
Lignocellulose is a significant plant component and varies among plant species (de Gonzalo et al. 2016). The demand for value-added products such as organic acids, biofuel, and microbial polysaccharides from lignocellulose biomass is increasing worldwide (Chaturvedi and Verma 2013). It is due to the renewable and abundant nature of the lignocellulose biomass. However, the recalcitrant nature of lignin remains a major bottleneck to unleashing useful products during various industrial operations. Due to inadequate treatment and waste management plans, lignin waste biomass from industrial and agricultural sectors continue to pose an environmental threat (Falade et al. 2017). Hence, valorizing lignocellulose waste is a welcome call for a proper waste management strategy.
Despite the commonly used chemical pre-treatment strategies to delignify lignocellulose biomass, there is a growing interest in biological delignification. In addition to the high capability of microorganisms to effectively degrade lignin, biological hydrolysis of lignin is also saddled with limited drawbacks (Datta et al. 2017). Unlike the chemical approach associated with high energy requirements and environmental threats, the biological hydrolysis of lignin is cost-effective, ecofriendly, and reduces environmental pollutants (Kumar and Chandra 2020).
The high capability of microorganisms to effectively degrade lignin is due to their ability to produce ligninmodifying enzymes (LMEs). Among the LMEs (laccase, manganese peroxidase, versatile peroxidase, etc.), lignin peroxidase is the most effective (Al-Dhabaan 2019). Lignin peroxidase (LiP), a heme peroxidase, utilizes hydrogen peroxide to oxidize a wide variety of aromatic model compounds (of a phenolic and non-phenolic nature) of lignin into mono-aromatic structures (Janusz et al. 2017). LiP thus could play a pivotal role in treating industrial waste and degradation of xenobiotics such as dyes, organophosphorus, and polycyclic aromatic hydrocarbons (Falade et al. 2017). Such versatile industrial applications of LiP increase the need to search for LiP-producing microorganisms.
Although white rot basidiomycetes have intensively been studied as a source of potent ligninolytic extracellular oxidative enzymes, the ligninolytic use of fungus is limited (Su et al. 2018). Bacteria candidature as ligninolytic agents is currently gaining attention in research. It is due to lesser ethical issues regarding bacteria use, environmental adaptation, spontaneous growth, and the ease of genetic manipulation of bacteria (Tian 2016). Hence, there is a growing interest in exploring bacteria as potential ligninolytic agents. In searching for bacteria that can produce lignin-modifying enzymes (LMEs), bacterial species classified into actinomycetes, α-Proteobacteria, and γ-Proteobacteria have been identified (Wang et al. 2016). Such bacteria include Streptomyces viridosporus, and strains of Nocardia and Pseudomonas (Zimmermann 1990; Bugg et al. 2011).
The Richards Bay environment, a central industrial hub in the Kwa-Zulu Natal province of South Africa, is characterized by loops of hemp of lignin-based wastes and untreated industrial wastewater, posing environmental hazards (Okello et al. 2022). Such an environment might serve as a reservoir for bacterial genome pool to exploit novel bacteria strains with excellent lignin-modifying enzymes. Therefore, this study intended to isolate and characterize potential ligninolytic bacterial strains with lignin peroxidase production capacity from compost samples: sawdust, decaying soil, and cow dung from the Richards Bay environment.
Unless otherwise stated, the chemicals and reagents used in this study were of analytical grades. Nutrient both, nutrient agar, and salts were purchased from Merck (Germany). The kraft lignin, Congo Red, ferric chloride, hexahydrate, sodium azide, pyrogallol, veratryl alcohol, guaiacol alcohol, Malachite green, Remazol Brilliant Blue, Luria broth base, pyrogallol, hydrogen peroxide, disodium salts, and glycerol were supplied by Sigma Aldrich (USA). Bacteriological agar was bought from Lasec (South Africa).
The compost samples (saw-dust, soil, and cow dung) were collected at locations within Richards Bay, South Africa, during summer in March 2021. The soil samples were collected along the uMlalazi river and eSkhaleni forest with coordinates of LA: 28.936164 and LO: 31.763771, and LA: –29.993591 and LO: 30.877430, respectively. Decomposed saw-dust sample was collected from Mtunizini’s Furniture Company (LA: –28.958938 and LO: 31.758050). Cow dung was collected at Farmhouse, University of Zululand (LA: 28.7576 and LO: 32.0497). The samples were kept in sterilized plastic containers and transported to the laboratory, where they were used within 24 hr. The samples (10 g) were mixed with sterile water (90 ml) and centrifuged (30 minutes; 170 rpm). Then, the suspension was diluted to 10–2, 10–3, 10–4, 10–5, and 10–6, respectively. Afterward, the different dilutions (0.1 ml) were added to the prepared medium and daub uniformly. The ethical clearance (UZREC 171110–030 PGM 2018/575) was obtained from the University of Zululand Research Ethics Committee (UZREC) for the use of bacteria for research purposes.
Ligninolytic bacteria were isolated using the enriched media (Minimal Salts Medium Containing Lignin; MSM-L) and the method described by Sasikumar et al. (2014). Briefly, 1 g/l kraft lignin as the sole carbon source was added to the minimal salts medium (MSM), composed of 4.55 g/l K2HPO4, 0.53 g/l KH2PO4, 0.5 g/l MgSO4, 5 g/l NH4NO3, and 0.1 g/l yeast extract. The reagents were vigorously mixed and autoclaved at 121°C for 15 minutes. Enrichment culture was prepared by adding each sample (5 g) to a 250 ml Erlenmeyer flask containing MSM-L broth (95 ml). The culture was then incubated (30°C; 140 rpm) in an orbital shaker (Infors HT Ecotron; Infors HT, Switzerland) for 168 hr. The enriched culture (1 ml) was transferred and mixed with 9 ml of sterilized NaCl (0.9%) solution at room temperature. Serial dilutions were made using 1 ml of the liquid solution, and 100 μl of the serially diluted solutions were poured and spread on MSM-L agar plates containing nystatin (50 mg/l) to prevent fungal development. The plates were then incubated at 30°C for 168 hours, until ligninolytic bacteria became visible. The cultures were then purified using the streaking technique to obtain single colonies, and these were kept in glycerol (20%) at –80°C for later use.
The method described by Taylor et al. (2012) was adopted to evaluate the bacterial ligninolytic potential using lignin model compounds (guaiacol and veratryl alcohols). The isolated bacterial suspension (5 μl; OD620 ≈ 1.0) was inoculated into different agar plates consisting of KH2PO4 (0.53 g/l), NH4NO3 (5 g/l), MgSO4 (0.5 g/l), K2HPO4 (4.55 g/l), 0.1 g/l of yeast extract, veratryl or guaiacol alcohol (0.1% v/v), and 15 g/l agar. The plates were then incubated for 168 hours at 30°C, after which the plates were flooded with Gram iodine solution to determine degradation zones.
The isolates with ligninolytic activity were evaluated for their ability to decolorize certain dyes, viz. Remazol Brilliant Blue R (RBBR), Congo Red (CR), Methylene Blue (MB), and Azure B (AZ), using the method of Bandounas et al. (2011). Briefly, 5 μl of the overnight culture (18 hours) was aseptically inoculated onto dye agar plates made up of KH2PO4 (0.53 g/l), NH4NO3 (5 g/l), MgSO4 (0.5 g/l), K2HPO4 (4.55 g/l), 0.1 g/l yeast extract, 40 mM of glycerol, dye (100 mg/l AZ, 100 mg/l MB, 50 mg/l CR or 100 mg/l RBBR), and 15 g/l of agar. The plates were then incubated at 30°C for 168 hours and observed regularly for growth and formation of zones of decolorization.
All positive isolates from the dye decolorization assay were evaluated qualitatively for peroxidase activity (Falade et al. 2017). The isolates were separately grown on nutrient agar plates and incubated at 30°C for 48 hours. Subsequently, hydrogen peroxide (0.4%; 30 μl) and pyrogallol (1%) were mixed with the grown colonies on the plates. The colonies with the yellow-brown color indicated a positive result.
Ligninolytic bacteria were also tested for laccase activity using the method described by Kameshwar and Qin (2017). The isolated ligninolytic bacteria strains were activated overnight and inoculated on MSL agar plates supplemented with 0.1% w/v ABTS, 1.6% w/v Agar, and 20% w/v glucose. The plates were incubated in the dark for 10 days at 30°C. The development of green color on colonies indicated positive results.
The bacterial isolates with ligninolytic activity were subjected to standard biochemical tests, including catalase, oxidase, starch hydrolysis, urease, indole, and Simmon’s citrate test (Badr 2018; Al-Dhabaan 2019).
The 16S rRNA genes of the bacterial isolates with ligninolytic activities were analyzed. The 16S rDNA was amplified, and sequence analyzed through BLAST (NCBI). The bacteria genomic DNA was extracted from the colonies using Quick-DNA™ Fugal/Bacterial Miniprep Kit (Zymo Research, USA). The targeted region of 16S rDNA was tagged with OneTaq ® Quick-Load® 2X Master Mix (New England Biolabs® Inc., USA) along with 16S–1492R (5’-CGGTTACCTTGTTAC-GACTT-3’) and 16S-27F (5’-AGAGTTTGATCMTG-GCTCAG-3’) primers, respectively. The products were then run through a gel and extracted with Zymoclean™ Gel DNA Recovery Kit (Zymo Research, USA). The DNA fragments extracted were further sequenced with Nimagen (The Netherlands), BrilliantDye™ Terminator v3.1 Cycle Sequencing Kit (BRD3-1000) and purified with Zymo Research (USA), ZR-96 DNA Sequencing Clean-up Kit™. The pure fragments were analyzed using the ABI 3500xL Genetic Analyzer (Applied Biosystems™, ThermoFisher Scientific Inc., USA). The file generated by the ABI analyzer was further analyzed with GLC Bio Main Workbench v7.6 with the aid of BLAST search (NCBI). The phylogenetic tree was constructed by neighbor joining methods, and 99% similarity with existing isolates in the database was considered (Xiong et al. 2020).
The bacterial isolates with ligninolytic enzyme activity were evaluated for lignin biodegradation capability following the method described by Dicko et al. (2020), with slight modification. The experiment was carried out on a medium containing kraft lignin (3 g/l) as a carbon source. The purified colonies were inoculated on nutrient broth and incubated for 48 hours at 30°C to activate the colonies. Afterward, bacterial suspensions (2%) in saline were cultured in conical flasks (250 ml) containing MSM-L broth fortified with glucose (1%) until the OD620 was 1 nm. It was incubated for 6 days at 30°C in a shaker (120 rpm). Aliquots (2 ml) were taken daily from each flask, bacterial growth (A620), and pH were measured accordingly. The collected aliquots were centrifuged (15,000 rpm; 5 minutes), and their supernatants (250 μl) were mixed with phosphate buffer (750 μl). The absorbance of color reduction and lignin degradation was read at A465 and A280, respectively, using a spectrophotometer. Graphs of bacterial growth, lignin degradation, pH vs. time, and bacterial growth were plotted.
The lignin peroxidase activity of the isolates was determined by monitoring the rate of H2O2-dependent oxidation of pyrogallol to purpurogallin (Falade et al. 2017). Pyrogallol (310 μl of 5% w/v) was pipetted into an Eppendorf tube with potassium phosphate buffer (100 mM; pH 6) and mixed. Afterward, the cultured supernatant (25 μl) was added to the solution and mixed by the vortex. The mixture without the enzymes served as blank. The reaction was initiated by the addition of 15 μl of 0.5% v/v of H2O2. The direct increase in absorbance was read at 420 nm every 30 seconds for 2–3 minutes at room temperature, using BioTeK Synergy™ HT 96-well Microtitre Plate Reader (BioTeK® Instruments Inc., USA). The enzymatic activity was subsequently calculated using the following equation:
Unless otherwise stated, all the experiments were triplicated, and the results were expressed as mean ± standard deviation (SD). The data for decolorization assay was analyzed by one-way ANOVA and comparison analysis by Dunn’s post hoc using GraphPad Prism software (version 6).
Table SI depicts the potential ligninolytic isolates from the compost samples. Twenty-six (26) bacteria strains were successfully isolated from cow dung, soil, and sawdust samples.
The results indicating the ability of the isolates to degrade and utilize guaiacol and veratryl alcohols are presented in Table I. After over 168 hours, the isolated microbes could grow on lignin monomers – veratryl and guaiacol alcohol – as a carbon source. While fifteen (58%) of the total isolates were able to degrade both guaiacol and veratryl alcohols, only seven (27%) of the isolates degraded veratryl alcohol only. Four (15%) of the isolates could not grow on any of the alcohols.
Degradation of lignin model compounds; guaiacol and veratryl alcohols by the bacterial isolates from the compost samples.
| S/N | Bacterial code | Isolation source | Guaiacol alcohol | Veratryl alcohol |
|---|---|---|---|---|
| 1 | Old A | Old cow dung | ++ | ++ |
| 2 | *Old A | Old cow dung | ++ | ++ |
| 3 | OOld A | Old cow dung | ++ | +++ |
| 4 | Old A1 | Old cow dung | + | + |
| 5 | *Old A2 | Old cow dung | + | ++ |
| 6 | Old A2 | Old cow dung | +++ | +++ |
| 7 | /*New A | Fresh cow dung | – | ++ |
| 8 | New A | Fresh cow dung | +++ | +++ |
| 9 | *New A | Fresh cow dung | – | ++ |
| 10 | New B | Fresh cow dung | ++ | ++ |
| 11 | ONew B | Fresh cow dung | – | – |
| 12 | *New B | Fresh cow dung | – | – |
| 13 | Saw A | Sawdust | – | – |
| 14 | Saw A1 | Sawdust | – | + |
| 15 | Saw A2 | Sawdust | +++ | +++ |
| 16 | Saw B | Sawdust | – | – |
| 17 | *Saw B | Sawdust | – | +++ |
| 18 | Saw B1 | Sawdust | – | ++ |
| 19 | Soil A | Soil | – | + |
| 20 | *Soil A | Soil | – | +++ |
| 21 | Soil A1 | Soil | + | ++ |
| 22 | *Soil B | Soil | + | +++ |
| 23 | Soil B | Soil | ++ | +++ |
| 24 | Soil B1 | Soil | +++ | +++ |
| 25 | Soil 2 | Soil | + | ++ |
| 26 | Soil 7 | Soil | + | ++ |
– – negative
+ – weak
++ – moderate
+++ – strongly positive
The ligninolytic potential of 15 isolates that degraded both alcohols (Table I) was further confirmed with dye decolorization assay, and the results are shown in Table II. It is apparent that all 15 isolates successfully degraded Azure B, although at various degrees of efficiency. Meanwhile, 12 isolates degraded Remazol Brilliant Blue R only, and 13 degraded Congo Red only. Only 10 isolates effectively degraded all the tested dyes out of which isolate Old A2 (obtained from old cow dung) showed the highest ligninolytic activity when compared to the other isolates.
Decolourization of synthetic lignin-like dyes (halo zone measured in mm) by the selected ligninolytic bacterial isolates.
| S/N | Bacterial code | RBBR (mm) | AZ (mm) | CR (mm) |
|---|---|---|---|---|
| 1 | Old A | 15.0 ± 0.10c | 35.0 ± 0.04b | 0.00 ± 0.00f |
| 2 | *Old A | 26.0 ± 0.00b | 40.0 ± 0.01d | 42.0 ± 0.01a |
| 3 | OOld A | 24.0 ± 0.06b | 30.0 ± 0.00c | 31.0 ± 0.10c |
| 4 | Old A1 | 19.0 ± 0.00d | 29.0 ± 0.05c | 0.00 ± 0.00f |
| 5 | *Old A2 | 0.00 ± 0.00f | 34.0 ± 0.04e | 10.0 ± 0.20d |
| 6 | Old A2 | 35.0 ± 0.10a | 42.0 ± 0.11d | 40.0 ± 0.08a |
| 7 | New A | 8.00 ± 0.50e | 40.0 ± 0.00d | 20.0 ± 0.01e |
| 8 | New B | 30.0 ± 0.00h | 50.0 ± 0.01a | 26.0 ± 0.02b |
| 9 | Saw A2 | 15.0 ± 0.04c | 35.0 ± 0.02b | 12.0 ± 0.50d |
| 10 | Soil A1 | 0.00 ± 0.00f | 8.00 ± 0.01g | 24.0 ± 0.10b |
| 11 | *Soil B | 0.00 ± 0.00f | 25.0 ± 0.02f | 19.0 ± 0.00e |
| 12 | Soil B | 34.0 ± 0.00a | 50.0 ± 0.02a | 19.0 ± 0.10e |
| 13 | Soil B1 | 10.0 ± 0.06e | 37.0 ± 0.01b | 40.0 ± 0.03a |
| 14 | Soil 2 | 49.0 ± 0.00h | 25.0 ± 0.08f | 49.0 ± 0.01h |
| 15 | Soil 7 | 35.0 ± 0.02a | 16.0 ± 0.04h | 35.0 ± 0.05i |
The superscripts represented significant different (p < 0.05).
RBBR – Remazol Brilliant Blue R, AZ – Azure B, CR – Congo Red
All 10 isolates that displayed positive results on the tested dyes were qualitatively evaluated for lignin peroxidase and laccase activities. The results revealed that all the isolates possess a prolific capability to produce lignin peroxidase (Table III). However, all the isolates showed to be non-laccase producers.
Lignin peroxidase and Laccase producing potential of the isolated bacteria.
| S/N | Bacterial code | Lignin peroxidase activity | Laccase activity |
|---|---|---|---|
| 1 | *Old A | +++ | – |
| 2 | OOld A | +++ | – |
| 3 | Old A2 | +++ | – |
| 4 | New A | +++ | – |
| 5 | New B | +++ | – |
| 6 | Saw A2 | +++ | – |
| 7 | Soil B | +++ | – |
| 8 | Soil B1 | +++ | – |
| 9 | Soil 2 | ++ | – |
| 10 | Soil 7 | ++ | – |
– – negative
+ – weak
++ – moderate
+++ – strongly positive
The results of the biochemical characteristics of the targeted 10 isolates with ligninolytic activity are presented in Table IV. The isolates showed positive catalase, starch hydrolysis, and indole tests. However, they could not utilize urease or citrate as a sole carbon source.
Characteristics of ligninolytic bacteria from cow dung, sawdust, and soil samples.
| Bacterial code | Catalase test | Oxidase test | Starch hydrolysis | Urease test | Indole test | Citrate test |
|---|---|---|---|---|---|---|
| *Old A | + | – | + | – | + | – |
| OOld A | + | + | + | – | + | – |
| Old A2 | + | + | + | – | + | – |
| New A | + | + | + | – | + | – |
| New B | + | + | + | – | + | – |
| Saw A2 | + | – | + | – | + | – |
| Soil B | + | – | + | – | + | – |
| Soil B1 | + | – | + | – | + | – |
| Soil 2 | + | + | + | – | + | – |
| Soil 7 | + | – | + | – | + | – |
– – negative
+ – positive
16S rDNA gene sequencing fingerprinting was performed the 10 isolates identified: seven P. aeruginosa isolates, two Enterobacter species (Enterobacter kobei and Enterobacter xiangfangensis) and one Escherichia coli isolate (Table V).
Identification of the isolated bacterial cultures by 16S rDNA.
| Bacterial name | Accession number (16S rRNA gene sequence) | Bacterial code |
|---|---|---|
| Pseudomonas aeruginosa | KR136350.1 | Soil B |
| Pseudomonas aeruginosa | KT799669.1 | *Old A |
| Pseudomonas aeruginosa | MF144461.1 | OOld A |
| Pseudomonas aeruginosa | KR136350.1 | New B |
| Pseudomonas aeruginosa | KR136350.1 | New A |
| Pseudomonas aeruginosa | CP031449.2 | Saw A2 |
| Pseudomonas aeruginosa | KT799669.1 | Soil B1 |
| Escherichia coli | LR025096.1 | Old A2 |
| Enterobacter kobei | CP032897.1 | Soil 7 |
| Enterobacter xiangfangensis | MH304301.1 | Soil 2 |
The results demonstrating the capability of the bacterial isolates to degrade and utilize kraft lignin are presented in Fig. 1. All the isolates showed appreciable levels of growth; it is noteworthy that the isolates’ growth corresponded with their lignin degradation capability and the changes in the pH levels. E. coli and P. aeruginosa showed the most appreciable lignin degradation ability compared to the other isolates.
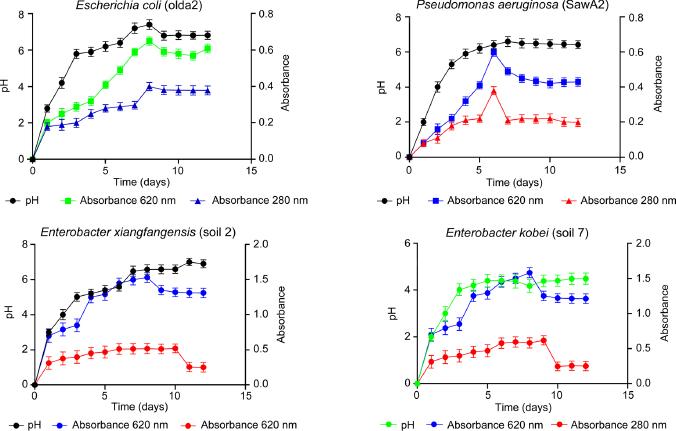
Graphs of lignin degradation capability, pH variation, and growth of E. coli, P. aeruginosa, E. kobei, and E. xiangfangensis.
The qualitative lignin peroxidase activity in the recovered supernatant of the bacteria cultures is presented in Table VI. While all the isolates produced the enzyme, only P. aeruginosa (CP031449.2) and E. coli (LR0250096.1) could produce an appreciable quantity of lignin peroxidase (1.99 × 10–1 U/l). In addition, the two bacteria also exhibited reasonable exponential enzyme activities, with optimum activity occurring between 5 to 9 days of growth (Fig. 2).
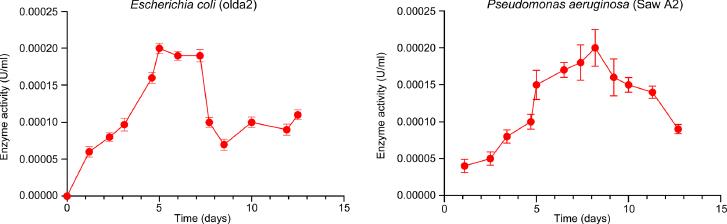
Enzyme activity (the recovered supernatant) of E. coli (LR0250096), and P. aeruginosa (CP031449.2) on the lignin enriched medium.
Quantitative analysis for the lignin peroxidase activity of the bacteria isolates.
| S/N | Bacterial strains | Lignin peroxidase activity (U/l) |
|---|---|---|
| 1 | Pseudomonas aeruginosa (MF144461.1) | 1.37 × 10–2a |
| 2 | Escherichia coli (LR025096.1) | 1.99 × 10–1b |
| 3 | Pseudomonas aeruginosa (KR136350.1) | 8.70 × 10–2c |
| 4 | Pseudomonas aeruginosa (CP031449.2) | 1.99 × 10–1b |
| 5 | Enterobacter xiangfangensis (MH304301.1) | 1.09 × 10–2d |
| 6 | Enterobacter kobei (CP032897.1) | 8.55 × 10–2c |
The superscripts represented significant different (p < 0.05).
This study isolated twenty-six potential ligninolytic bacteria from samples collected in the Richards Bay environment. Most of the isolates were identified (Table SI) as P. aeruginosa (88%), Enterobacter spp. (8%), and E. coli (4%). These ligninolytic bacteria were all found to be γ-Proteobacteria. The phylogenetic tree, showing the relationship between the P. aeruginosa and the Enterobacter hormaechei in this study, and other ligninolytic bacteria is presented in Fig. 3. The identified isolates all showed 99% similarity to the previously isolated strains (Xiong et al. 2020).
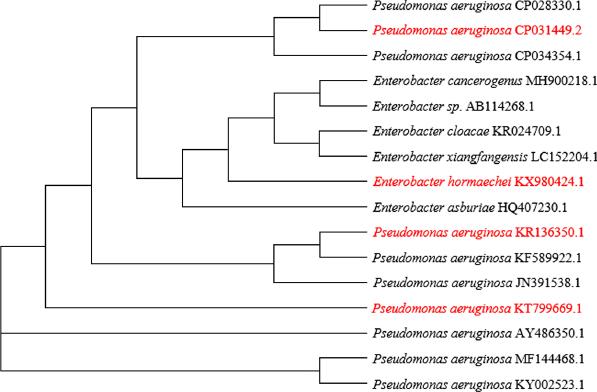
The phylogenetic tree of the bacteria from this study (red) in relation to some other ligninolytic bacteria in the NCBI database.
According to Lopez et al. (2006), using lignin as a carbon source for the selection of ligninolytic microorganisms is regarded as one of the most appropriate methods worldwide. However, the utilization of lignin model compounds such as veratryl and guaiacol alcohols, and the decolorization of industrial dyes as exhibited by the isolates (Table I and II), is the evidence that the colonies have the potential to utilize and degrade lignin (Husain 2006; Bandounas et al. 2011; Falade et al. 2017).
However, growth on polymeric lignin or lignin monomers is not necessarily a good measure of the ligninolytic potential. The ligninolytic potential, independent from lignin utilization, was studied with the decolorization of synthetic lignin-like dyes (Falade et al. 2017). The selected organisms were able to decolorize Azure B. Although most LMEs can decolorize various dyes, previous studies (Falade et al. 2017) have revealed that Azure B decolorization is associated explicitly with high redox potential agents like lignin peroxidases (LiP). Other LMEs, such as laccase and manganese peroxidase-producing bacteria, cannot decolorize Azure B (AZ). Since fifteen of the isolated ligninolytic bacteria degraded AZ (Table II), it is apparent that these bacterial isolates were potential LiP producers. However, out of the 15 isolates, 10 isolates were able to decolorize Azure B only. These pure colonies gave a positive pyrogallol test for peroxidase, and the supernatant of cultures quantitatively revealed the activity of peroxidase (Arora and Gill 2011). Therefore, they were selected as potential lignin peroxidase producers, and this was confirmed by the production of a crude enzyme complex that contained a peroxidase, most likely, a lignin peroxidase. They were not laccase producers (Table III). All the selected isolates also showed high catalase and oxidase activities and were capable of hydrolyzing starch (Table IV).
The lignin degradation capabilities of these bacterial strains were further examined, and the evidence for ligninolytic activity was based on) the bacterial growth on lignin enriched medium and the lignin degradation. Most of the tested bacterial strains were able to degrade lignin. The isolated organisms were cultured on lignin (with 1% glucose), and after a period of cultivation, phenolic (or derived lignin monomers) appeared and increased with growth in the culture. We believe that the organisms produced these phenolic compounds in the culture as a result of lignin (a polyphenolic complex) being degraded. We also agree that the lignin degradation should be monitored as directly as possible. However, synthetic dyes may mimic lignin (Sana et al. 2017). In this present study, the lignin degradation was prominent with E. coli LR0250096.1 and P. aeruginosa CP031449.2, as their growth increased with the degradation of lignin (Fig. 1). These results are consistent with literature reports on the ligninolytic activity of Pseudomonas sp. and E. coli species (Bandounas et al. 2011; Sana et al. 2017).
Interestingly, of 26 isolates obtained, ten were identified as true lignin peroxidase producers with E. coli LR0250096.1, and P. aeruginosa CP031449.2 considered the best lignin peroxidase producers, exhibiting activity of about 1.99 × 10–1 U/l (Table VI, Fig. 2). Other bacteria strains such as Klebsiella pneumonia, Bacillus species, Citrobacter species, Serratia marcescens, and Pseudomanos spp. have also been reported to have LMEs-producing potential (Zainith et al. 2019; Kumar and Chandra 2020).
This study attempted to isolate and characterize lignin-degrading bacteria capable of producing lignin peroxidase. The isolated Pseudomonas and Enterobacter species and E. coli were identified as true lignin peroxidase producers. The highest ligninolytic activity exhibited by P. aeruginosa (CP031449.2) and E. coli (LR025096.1) suggests that these isolates could be exploited in the industrial treatment of lignin and detoxification of recalcitrant industrial dyes that invariably end up in the environment. For further studies, enzyme purification, enzyme’s efficiency, and the mechanism of its action will be ascertained.