | 21 (70.0%) | 45 (68.2%) | 155 (60.3%) | 0.1459 |
| CLI | 44 (23.0%) | 8 (23.5%) | 12 (40.0%) | 20 (30.3%) | 64 (24.9%) | 0.2508 |
| TET | 172 (90.1%) | 31 (86.1%) | 26 (86.7%) | 57 (86.4%) | 229 (89.1%) | 0.4914 |
p-value < 0.05 is considered statistically significant
The strains in this study represented the following macrolide/lincosamide resistance phenotypes: cMLSb – 74 strains (47.4%), iMLSb – 48 strains (30.8%), M-type – 33 strains (21.2%), and L-type – one strain (0.6%). The distribution of the above phenotypes was 44.2%, 31.0%, 24.8%, and 0.0% in the vaginal group and 53.3%, 33.3%, 11.1%, and 2.2% in the extra-vaginal group.
Nine genes were detected to be associated with macrolide and lincosamide resistance. Their frequency was as follows: ermB (54.2%) >ermA/TR (30.3%) >mefA (20.7%) >ermC (18.1%) >msrD (14.8%) >mefE (8.4%), lsaC (8.4%) >lnuB (7.7%) >lsaE (6.5%). The indicated genes were the only resistance determinants in 63.8% of all ermA/Tr-positive isolates and 56.0% of all ermB-positive isolates. The genes mefE, msrD, lsaC, and lsaE always appeared in combination with other genetic resistance elements. The most frequently observed combinations between genes were ermB + ermC, ermB + lcaC, mefA + mefE, mefA + msrD, lnuB + lsaE (Table IV). Regarding tetracycline resistance, we detected two genes: tetM (89.1%) and tetO (14.4%). Both genes were found in eight isolates (3.5%). The main genetic profiles identified in resistant strains were: ermB + tetM, ermA/ TR + tetM, mefA + tetM and ermB + tetO (Table IV).
| ermB (n = 84) | ermА/TR (n = 47) | ermC (n = 28) | mefA (n = 32) | mefE (n = 13) | msrD (n = 22) | lsaC (n = 13) | lsaE (n = 10) | lnuB (n = 12) | tetM (n = 204) | tetO (n = 33) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ermB | 56.0%* | 12.8% | 50.0% | 3.1% | – | 4,5% | 92,3% | 70.0% | 58,3% | 24.5% | 60.6% |
| ermА/TR | 7.1% | 63.8%* | 39.3% | 18.8% | – | – | 7,7% | – | – | 17.2% | 6.1% |
| ermC | 16.7% | 23.4% | 10.7%* | 18.8% | 7.7% | 13.6% | 7,7% | – | – | 10.8% | 18.2% |
| mefA | 1.2% | 12.8% | 21.4% | 9.4%* | 100.0% | 95.5% | – | – | – | 12.7% | 9.1% |
| mefE | – | – | 3.6% | 43.8% | – | 54.5% | – | – | – | 5.9% | – |
| msrD | 1.2% | – | 10.7% | 65.6% | 92.3% | – | – | – | – | 9.3% | 9.1% |
| lsaC | 14.3% | 2.1% | 3.6% | – | – | – | – | 30.0% | 25.0% | 4.9% | 18.2% |
| lsaE | 8.3% | – | – | – | – | – | 23.1% | – | 83.3% | 3.4% | 9.1% |
| lnuB | 8.3% | – | – | – | – | – | 23.1% | 100.0% | 8.3%* | 3.9% | 9.1% |
| tetM | 59.5% | 76.6% | 78.6% | 81.3% | 92.3% | 86.4% | 76.9% | 70.0% | 66.7% | 41.2%* | 24.2% |
| tetO | 23.8% | 4.3% | 21.4% | 9.4% | – | 13.6% | 46.2% | 30.0% | 25% | 3.9% | 9.1%* |
* resistant isolates in which only one gene was detected
Among all macrolide-resistant strains (n = 155), the most common serotypes were Ia (25.8%), III (23.9%), and V (21.9%). In clindamycin-resistant isolates (n = 64), the same serotypes were prevalent: Ia (25.0%), III (21.9%), and V (18.8%). Regarding serotype distribution in tetracycline-resistant isolates (n = 229), serotype Ia was the leading type (24.9%), followed by V (21.4%) and III (19.7%) (Fig. 1). In the extra-vaginal strains serotypes Ia, III, and V were most common in strains resistant to macrolides and lincosamides and II, III, and Ia in ones resistant to tetracyclines.
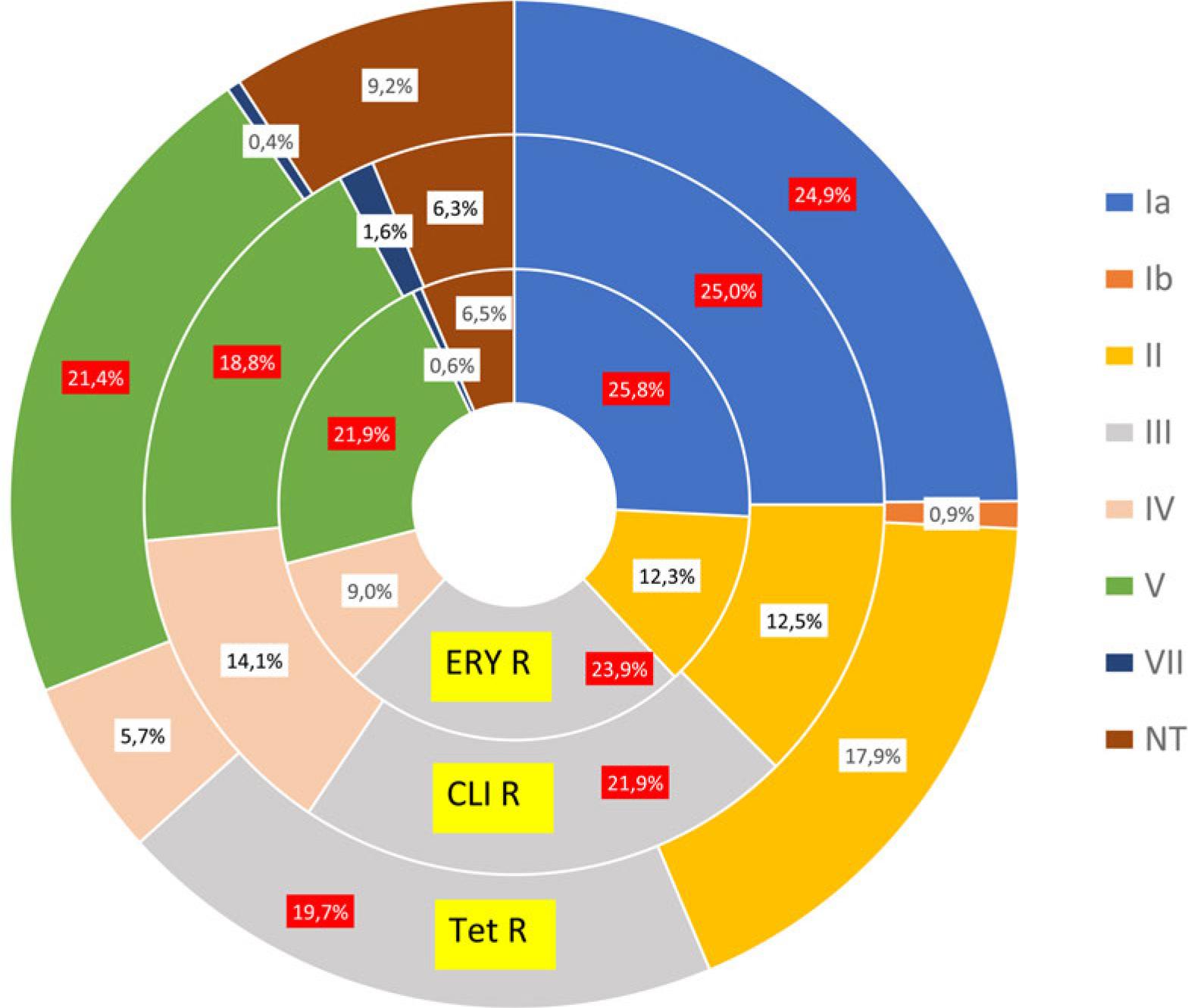
Association between resistance and serotype – total number. The predominant serotypes are colored red.
A particular resistance phenotype was most commonly associated with the following serotypes: cMLSb (Ia, V, and III), iMLSb (III, V, and IV), M-type (Ia, V, and II) and L-type (Ia) (Fig. 2). Regarding to the extravaginal group, these associations were: cMLSb (Ia, V, and IV), iMLSb (III, V, and II), M-type (Ia and V), and L-type (Ia).
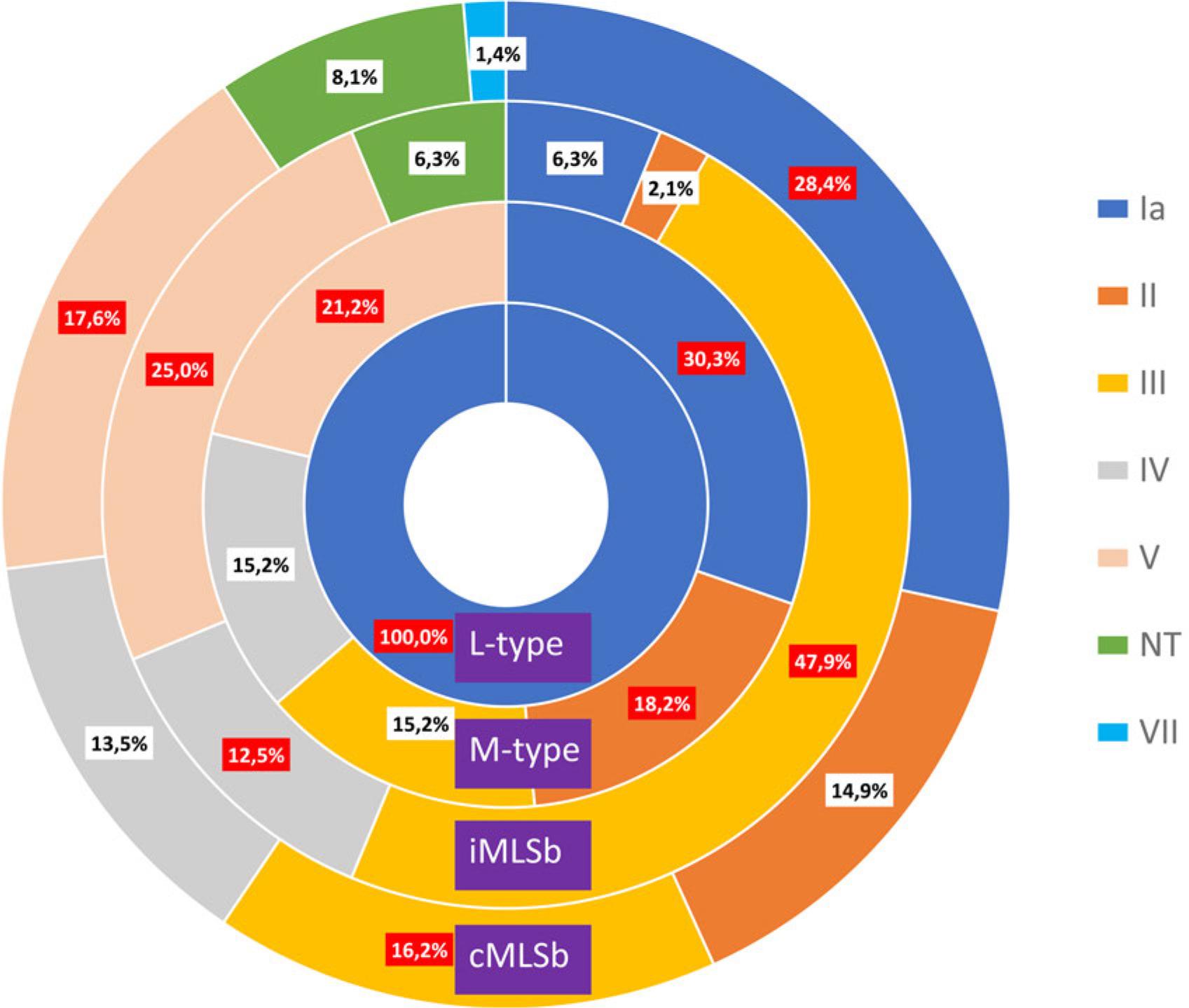
Association between phenotype and serotype – total number. The predominant serotypes are colored red.
In the current study, we examined the genes responsible for the main mechanisms of resistance to macrolides and clindamycin: 1) the erm genes, which products cause a modification in the fixation site in ribosomes; 2) the lsa and msr family of the genes associated with the production of ABC transporters; 3) the mef genes encoding efflux pumps that lead to bacterial resistance to macrolides alone; 4) the presence of lnu family genes leads to adenylation of the clindamycin molecule (Liu et al. 2023). The results showed that most strains harbored at least one erm gene, determining the prevalence of cMLSb (47.4%) and iMLSb (30.8%) phenotype. They are associated with high levels of antibiotic resistance due to a target modification that cross-reacts with macrolides, lincosamides, and streptogramins. A high percentage of resistant isolates virtually eliminates respective groups of antibiotics from treatment regimens. One strain had the rare L-phenotype with the only positive gene lnuB, confirming the association between them reported in other sources (Zhou et al. 2019).
In the previous study, 107 GBS strains (82 vaginal and 25 extra-vaginal samples) were isolated from Bulgarian patients in 2018–2019. The resistance rates to erythromycin and clindamycin were 58.9% and 15.9%, respectively (Gergova et al. 2021). In comparison, the current study reveals an increase of 1.4% and 9.0% in macrolide and lincosamide resistance, respectively. Similar results on resistance dynamics to macrolides/lincosamides have been reported in the last decade in some European countries such as Denmark 23.8%/26.0%, Portugal 16.1–35.1%/14.2– 33.9%, and Serbia 23.1–26.7%/22.1%. In the USA, the reported resistance rate for the same period was up to 54.8%/43.2%, in Argentina 25.0%/26.0%, and in Brazil 25.9%/5,2% (Francois Watkins et al. 2019; Arias et al. 2022; Barros et al. 2024). The problematic trend of increased resistance among invasive GBS isolates has been reported in China and Taiwan, with rates ranging from 78.0% and 49–68.0%, respectively, for macrolides, and 68.0% and 51.4–66.0%, for lincosamides (Khademi and Sahebkar 2020; Liu et al. 2023; Wang et al. 2023; Gergova et al. 2024).
Genes of the tet family mediate the main mechanisms of resistance of GBS to tetracyclines: the tetK and tetL genes encode efflux pumps, while the predominant genes tetM and tetO encode ribosomal protection proteins (Haenni et al. 2018; Haimbodi et al. 2021). Conjugative elements and transposons are linked to multidrug resistance as they contain genes that cause reduced susceptibility to various classes of antibiotics. Combined resistance to macrolides and tetracycline is usually associated with inserting ermB into transposons carrying tetM (Zhou et al. 2017; Khan et al. 2023; Gergova et al. 2024). In the current study, 59.5% of the isolates with ermB also possessed the tetM gene, indicating a possible association between tetracycline and macrolide resistance. Resistance to tetracycline was 89.1%, raising concerns about the role of tetracyclineresistant isolates in the spread of macrolide resistance among GBS (Gergova et al. 2024). Similar results were found in many countries after 2010, with resistance rates ranging from 68.9–80.1% in China, 83,9–86% in North and South America, 82.6% in Africa, in Europe from 81.6% in Iceland to 85.8% in Portugal, 86.0% in Serbia, 86.5–91.0% in France, and 94,6% in Bulgarian study (Botelho et al. 2018; Francois Watkins et al. 2019; Plainvert et al 2020; Gergova et al. 2021; Kekic et al 2021; Arias et al. 2022; Gergova et al. 2024).
Beta-lactams are the first-line drugs used to treat GBS-associated infections. Rare cases of GBS strains with low penicillin susceptibility have been reported in humans and cattle due to amino acid substitutions in PBPs (Hayes et al. 2020; Oliveira et al. 2024). For some GBS strains, we observed unusual growth near the MIC breakpoint (still in susceptibility values) for penicillin and vancomycin. Japanese authors reported a high prevalence of GBS isolates with MICs close to penicillin breakpoints, which increased over time and contributed to the rise of MDR strains non-susceptible to most common classes used for treatment of GBS-associated infections (Seki et al. 2015).
GBS serotype division is based on capsular polysaccharides. Capsules are one of the most important virulence factors involved in antigen mimicry, decreasing the efficacy of phagocytosis and mediation of biofilm formation (Shabayek et al. 2018; Campisi et al. 2021). Ia, III, and V were the most common serotypes in the present study. Only three strains were of serotype Ib, and no isolates were of VI or VIII serotypes. Bacterial serotypes I–V were identified in 98.0% of colonizing GBS isolates worldwide, while serotypes VI–IX were more common in Asia (Russell et al. 2017). The most common serotypes in Europe were III, Ia, and V; in Africa – serotypes II, V, III, and Ia, in Asia – Ia, III, V, in North America Ia, V, and II and in South America Ia, II, and III (Francois Watkins et al. 2019; Bianchi-Jassir et al. 2020; Slotved et al. 2021; Maria Silva et al. 2023; Barros et al. 2024). Among invasive infant strains, the most common isolates were III, Ia, and V, with III being the most common on all continents.
In contrast, Zhang et al. (2021) reported that type Ib/ST10 strains were prevalent in late-onset neonatal diseases in China. In the elderly population, invasive isolates were mainly Ia, V, and Ib (Bianchi-Jassir et al. 2020). Our research has confirmed the distribution of the most common serotypes in Europe and all but one isolate belonging to group Ia–V. Non-typeable isolates accounted for 8.6% due to the PCR method producing more non-typeable GBS than sequencing (Russell et al. 2017). As in the current study, no significant association between serotypes and source was found in South America. In contrast to our results, tetracycline resistance was significantly associated with the colonization of isolates (Hernandez et al. 2022).
The development of antibacterial resistance and the emergence of MDR-GBS strains (simultaneously resistant to three or more different classes of antimicrobial agents) indicate the need for more strict antibiotic politics, prompt measures to reduce uncontrolled antibiotic administration, and the introduction of new and more effective antibiotic molecules in clinical practice. The frequent combination of the most problematic serotypes III and Ia with MLSb resistance represents an alarming potential for the development of common invasive streptococcal diseases in Bulgaria. The relationship between serotype distribution and antimicrobial non-susceptibility conducted in the current research is of important value to indicate the importance of developing vaccines against the most common serotypes according to geographical prevalence. This work provides a foundation for future studies aimed at monitoring, preventing and controlling the development of resistant and MDR GBS.