
The Mediterranean Sea is one of the world’s most affected marine regions by biological invasions. Over 950 alien species have been documented, with the majority originating from the Red Sea via the Suez Canal (Galil et al., 2015; Zenetos et al., 2012). This process, known as Lessepsian migration, has profoundly altered native biodiversity and ecosystem functioning in the eastern Mediterranean (Katsanevakis et al., 2014; Streftaris et al., 2005). Among the most successful invaders are penaeid shrimps, which have established dense populations along the Levantine coasts (Çinar et al., 2011).
The kuruma shrimp, M. japonicus (Spence Bate, 1888), is native to the Indo-West Pacific, ranging from East Africa to Japan and Australia (Hayashi, 1996). It was first recorded in the Mediterranean off the Egyptian coast in 1927 and has since expanded throughout the Levantine basin, including Turkish, Israeli and Cypriot waters (Holthuis, 1958; Özvarol & Kazancı, 2017). The species is highly prized for its large size and meat quality, supporting both artisanal fisheries and intensive aquaculture in its native range (Hamasaki & Kitada, 2006).
Recent molecular studies have revealed that M. japonicus is a species complex comprising two cryptic forms (Forms I and II). Form I is restricted to the East China Sea (including Japan), while Form II is widely distributed in the South China Sea, Australia, the Red Sea, the Mediterranean and the western Indian Ocean (Francis et al., 2021; Tsoi et al., 2005). The native stock along the Indian coastline has been identified as Form II (Vinay et al., 2019). A high-quality chromosome-level genome assembly (1.64 Gb) has recently been published, providing valuable genomic resources for understanding the species’ biology and supporting selective breeding programs (Wei et al., 2025).
In Turkish waters, M. japonicus has become a regular component of coastal fisheries catches, particularly in the Gulf of Antalya, where it inhabits sandy and sandy-mud bottoms at depths of 10–100 m (Özvarol & Kazancı, 2017). Despite its increasing economic importance, detailed biological information on Mediterranean populations remains limited. Most existing studies focus on the species’ biology in its native Indo-Pacific range (Minagawa et al., 2000; Ohtomi et al., 2003) or on other penaeids (Tom & Lewinsohn, 1983). This knowledge gap hinders the development of effective management strategies for this emerging fishery resource.
Reproductive traits such as sex ratio, spawning season and size at first sexual maturity (TL50) are key determinants of population productivity and are influenced by environmental factors, including temperature and salinity (Dall et al., 1991; Hartnoll, 2001; King, 1995). Characterizing these parameters provides essential baseline data for fisheries management, including the establishment of minimum landing sizes and seasonal fishing closures to protect spawning stock biomass (Hamasaki & Kitada, 2006). Given the scarcity of biological data for Mediterranean populations and noting that recent studies on penaeid shrimps in the region have largely been limited to length–weight relationships (Özvarol & Kazancı, 2017; Sellars et al., 2009), this study provides an integrated assessment of reproductive biology, population structure and growth of M. japonicus in the Gulf of Antalya, combining GSI analysis, maturity staging, length–weight relationships (LWR) and size at maturity estimation within a single-year monthly sampling framework, thereby expanding upon previous regional studies.
Sampling was conducted monthly from January to December 2018 in the Gulf of Antalya, along the coastal area between Side Lighthouse and Lara (Fig. 1), at depths ranging from 10 m to 50 m.
Map of the study area showing the sampling locations in the Gulf of Antalya (eastern Mediterranean).
Specimens were obtained using shrimp trammel nets during routine fishing operations. The same type of fishing gear was used throughout the study period. All samples were collected at regular monthly intervals and transported to the laboratory for analysis. Due to the use of trammel nets, the size composition of the samples may have been influenced by gear selectivity, potentially leading to undersampling of smaller individuals. This limitation is considered in the interpretation of size at maturity (TL50) estimates (see Discussion). Samples were collected monthly from the study area between January and December 2018 using shrimp trammel nets. All individuals were sexed and measured following standard procedures. The number of male and female individuals collected in each month is provided in Supplementary Table 1.
Seawater temperature, salinity and pH were measured in situ at each sampling site using a calibrated multiparameter probe (SKU 599502-00; YSI, Xylem Inc., Yellow Springs, OH, USA). Sediment composition was analyzed to characterize the general substrate type of the sampling area. Environmental measurements were recorded together with the sampling date and location.
Total length (TL) was measured from the tip of the rostrum to the tip of the telson using digital calipers (±0.1 cm). Body weight (W) was recorded to the nearest 0.01 g using an analytical balance. Sex was determined macroscopically based on the presence of the thelycum in females and the petasma in males (Pérez Farfante & Kensley, 1997).
Fulton’s condition factor (K) was calculated separately for males and females using the equation
where W represents body weight (g) and TL represents total length (cm) (Froese, 2006).
Homogeneity of variances was checked with Levene’s test; when assumptions were violated, the Kruskal–Wallis test was applied. Differences between sexes were assessed with independent-samples t-tests (for normally distributed data) or Mann–Whitney U-tests (for non-normal data).
Ovaries were removed and weighed to the nearest 0.001 g using an analytical balance. The gonadosomatic index (GSI) was calculated for females using the formula (King, 1995).
Ovarian development was classified into five stages based on macroscopic examination, following criteria adapted from Minagawa et al. (2000) and Yano (1988):
Stage I (Immature): Thread-like, transparent ovaries, no visible oocytes.
Stage II (Developing): Slightly enlarged, creamy white to pale yellow.
Stage III (Late Developing): Enlarged, pale green to yellow, occupying much of the cephalothorax.
Stage IV (Mature): Large, turgid, dark green to olive, with visible oocytes.
Stage V (Spent): Flaccid, reduced, reddish or translucent.
The LWR was estimated using the power function
The size at which 50% of females reached sexual maturity (TL50) was estimated by fitting a logistic regression model to the proportion of mature females (stages III and IV) in each 1-cm length class (Somerton, 1980).
All statistical analyses were performed using IBM SPSS Statistics version 25 (IBM Corp., Armonk, NY, USA). The sex ratio was tested against a 1:1 null hypothesis using a Chi-square (χ2) test. Differences in mean sizes between sexes were compared using an independent-samples t-test. Monthly GSI values were compared using one-way analysis of variance (ANOVA). Significance was set at α = 0.05.
Seawater temperature ranged from 15.5°C (January) to 29.3°C (August), with a yearly average of 21.8°C. Salinity values ranged from 39.3%o (July) to 40.2%o (September). Surface pH varied from 8.2 (March) to 9.0 (April). Sediment composition at sampling sites consisted primarily of muddy and muddy-sandy substrates.
Monthly mean GSI correlated positively with seawater temperature (Pearson’s r = 0.85, p < 0.01). The highest GSI values were recorded in July and August. No significant correlation was found between GSI and salinity or pH (p > 0.05). Given the monthly sampling resolution (1 year), the analysis was limited to Pearson correlations, as more advanced modeling would be prone to overfitting.
A total of 516 specimens were examined, comprising 267 females (51.7%) and 249 males (48.3%). The overall sex ratio (1.07:1) did not differ significantly from the expected 1:1 ratio (χ2 = 0.63, p > 0.05). Females exhibited significantly larger TLs than males (p < 0.001). Female TL ranged from 12.60 cm to 21.07 cm (16.9 ± 2.16 cm). Male TL ranged from 12.84 cm to 17.9 cm (14.9 ± 1.23 cm). The largest female weighed 78.16 g. The largest male weighed 32.6 g.
LWRs were calculated separately for females, males and the pooled data. The estimated parameters are summarized in Table 1.
LWR parameters for M. japonicus in the Gulf of Antalya.
| Group | n | A | B | 95% CI of b | R2 |
|---|---|---|---|---|---|
| Females | 267 | 0.0038 | 3.3146 | 3.09–3.34 | 0.947 |
| Males | 249 | 0.0127 | 2.7352 | 2.61–2.90 | 0.836 |
| Pooled | 516 | 0.0035 | 3.1814 | 3.08–3.30 | 0.924 |
The b-value for females (3.315) was significantly greater than 3 (p<0.05), indicating positive allometric growth.
The b-value for males (2.735) was significantly less than 3 (p<0.05), indicating negative allometric growth.
LWR, length–weight relationship.
Monthly variation in Fulton’s condition factor (K) for males and females is presented in Fig. 2. Fulton’s K values ranged between 0.037 and 0.050 throughout the study period. Mean K values did not differ significantly between sexes (t-test, p > 0.05). However, seasonal variation in K was statistically significant (ANOVA, p < 0.05).
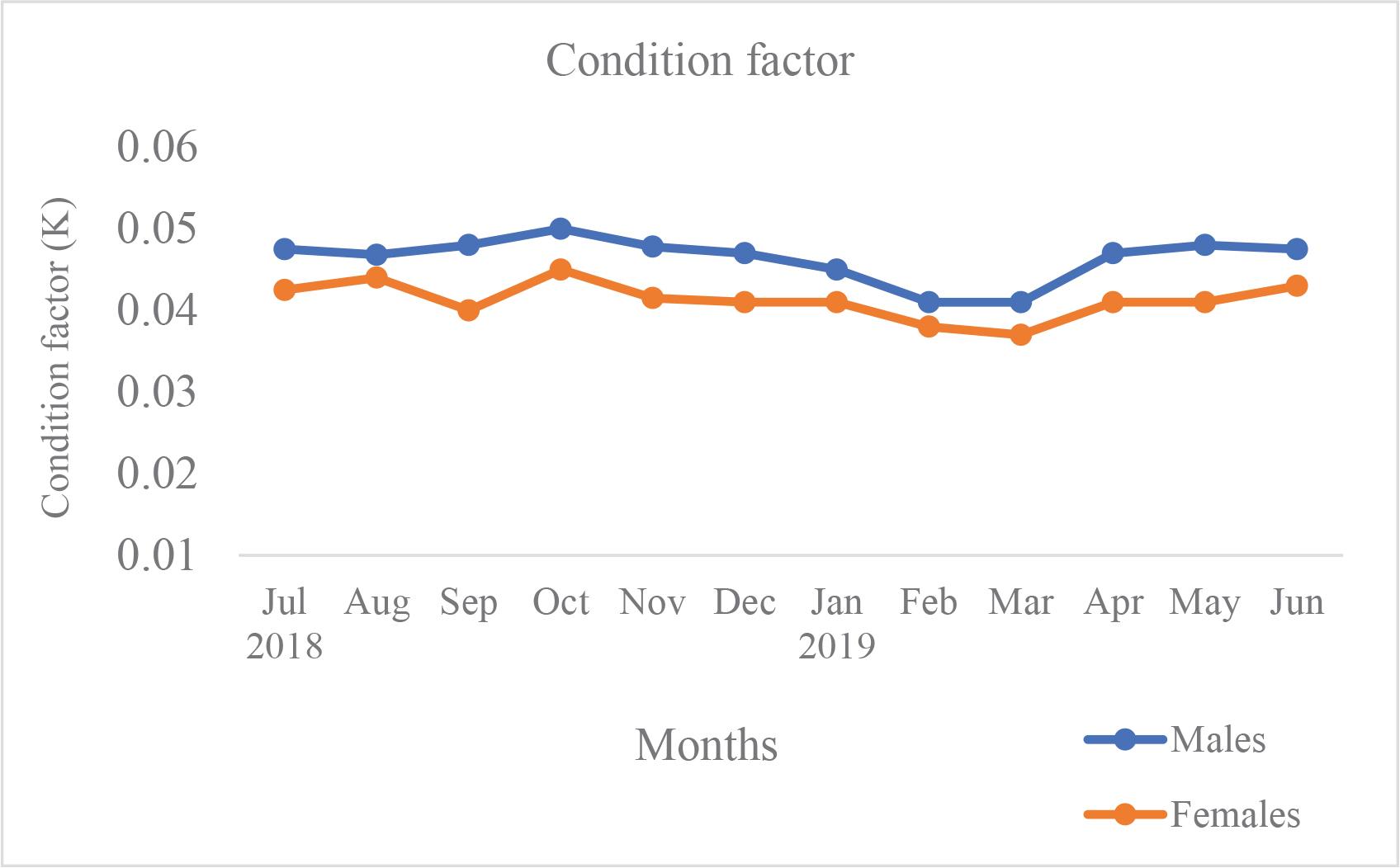
Monthly variation of Fulton’s condition factor (K) in male and female M. japonicus. Values represent mean ± SD.
Monthly GSI values for females are presented in Fig. 3. Values remained below 2.0 from December to February and increased thereafter, exceeding 6.0 in July (6.13) and August (6.06). Monthly differences were statistically significant (ANOVA, p < 0.001), indicating a clear seasonal pattern in reproductive activity.
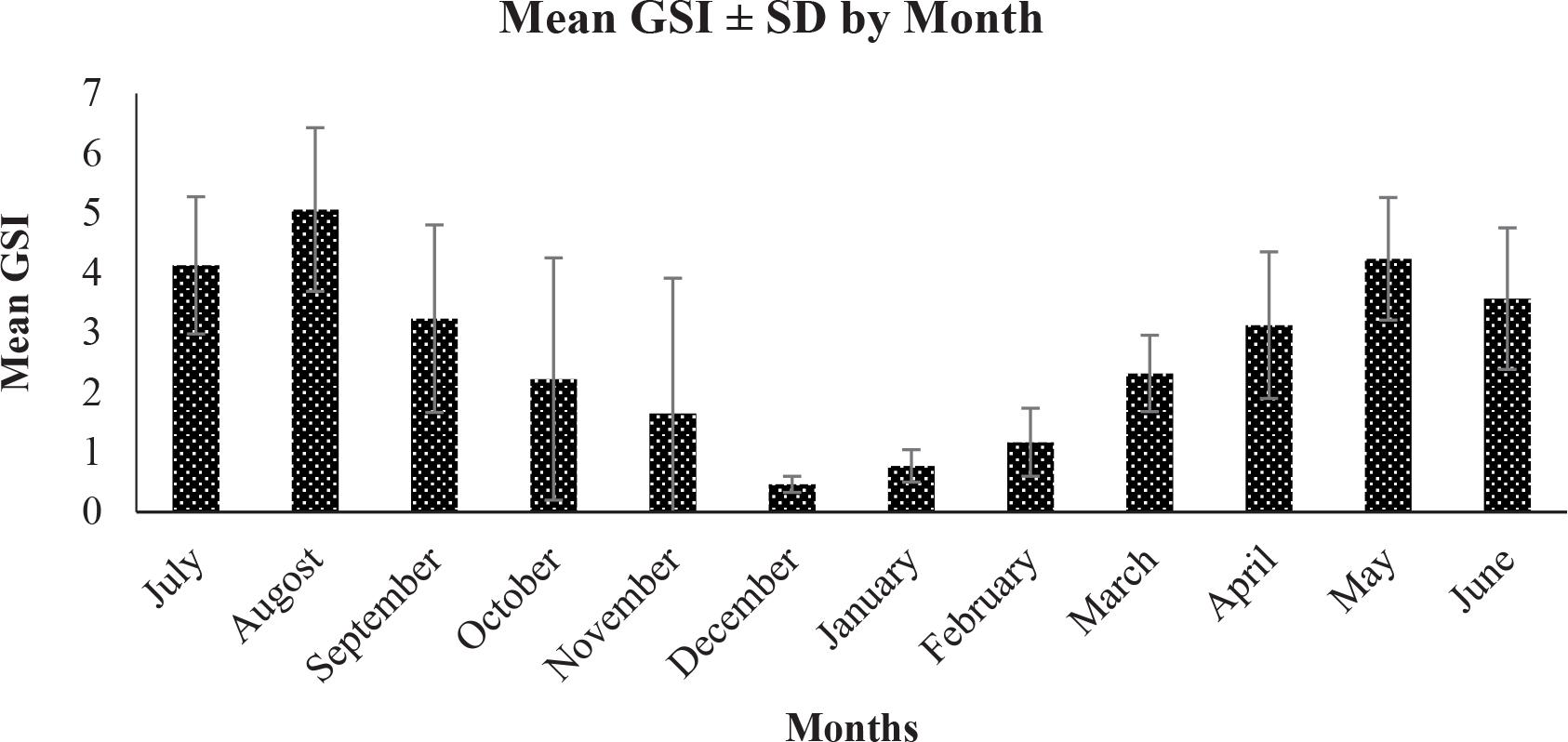
Monthly variation in the GSI of female M. japonicus. GSI, gonadosomatic index.
Monthly variation in the GSI of M. japonicus females and the distribution of gonadal maturity stages are presented in Figs 3 and 4, respectively. Immature females (stage I) were primarily observed between December and February. Mature females (stages III and IV) were present from May to September, while spawning individuals (stage V) appeared from June onward, peaking during July and August. Seasonal variation in GSI values and maturity-stage distributions was analyzed based on monthly sampling. The monthly distribution of sampled individuals is presented in Supplementary Table 1.
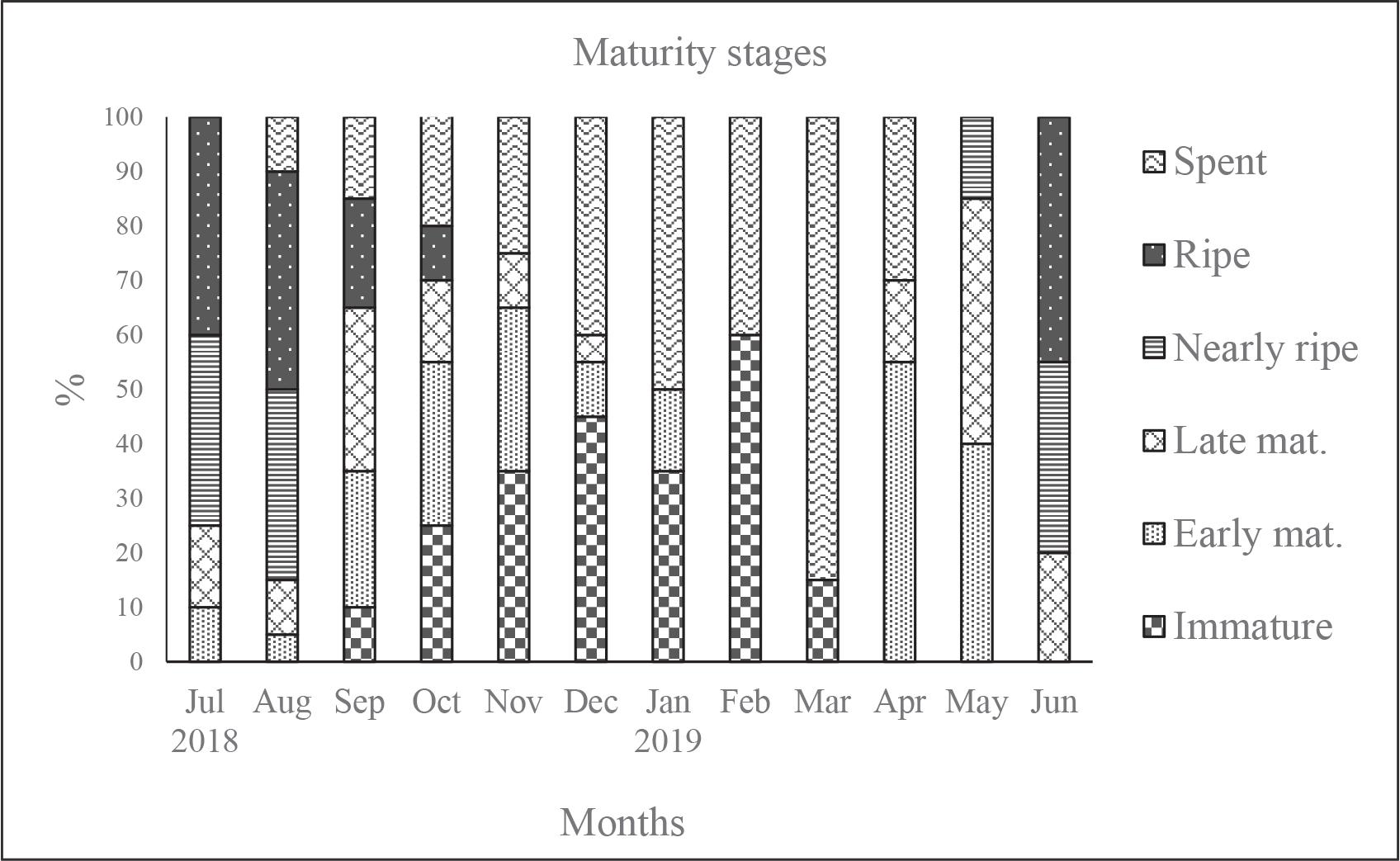
Monthly variation in the percentage distribution of gonadal maturity stages of female M. japonicus in the Gulf of Antalya.
Logistic regression analysis estimated the size at which 50% of females reached sexual maturity (TL50) at 15.3 cm TL (95% CI: 14.9–15.7 cm). The model parameters were statistically significant (slope = 1.204, p < 0.001; intercept = –18.42, p < 0.001), with a Nagelkerke R2 of 0.98 (Fig. 5). The smallest mature female observed measured 13.5 cm TL (stage III), while all females above 16.3 cm TL were mature.
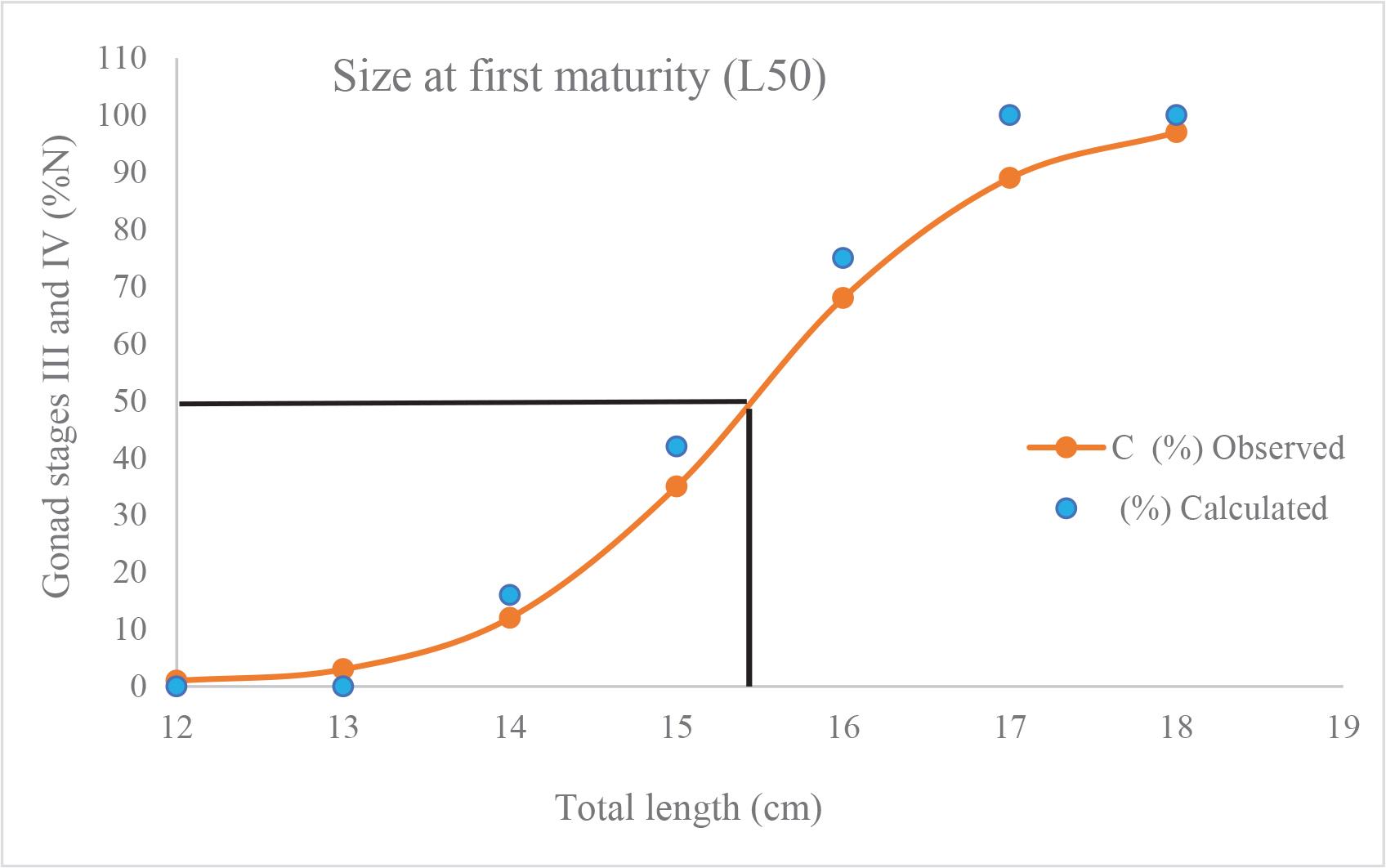
Size at first sexual maturity (TL50) of female M. japonicus.
The present study examines population structure, growth dynamics and reproductive characteristics of M. japonicus in the Gulf of Antalya. Females reached significantly larger sizes than males, a pattern widely recognized in penaeid shrimps and attributed to differential energy allocation, where females invest more heavily in somatic growth to maximize reproductive output (Dall et al., 1991; Hansford & Hewitt, 1994). Comparable size differences have been reported in Japanese populations (Minagawa et al., 2000; Ohtomi et al., 2003) and in other commercially important penaeids, such as Penaeus monodon (Li et al., 2016).
The observed sex ratio (1.07:1) did not deviate significantly from parity, contrasting with several earlier studies that reported female-biased populations in M. japonicus from the southeastern Mediterranean (Tom & Lewinsohn, 1983) and the Gulf of Antalya (Özvarol & Kazancı, 2017). Such variability may arise from differences in sampling gear, seasonal timing or habitat-specific segregation of sexes, which have been documented in penaeids (Dall et al., 1991). The near parity in this study suggests that both sexes are similarly represented in the sampled fishing grounds during the study period.
LWRs further support sex-specific growth patterns. Females exhibited positive allometric growth (b = 3.315), indicating a disproportionate increase in weight relative to length, whereas males showed negative allometry (b = 2.735). This divergence likely reflects physiological differences associated with reproduction. In females, increasing body mass is closely linked to ovarian development and the accumulation of energy reserves necessary for gametogenesis (Teshima, 1998). Conversely, males may allocate relatively less energy to somatic growth, potentially prioritizing reproductive structures or mating activity (Primavera et al., 1998).
The LWR results obtained in this study are highly consistent with those reported by Özvarol & Kazancı (2017) from the same region (Gulf of Antalya), where females also exhibited positive allometry (b = 3.216) and males negative allometry (b = 2.756), confirming the robustness of these sex-specific growth patterns in M. japonicus. However, while Özvarol & Kazancı (2017) focused solely on LWR and size distribution with a limited sample size (n = 108), the present study substantially extends these findings by integrating LWR with comprehensive reproductive parameters (GSI, maturity stages, TL50, spawning season) based on a larger sample (n = 516). Similar sex-dependent growth trajectories have also been documented for M. japonicus in other parts of the northeastern Mediterranean (Kumlu et al., 1999) and across other penaeid species (Primavera et al., 1998).
Several factors may explain the extended reproductive period (April–November) observed in the Gulf of Antalya. First, seawater temperatures above 20°C persist for more than 6 months in the region, providing a prolonged thermal window for gonadal development and spawning. It is well documented that spawning in M. japonicus and other penaeid shrimps begins when sea temperature exceeds 20°C (Dall et al., 1991; IUCN MedMis, 2025; Kumlu et al., 1999). From an ecological perspective, an extended spawning season increases the probability of recruitment success by spreading larval release across variable environmental conditions, potentially conferring a competitive advantage over native species with shorter, more synchronized spawning periods. The eastern Mediterranean is a recognized climate change hotspot, with sea surface temperatures rising approximately 1–1.5°C over the past 3 decades (Albano et al., 2021). Further warming is expected to prolong the reproductive season even more and may cause shifts in TL50 (Tsirintanis et al., 2022).
Fulton’s condition factor (K) ranged between 0.037 and 0.050, with no significant difference between sexes but significant seasonal variation. Condition values showed a slight increase during late summer and autumn, coinciding with the main reproductive period indicated by GSI and maturity stage analyses. Comparable seasonal patterns in condition factor have been reported for other penaeid species, varying in relation to reproductive activity and environmental conditions (Kumlu et al., 1999; Teshima, 1998).
Environmental factors, particularly temperature, played a central role in shaping reproductive dynamics. Seawater temperatures ranged from 15.5°C to 29.3°C and were strongly correlated with GSI (see Results), indicating that warmer months favor gonadal development. This relationship aligns with previous studies showing that temperature strongly influences reproduction in penaeid shrimps, affecting gametogenesis, spawning frequency and larval survival (King, 1995; Teshima, 1998). In Mediterranean populations, peak reproductive activity also coincides with the warmest months, consistent with observations from M. japonicus in the Gulf of Antalya (Kumlu et al., 1999; Tom & Lewinsohn, 1983). Salinity and pH remained within ranges that did not significantly affect gonadal development or body condition, consistent with prior reports indicating that M. japonicus tolerates moderate fluctuations in these parameters (Özvarol & Kazancı, 2017).
A methodological caveat should be noted: trammel nets may undersample smaller individuals, which could theoretically lead to a slight overestimation of TL50. As demonstrated by Brčić et al. (2019) for Mediterranean crustacean fisheries, size-dependent selectivity is a well-documented phenomenon in passive fishing gears. However, the smallest mature female observed (13.5 cm TL) was within the sampled size range, and the estimate (15.3 cm) aligns with previous Mediterranean reports (Tom & Lewinsohn, 1983), suggesting that any selectivity bias is minimal.
The seasonal pattern of reproductive activity was clearly defined. GSI values and maturity-stage distributions indicated an extended spawning period from April to November, with a pronounced peak during July and August. This extended reproductive window is consistent with observations from other parts of the Mediterranean (Tom & Lewinsohn, 1983) but appears longer than that reported for some native populations in Japan, where spawning is typically confined to late spring and summer (Minagawa et al., 2000; Ohtomi et al., 2003). However, in warmer regions influenced by oceanic currents, such as Shibushi Bay, spawning periods may also extend into autumn (Ohtomi et al., 2003). The similarity between these warmer habitats and the eastern Mediterranean suggests that temperature regimes are a key factor shaping reproductive phenology. The prolonged spawning season observed here may enhance recruitment success and contribute to the species’ establishment and persistence in non-native environments. The observed temporal patterns in reproductive parameters are consistent with previous findings for penaeid species in the region. The availability of monthly sample sizes (Supplementary Table 1) provides additional support for the interpretation of these patterns.
The analysis of gonadal maturity stages corroborated the temporal pattern indicated by GSI values. Immature individuals dominated during the winter months, while mature and spawning females were frequent from late spring through summer. The presence of spent individuals from early summer onward indicated active spawning during this period. Such synchronization between physiological indices and morphological staging strengthens the reliability of the reproductive assessment and aligns with established methodologies in crustacean biology (Minagawa et al., 2000; Yano, 1988).
The estimated size at first sexual maturity (TL50 = 15.3 cm) represents a key life-history parameter with direct implications for fisheries management. This value is slightly higher than the 13.0 cm reported in the Ariake Sea, Japan (Minagawa et al., 2000), but falls within the 14–15 cm range previously observed in Mediterranean populations (Tom & Lewinsohn, 1983). Geographic variation in size at maturity is often linked to environmental factors such as temperature, food availability and population density (King, 1995). The TL50 estimated here suggests that a minimum landing size above 15.3 cm would allow most females to reproduce at least once before being harvested.
The ecological success of M. japonicus in the Mediterranean can be partly explained by the combination of life-history traits identified in this study. An extended reproductive season, flexible growth patterns and a moderate size at maturity are characteristics commonly associated with species capable of rapidly establishing populations in new environments (Albano et al., 2021; Tsirintanis et al., 2022). These traits likely enhance both reproductive output and resilience to fishing pressure, facilitating the species’ continued expansion in the region.
The successful establishment of M. japonicus in the Gulf of Antalya reflects its Lessepsian origin. Several life-history traits explain its ecological success: an extended reproductive season, sex-specific growth patterns and a moderate size at maturity (Albano et al., 2021; Tsirintanis et al., 2022). As a species originating from tropical and subtropical waters, M. japonicus appears to retain a flexible reproductive phenology. This flexibility allows it to adapt to the warm-temperate conditions of the eastern Mediterranean, where summer temperatures regularly exceed 20°C (Dall et al., 1991; IUCN MedMis, 2025; Kumlu et al., 1999). Its prolonged spawning season may also provide a demographic advantage over native species with shorter, more synchronized reproduction. Competition for food and habitat with native penaeid shrimps is, therefore, possible.
For fisheries management, the results support integrating biological reference points into regulatory frameworks. The coincidence of peak spawning activity with the summer months suggests that seasonal closures during this period could effectively protect reproductive individuals. Although trawl bans currently provide partial protection, additional measures targeting small-scale fisheries may be necessary to reduce fishing pressure during critical reproductive periods. Furthermore, implementing a minimum landing size above 15.3 cm would contribute to stock sustainability by allowing individuals to reach sexual maturity before harvest. These management measures are widely recognized as effective tools to protect spawning stock biomass and ensure long-term sustainability (Froese et al., 2016; Hilborn & Walters, 1992).
This study provides baseline data on the population structure, growth and reproductive biology of M. japonicus in the Gulf of Antalya. The sex ratio, positive allometric growth in females, negative allometric growth in males, extended reproductive season from April to November with peak spawning in July–August and size at first sexual maturity of 15.3 cm TL are key findings. These results contribute to understanding the species’ biology in the Mediterranean and provide essential parameters for the sustainable management of this valuable fishery resource. While the data were collected in 2018, the estimated biological reference points represent intrinsic species traits that remain valid for current fisheries management and provide a baseline for detecting future climate- or fishery-induced changes. Future research should focus on evaluating the potential for selective breeding programs in Mediterranean populations, as demonstrated in Australian and Indian stocks (Francis et al., 2021; Preston et al., 2004). The recent availability of a chromosome-level genome assembly (Wei et al., 2025) will facilitate genetic studies and support the development of marker-assisted selection for desirable traits such as growth rate and disease resistance.