
Raspberry (Rubus idaeus L.) is an economically important perennial crop with an annual production of almost one billion tonnes in the world, and Bosnia and Herzegovina (BiH) is one of the leading raspberry producers in the world (FAOSTAT, 2023). Raspberry production is affected by many pests and diseases, with viruses being of particular importance (Martin et al., 2013). Raspberry mosaic disease (RMD) caused by raspberry yellow net virus (RYNV), raspberry leaf mottle virus-RLMV or black raspberry necrosis virus (BRNV), as well as raspberry leaf blotch virus (RLBV) have been reported as the most important viral diseases threatening raspberry cultivation in BiH (Delic et al., 2020; Radulovic et al., 2021; Voruna et al., 2023). As viruses are intracellular pathogens, chemical control methods are inefficient. Viruses that spread within orchards via vectors from plant to plant are considerably more difficult to manage, and the removal of infected plant debris does not represent an effective preventive strategy (Jones, 2021). In view of these facts, novel plant protection methods need to be introduced to effectively control the spread of raspberry viruses. One promising option is the use of microbial bioagents capable of inducing systemic resistance in plants and increasing their level of self-defence against viral infection (Abdelkhalek et al., 2022). However, it is advantageous to know the natural composition of the plant microbiome and its interaction with disease in the search for effective bioagents or resistance inducers. The primary objective of this study was to investigate the variations in bacterial colonisation in raspberry samples from BiH in order to explore differences in microbial composition between virus-infected and virus-negative plants.
Leaves of raspberry plants (52 in total) were collected from 13 locations in Bosnia and Herzegovina during springs 2019-2023, including commercial raspberry orchards, nurseries and two samples from the border control (imported planting material from Serbia) (Supplementary materials, S1). Sample selection was performed to include equal numbers of leaves from each combination of cultivar, year, and location to minimize potential confounding effects. Leaves were sampled from different raspberry cultivars, including those showing virus-like symptoms such as blotching, chlorosis, fruit crumbling, and chlorotic spotting and lining, as well as asymptomatic plants. DNA was extracted according to the protocol described in Poudel et al. (2013). The samples were tested for the presence of ten viruses (RYNV, BRNV, RLBV, RLMV, raspberry bushy dwarf virus-RBDV, tobacco ringspot virus-TRSV, cherry leaf roll virus-CLRV, tomato ringspot virus-ToRSV, Arabis mosaic virus-ArMV, apple mosaic virus-ApMV (Delic et al., 2020; Radulovic et al., 2021; Voruna et al., 2023). We evenly (by infection, year, cultivar and location) selected already extracted total nucleic acid from 24 virus-infected and 28 virus-negative raspberry samples and subjected them to metabarcoding microbiome analysis. Chloroplast excluding primer pair 515/i799R was used for PCR amplification of V4 region of 16S rRNA gene (Artimová et al., 2022). Libraries were prepared with the TruSeq LT PCR-free kit and sequenced on an Illumina MiSeq platform (V3 600-cycle kit). The resulting sequences were demultiplexed and quality filtered using SEED2 (Větrovský et al., 2018). Sequences were clustered into OTUs and chimera cleared using Vsearch (Rognes et al., 2016). Singleton and doubleton OTUs were excluded from the analysis. The most abundant sequences from each OTU were identified using RDP classifier 2.13 (Wang et al., 2007). Diversity was analysed using the vegan package (Oksanen, 2013) in R (R core team, 2022). Differential abundance was analysed using LeFSe (Segata et al., 2011). LEfSe analysis was performed using an LDA score cut-off of 2.0 and a significance threshold of p < 0.05. Sequences are deposited under bioproject number PRJNA1184962.
Of the ten viruses the samples were tested for, only four were detected in the analysed raspberry plants: RLMV, RYNV, RLBV and RBDV. Apart from six samples positive for RYNV and two samples confirmed for RBDV, 12 samples were in double or and two in triple virus infection with causal agents of RMD (RLMV and RYNV) and RLBV. RBDV was found only in single infection in raspberry samples imported from Serbia.
A signifikant difference was found between the virus-infected and negative groups in the alpha diversity indices. Both Shannon's and Chao1 diversity indices were lower in virus-positive samples (Fig. 1A). The loss of diversity was also indicated by a lower evenness of the genus distribution. Nonmetric multidimensional scaling (NMDS) analysis showed that the bacteriome of virus infected and negative raspberry samples formed distinct clusters (Fig 1B). The hypothesis of a different microbial community in virus-infected plants was statistically confirmed by PERMANOVA analysis (p=0.001). Proteobacteria was the most abundant phylum in all samples (Supplementary Figure S2). Differential abundance analysis based on LeFSe (Supplementary materials, S3) showed a signifikantly higher proportion of Proteobacteria in virus-infected samples, while Firmicutes, Actinobacteria, Acidobacteria and Verrucomicrobia were more abundant in virus-negative plants. There are clear differences in the proportion of bacterial genera in virus-infected and virus-negative plants. The heat map of the most abundant genera (Supplementary materials, S4) shows distinct clusters of genera more abundant in virus positive/negative samples. Five genera including Methylobacterium, Sphingomonas, Massilia were signifikantly (p <0.05) more abundant in infected plants, while 10 genera such as Pantoea, Staphylococcus or Corynebacterium were more abundant in healthy plants according to Lefse analysis.
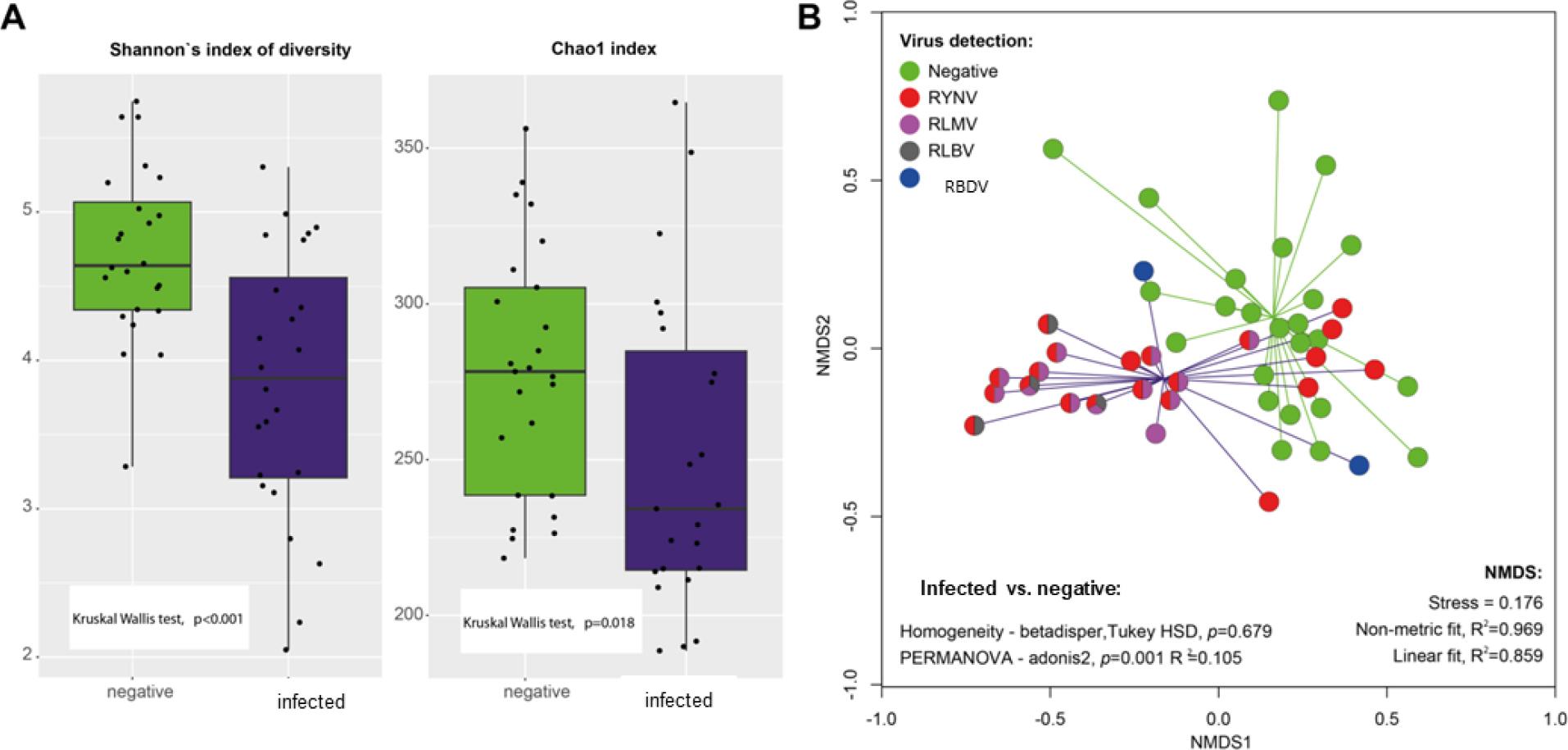
A) Alpha diversity indices of bacterial community in virus positive and negative raspberry samples; B) Beta diversity of prokaryotic community in virus positive and negative raspberry samples.
Significant changes in the raspberry bacteriome due to fungal infection were confirmed by Pylak et al. (2023). Despite their higher abundance in virus-infected plants, Methylobacterium or Sphingomonas are considered part of the core microbiome of several plants, including those of Rubus genera (Leino, 2023). On the other hand, Pantoea, which was more abundant in virus negative plants, is also very abundant in plant leaf bacteriomes and has been identified as an antagonist of many plant diseases Dutkiewicz et al. (2016). There are no available studies linking the raspberry microbiome to virus infection, but a study of aphid-borne Potato virus Y (PVY), Sugarcane mosaic virus (SCMV) and Wheat mosaic virus showed signifikant negative effects of plant viruses on root endophytes of the studied host plant (Cao et al., 2024; Liu et al., 2023; Zhang et al., 2022). There is a lack of information on how the process of bacterial colonisation of the plant is altered in response to virus infection, or whether the prokaryotic community was different prior to virus infection. Plants drastically alter their primary and secondary metabolism in response to active virus infection (Jiang and Zhou, 2023). Such changes may affect their colonisation by bacteria either through altered exudate composition, hormone manipulation, oxidative stress and subsequent reduction in plant defence capacity. This may explain the similarity of individual RYNV-infected samples, which harbour a community similar to that of healthy plants. RYNV is a DNA pararetrovirus that can occur in episomal form or integrate into the plant genome. When integrated into the plant genome, it cannot replicate and does not express viral symptoms, although it can be detected by molecular methods (Diaz-Lara et al., 2020; Harper et al., 2024). Both RMD and RLBV are semi-persistently transmitted by aphids and eriophyid mites, respectively (Martin et al., 2013), whereas RBDV is a pollen-borne virus (Martin et al., 2013). As RBDV-infected plant samples had a similar bacteriome to virus-negative samples, the transmission route of the viruses may also play a role in bacterial colonisation, possibly through co-transmission (Tamborindeguy et al., 2023). Generally, plants with mixed virus infections expressed stronger symptoms that influence yield quality, which could explain why the samples with mixed virus infections exhibited the strongest shifts in bacterial community composition.
This study reveals distinct differences in the leaf-associated bacterial communities of virus-infected and virus-negative raspberry plants. While lower diversity and compositional shifts were observed in virus-infected samples, these associations should not be interpreted as causal. Differences could reflect virus-driven changes, microbiome-mediated susceptibility, or co-occurring environmental factors. As samples were collected from multiple cultivars and sites, we cannot exclude the possibility that host genotype or environmental conditions contributed to the observed differences. Further experimental work is needed to clarify virus-bacteriome relationships.