
Acinetobacter baumannii is a Gram-negative opportunistic pathogen that often causes diseases such as pneumonia, bacteremia, and urinary tract infections in clinical practice (Harding et al. 2018). Our previous research found that this bacterium infects and enters host cells. Its particular pathogenic factors, outer membrane protein A and outer membrane protein 34, can cause host cell pyroptosis and autophagy, leading to the release of inflammatory factors and causing damage to host cells (An et al. 2019a; 2019b). In recent years, research on A. baumannii and its virulence factors has further revealed how this bacterium interacts with host cells to manipulate the inflammatory response. Scientists are trying to find ways to cope with its infection by combating A. baumannii itself and regulating the host immune response (Li et al. 2018; Zhao et al. 2021; Meng et al. 2024).
Cuprotosis is a newly discovered form of programmed cell death, which differs from apoptosis, pyroptosis, and ferroptosis. This process depends on changes in intracellular copper concentration. The primary mechanism that triggers copper death is the abnormal increase of copper ions in cells, which promotes their binding to thioacyl proteins in the tricarboxylic acid cycle (TCA), causing abnormal oligomerization of thioacyl proteins and a decrease in iron sulfur cluster protein levels, inducing protein toxicity stress and leading to cell death (Li et al. 2022). In the process of cuprotosis, there is a critical upstream regulatory protein – ferredoxin 1 (FDX1), a reductase and a direct target of copper ions. It can reduce Cu2+ to a more toxic Cu+. Knocking out FDX1 can reduce the acylation of lipoacylated proteins DLAT (dihydrolipoyltransferase) and DLST (dihydrolipoylsuccinyltransferase), preventing them from binding to copper ions and protecting cells from copper toxicity (Tsvetkov et al. 2022). However, it is currently unclear whether cuprotosis occurs during the infection of host cells by A. baumannii.
LncRNA-GAS5, also known as growth arrestspecific transcription factor 5, is located at 1q25 and includes 12 exons encoding GAS5a and GAS5b, as well as 11 introns encoding small nucleolar RNA (snoRNA) (Kino et al. 2010; Yang et al. 2020). LncRNA-GAS5 regulates the cell cycle and programmed cell death through endogenous competitive RNAs or interactions with transcription factors and regulates cellular immune function (Hu et al. 2014; Gao et al. 2021). Our previous research has shown that LncRNA-GAS5 degrades the synaptic fusion protein syntaxin 17 (STX17), leading to disrupted autophagy degradation and exacerbating the cellular inflammatory response mediated by A. baumannii and acute lung injury in mice with sepsis (An et al. 2021). However, the role of A. baumannii infection on cuprotosis protein FDX-1 and the regulatory role of LncRNA-GAS5 in this process is currently unclear.
A. baumannii ATCC® 19606™ was purchased from ATCC® in the United States. The pcDNA3.1-GAS5 overexpression plasmid and sh-GAS5 knockdown plasmid were previously constructed and preserved in our laboratory. The human embryonic kidney cell line HEK293T was also preserved in our laboratory. The Lipofectamine™ 3000 transfection kit was sourced from Invitrogen™ (Thermo Fisher Scientific, Inc., USA); FDX-1 was purchased (ab108257) from Abcam Limited (UK); goat anti-rabbit or anti-mouse fluorescent secondary antibodies were purchased from LI-COR Biosciences in the United States. The Veriti™ and Real time PCR (7500) instruments were products of Applied Biosystems™ (Thermo Fisher Scientific, Inc., USA). Nucleic acid and protein electrophoresis apparatus, electrophoresis tank, and gel imaging system were all products of BioRad Laboratories, Inc. (USA). The Odyssey® dual-color infrared fluorescence imaging system was purchased from LI-COR Bioscences (USA).
The methods for culturing and infecting HEK293T cells with A. baumannii were as follows: prefrozen A. baumannii was rapidly thawed and resuspended in a liquid LB medium for 8 hours. Then, 1 × 107A. baumannii was used to infect 1 × 106 HEK293T cells at different times.
Transfect HEK293T cells with overexpression plasmids pc3.1-GAS5 or knockdown plasmids sh-GAS5 with a concentration of 1 μg/μl using Lipofectamine™ 3000 liposome transfection reagent. The transfection test dose for each well of the 6-well plate was determined according to the instructions of the reagent. 500 μl of Opti-MEM™ (Gibco™, Thermo Fisher Scientific, Inc., USA), 7.5 μl of lipo3000 reagent, 10 μl of p3000 reagent, and 5 μg of overexpression plasmids pc3.1-GAS5 or knockdown plasmids sh-GAS5 were taken to form the transfection system. Mix well and let stand for 15 minutes. Afterwards, HEK293T cells in the logarithmic growth phase were added and collected 48 hours later.
HEK293T cells were collected from different groups infected with A. baumannii at different times, and those overexpressing or knocking down LncRNA-GAS5 before reinfecting A. baumannii. RIPA lysis solution was used to lyse cells on ice for 10 minutes, followed by centrifugation at 12,000 × g for 10 minutes; the BCA method was used for protein quantification. SDS-PAGE gel of 15% concentration was used for electrophoresis, and then the protein was transferred to the 0.22 μm PVDF Millipore membrane (Merck KGaA, Germany). The membrane was sealed at room temperature for 1 h with 5% skimmed milk powder, cut into the target strip and incubated with FDX-1 antibody (diluted with 1:1000) at 4°C overnight, The next day, the membrane was washed with TBST for three times, 10 min each time, then incubated with fluorescent labeled goat anti-rabbit antibody (diluted with 1:15000) at room temperature for 1 h, washed with TBST for three times, 10 min each time. Finally, the detection of protein expression was used by Odyssey® CLx equipment (LI-COR Bioscences, USA).
Data was conducted using IBM SPSS Statistics for Windows v17.0 (IBM Corp., USA), and the results were expressed as mean ± standard deviation. GraphPad Prism version 5.0 (GraphPad Software, USA, www.graphpad.com) was used as a bar chart; Student’s t-test was used for significance analysis, and *p < 0.05 or #p < 0.05 were considered statistically significant.
Firstly, we infected HEK293T cells with A. baumannii with MOI = 10 for 4 h, 8 h, and 16 h, and then detected the expression of FDX-1 using Western blot. The results showed that infection with A. baumannii led to a decrease in FDX-1 expression in HEK293T cells, significantly different from the uninfected group (p < 0.05), but not time-dependent. This proves that A. baumannii inhibits cuprotosis in HEK293T cells (Fig. 1).
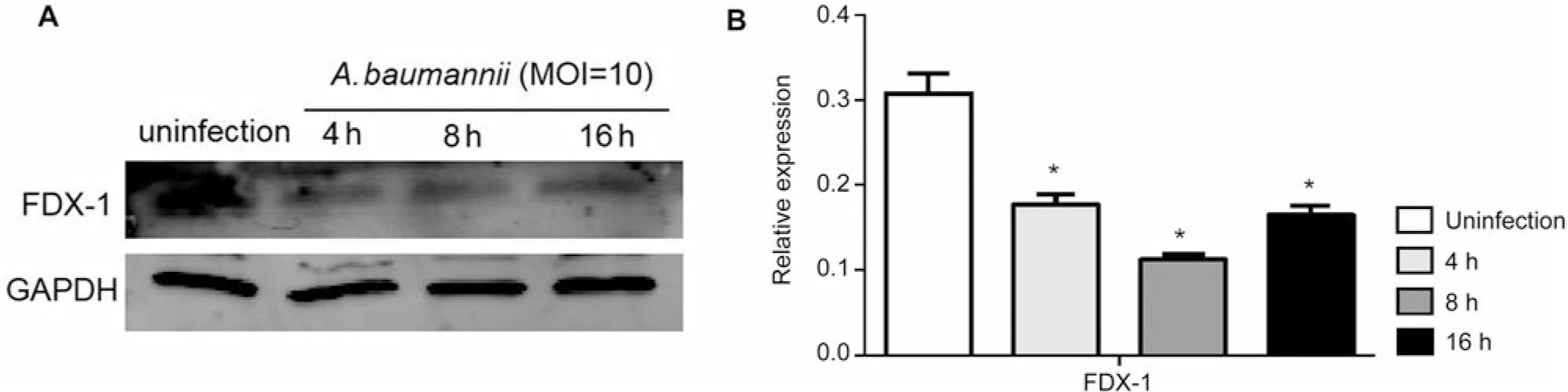
Acinetobacter baumannii inhibits the expression of copper death protein FDX-1 in HEK293T cells.
A) Western blot was used to detect the expression of FDX-1 protein; B) using three independent experiments as statistical analysis data, there was a significant difference in FDX-1 protein expression compared to the uninfected group, *p < 0.05.
Next, we used Western blot to detect the expression of FDX-1 protein and found that overexpression of LncRNA-GAS5 can inhibit FDX-1 expression, which is significantly different from the empty plasmid group (pcDNA3.1), *p < 0.05 (Fig. 2A and 2B). Knocking down LncRNA-GAS5 can enhance FDX-1 expression, with significant differences compared to the empty plasmid group (sh-NC), *p < 0.05 (Fig. 2C and 2D). This result demonstrates that LncRNA-GAS5 has a regulatory effect on the expression of FDX-1.
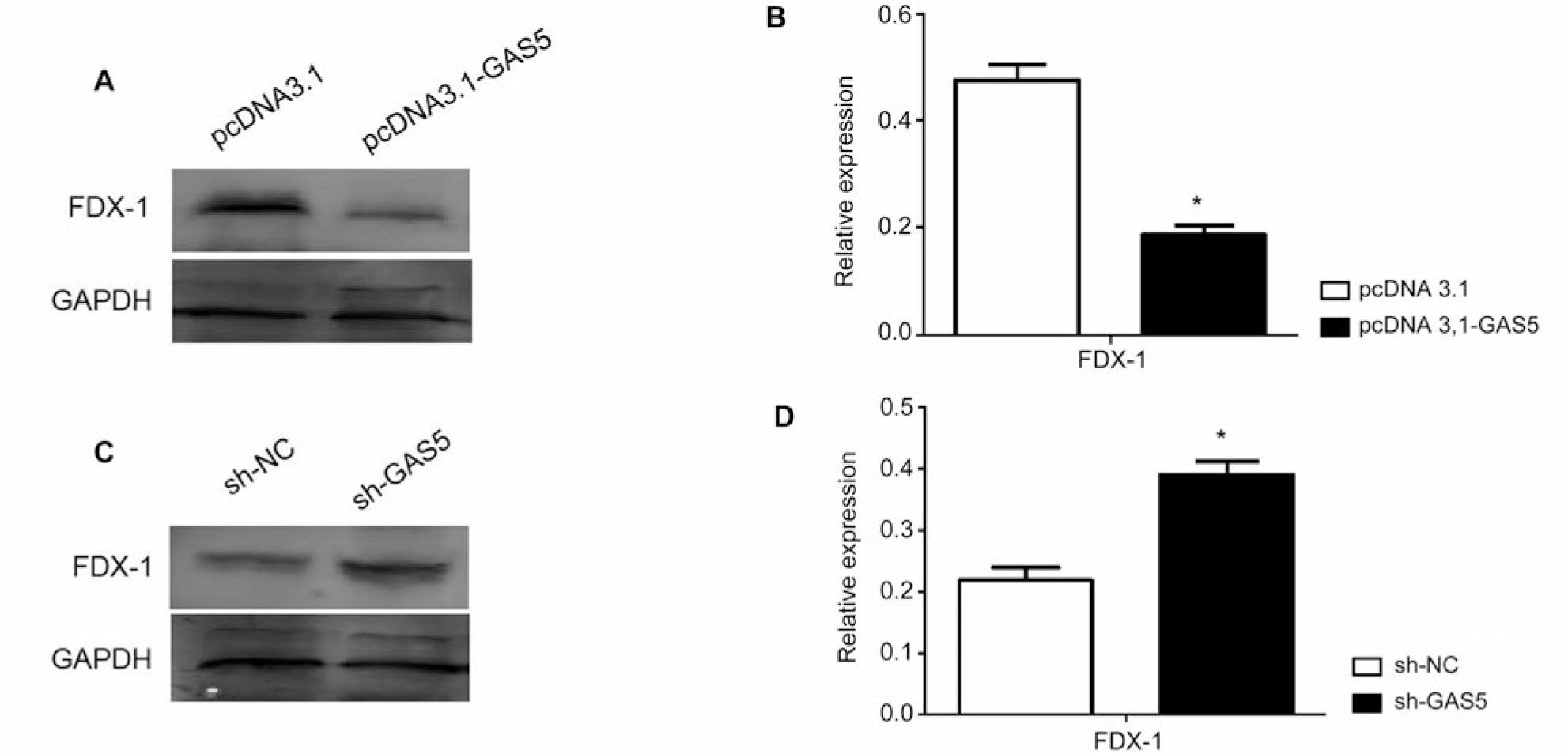
LncRNA-GAS5 regulates the expression of copper death protein FDX-1.
A) Transfection of HEK293T cells with pcDNA3.1 or pc3.1-GAS5 for 48 hours and detection of the FDX-1 protein expression using Western blot; B) using three independent experiments as statistical analysis data. The expression of FDX-1 showed significant differences between the pcDNA3.1-GAS5 transfection group and the pcDNA3.1 transfection group, *p < 0.05; C) transfection of HEK293T cells with sh-NC or sh-GAS5 for 48 hours was used to detect the expression of FDX-1 protein using Western blot; D) using three independent experiments as statistical analysis data. The expression of FDX-1 showed significant differences between the sh-GAS5 transfection group and the sh-NC transfection group, *p < 0.05.
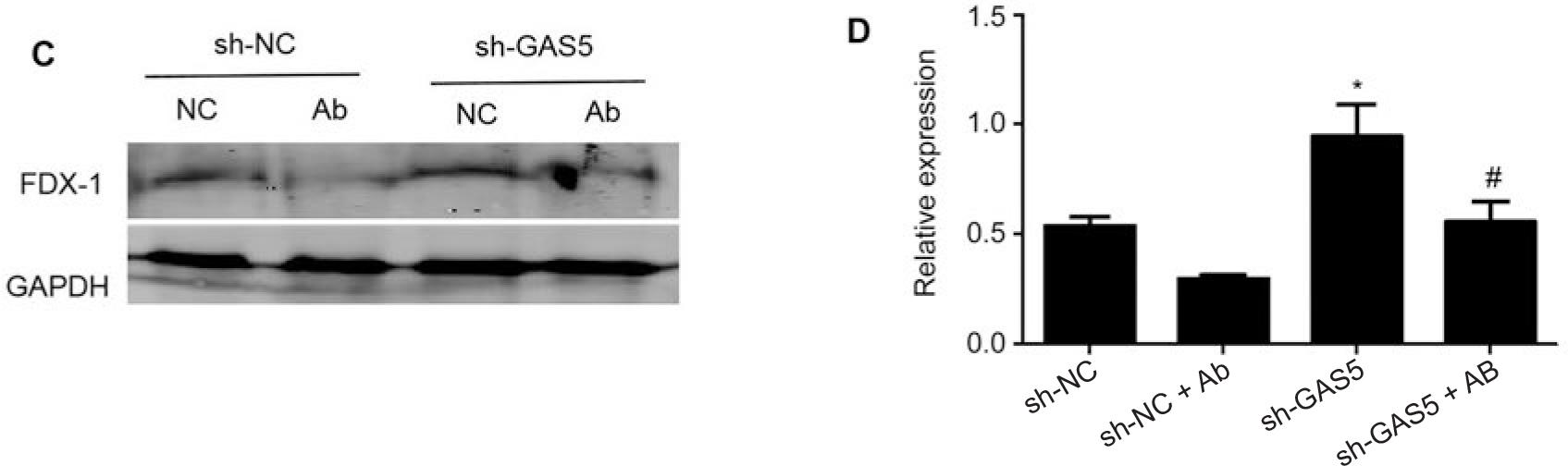
Finally, overexpression of LncRNA-GAS5 in HEK293T cells and subsequent infection with A. baumannii significantly enhances the inhibitory effect of A. baumannii on FDX-1 (Fig. 3A and 3B). On the contrary, knocking down LncRNA-GAS5 in HEK293T cells and then infecting A. baumannii revealed that knocking down LncRNA-GAS5 could restore the inhibitory effect of A. baumannii on FDX-1 (Fig. 3C and 3D). This result demonstrates that knocking down LncRNA-GAS5 restores the inhibitory effect of A. baumannii on FDX-1.
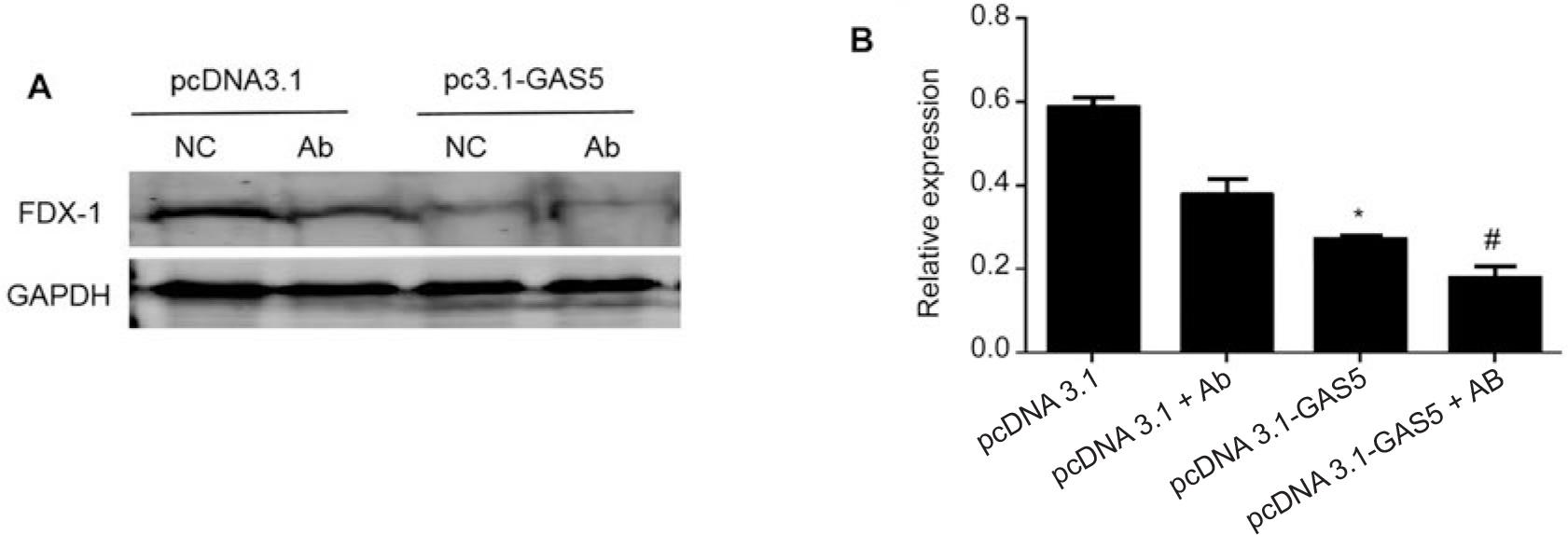
LncRNA-GAS5 regulates the inhibitory effect of Acinetobacter baumannii on the expression of copper death protein FDX-1.
A) After transfection of HEK293T cells with pcDNA3.1 or pcDNA3.1-GAS5 for 48 hours, the cells were infected with Acinetobacter baumannii with MOI = 10 for 4 hours, and the expression of FDX-1 protein was detected by Western blot; B) using three independent experiments as statistical analysis data, there were significant differences in FDX-1 protein expression between the empty plasmid group and the group infected with Acinetobacter baumannii alone or between the pcDNA-3.1-GAS5+Ab group and the pcDNA3.1-GAS5 transfection group, *p < 0.05 or #p < 0.05, respectively; C) after transfection of HEK293T cells with sh-NC or sh-GAS5 for 48 hours, the cells were infected with Acinetobacter baumannii with MOI = 10 for 4 hours, and the expression of FDX-1 protein was detected by Western blot; D) using three independent experiments as statistical analysis data, there were significant differences in FDX-1 protein expression between the pcDNA-3.1 group and the group infected with Acinetobacter baumannii alone, or between the sh-GAS5+Ab group and the sh-GAS5 transfection group *p < 0.05 or #p < 0.05, respectively.
Previous studies have found that LncRNA-GAS5 has the function of regulating gene transcription, and its forms of action can be divided into two. The first form is that LncRNA-GAS5 can directly bind to target genes to regulate gene expression. The second form is its indirect regulation of gene transcription by combining with specific regulatory proteins to form complexes (Zhang et al. 2019; Li et al. 2020). We found that overexpression of LncRNA-GAS5 inhibited FDX-1 expression while knocking down LncRNA-GAS5 activated FDX-1 expression. Knocking down LncRNA-GAS5 promotes FDX-1 expression and restores the inhibition of FDX-1 expression caused by A. baumannii. This result indicates that knocking down LncRNA-GAS5 can be a new target for treating A. baumannii infection. In the future, this method can regulate the inhibition of cuprotosis by A. baumannii, thereby enhancing the inflammatory defense of cells against this bacterium. However, further research is needed on how LncRAN-GAS5 binds to FDX-1 and regulates its expression.
In addition, we have preliminarily found that A. baumannii inhibit cuprotosis regulatory protein FDX-1, and LncRNA-GAS5 has a regulatory effect on this process. At present, detecting changes in copper ion levels in cells can be used as an indicator to evaluate cuprotosis. However, we still need clarification about the effect of A. baumannii on changes in copper ion levels in cells. In the future, we will use ICP-OES (Inductively Coupled Plasma Optical Emission Spectroscopy) to detect changes in copper ions in cells, which can further determine the role of A. baumannii in cuprotosis and the regulatory role of LncRNA-GAS5 in this process.
In conclusion, this study found that infection of HEK293T cells with A. baumannii inhibited cuprotosis regulatory protein FDX-1, which is different from the previously discovered forms of programmed cell death mediated by A. baumannii. We preliminarily infer that A. baumannii may promote its own proliferation and survival by inhibiting cuprotosis, which may be another new mechanism by which it manipulates inflammatory cell death. Furthermore, it is worth further investigating whether the infection process of A. baumannii interferes with other molecules involved in cuprotosis.