
Figure 1:
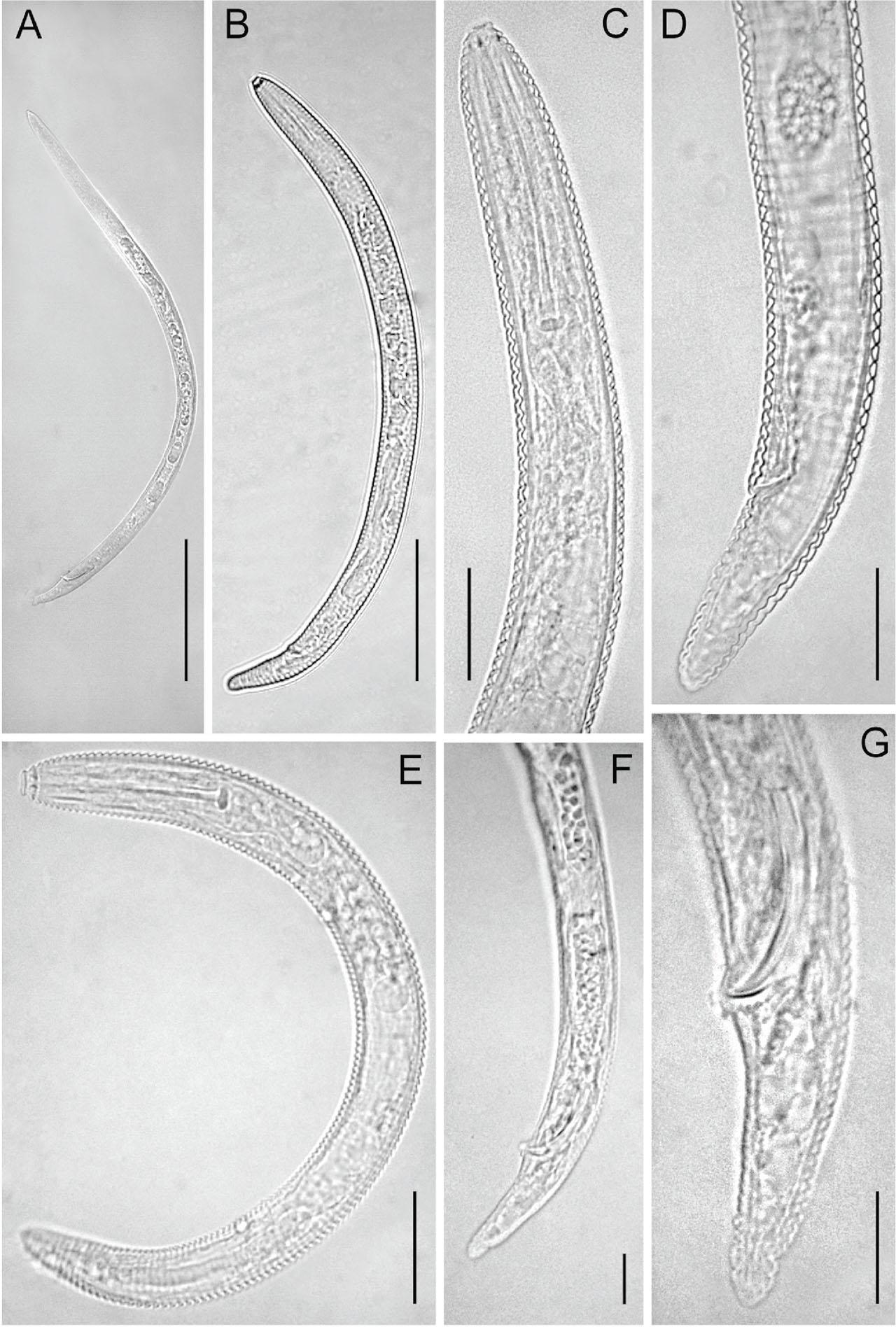
Figure 2:
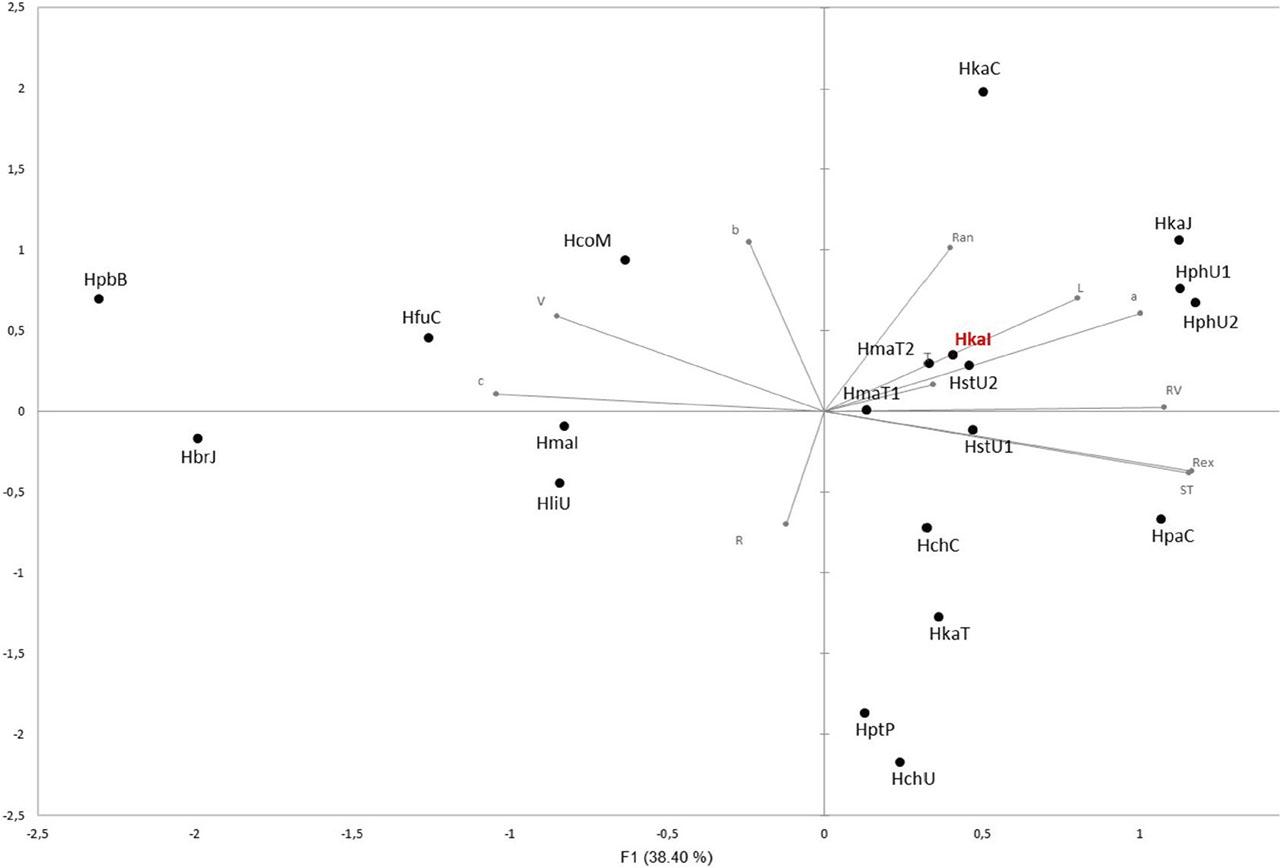
Figure 3:
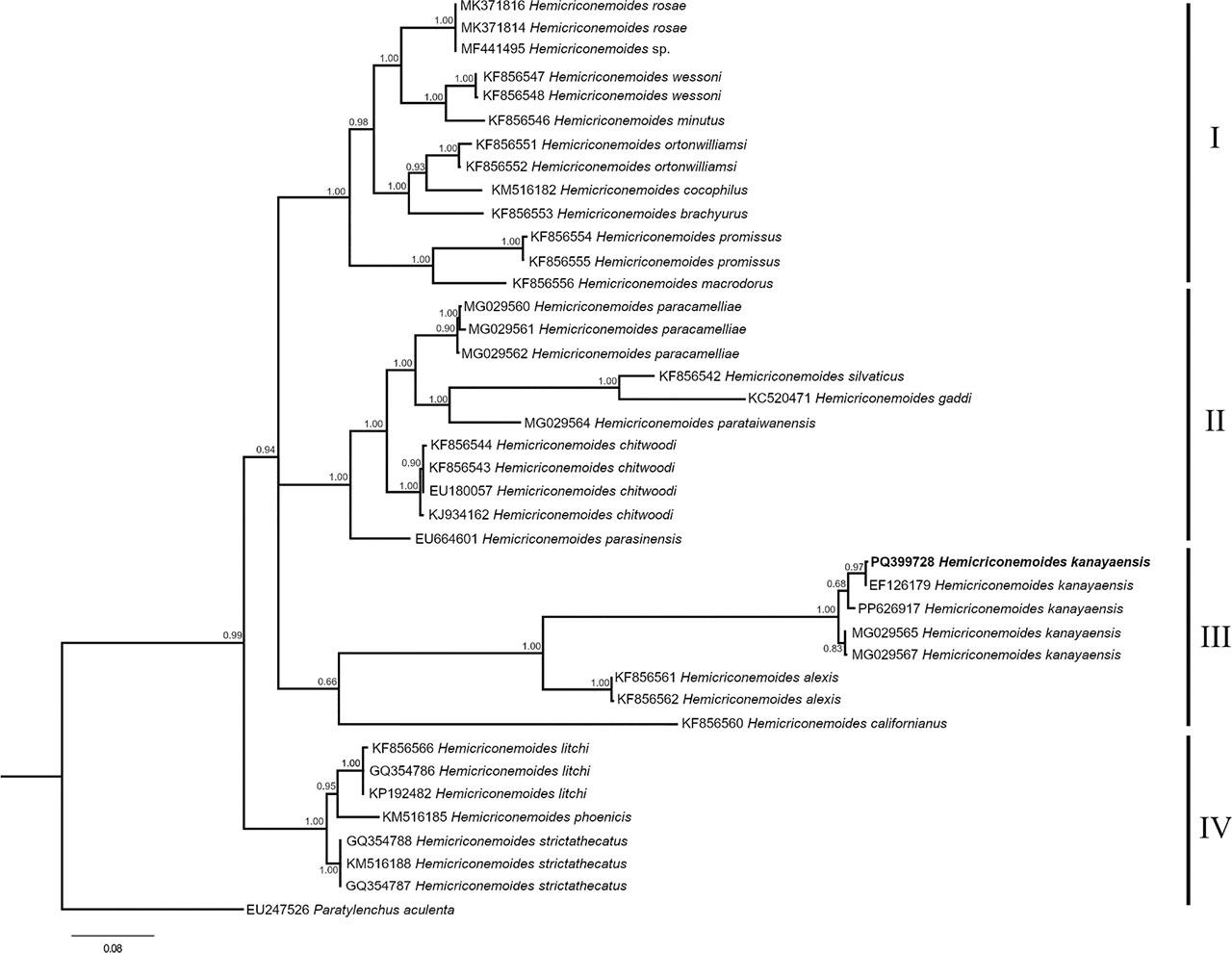
Figure 4:
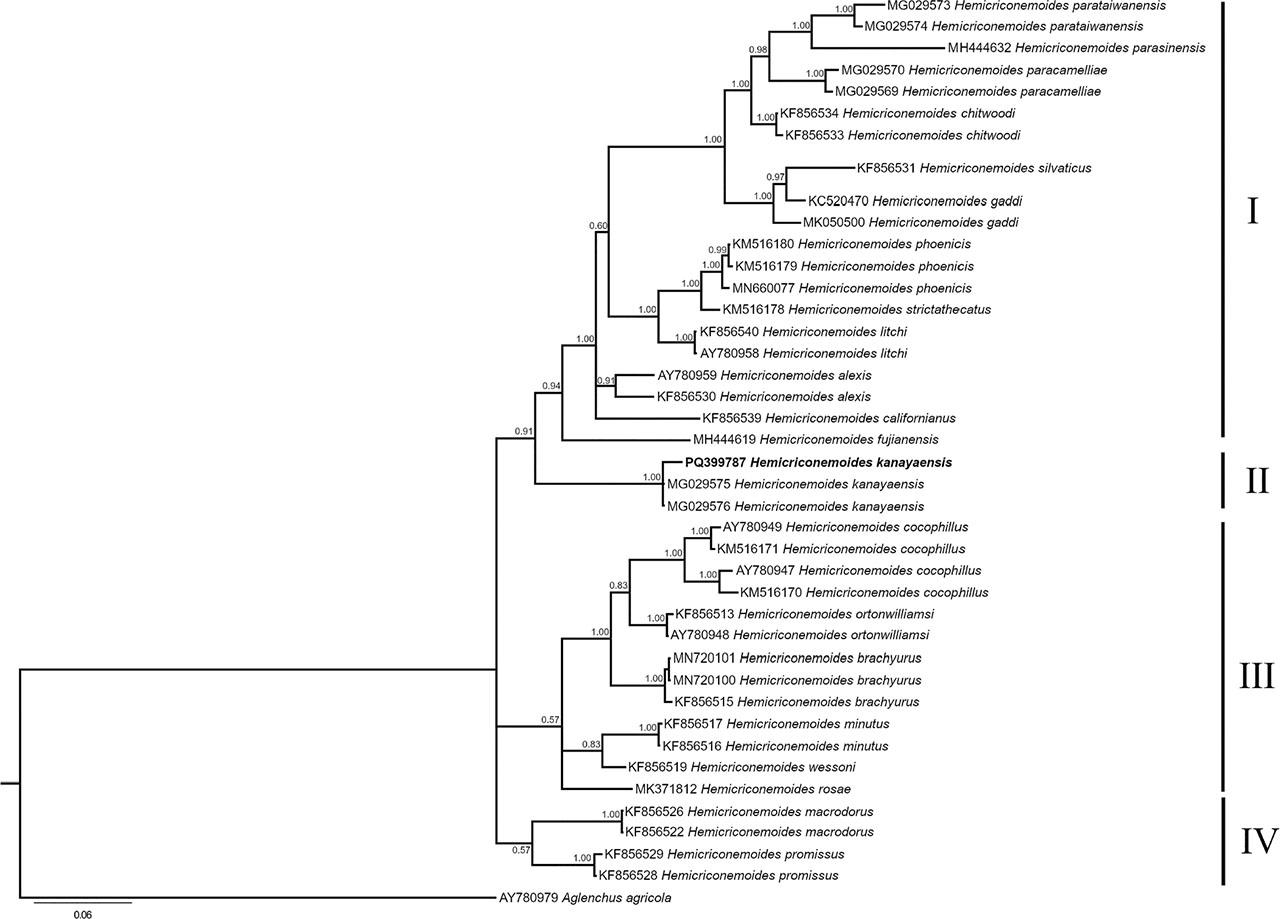
Figure 5:
Supplementary Figure 1:
Correlations between variables and components_
| F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | F9 | F10 | F11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L | 0,290 | 0,345 | 0,238 | −0,187 | −0,459 | −0,314 | −0,165 | −0,296 | −0,184 | −0,102 | 0,486 |
| a | 0,362 | 0,298 | 0,126 | −0,431 | 0,107 | −0,072 | 0,090 | 0,265 | 0,097 | −0,499 | −0,475 |
| b | −0,086 | 0,515 | −0,249 | −0,260 | −0,288 | 0,454 | −0,063 | −0,064 | 0,349 | 0,408 | −0,113 |
| c | −0,376 | 0,053 | 0,223 | −0,220 | 0,356 | −0,413 | −0,466 | −0,208 | 0,439 | 0,071 | −0,041 |
| V | −0,307 | 0,290 | 0,421 | −0,066 | 0,130 | −0,042 | 0,371 | 0,574 | −0,022 | 0,184 | 0,347 |
| R | −0,043 | −0,343 | 0,415 | −0,508 | 0,100 | 0,552 | −0,185 | −0,136 | −0,278 | 0,018 | 0,075 |
| Rex | 0,421 | −0,182 | 0,105 | −0,071 | 0,245 | 0,048 | 0,502 | −0,317 | 0,528 | 0,107 | 0,261 |
| RV | 0,390 | 0,012 | −0,319 | −0,002 | 0,334 | 0,140 | −0,484 | 0,421 | 0,085 | −0,008 | 0,443 |
| Ran | 0,144 | 0,497 | 0,078 | 0,234 | 0,574 | 0,096 | 0,023 | −0,348 | −0,413 | 0,170 | −0,108 |
| ST | 0,418 | −0,188 | 0,230 | −0,031 | −0,113 | −0,279 | −0,126 | 0,214 | −0,096 | 0,681 | −0,333 |
| T | 0,125 | 0,082 | 0,543 | 0,584 | −0,166 | 0,321 | −0,264 | 0,053 | 0,314 | −0,175 | −0,097 |
Morphometric data of male of Hemicriconemoides kanayaensis_ All measurements are in μm and in the form: mean ± s_d_
| Character | This study, Iran | Chen et al. 2007, Taiwan (Pinglin) | Chen et al. 2007, Taiwan (Rueisuei) | Nakasono & Ichinohe 1961, Japan |
|---|---|---|---|---|
| N | 8 | 8 | 8 | 7 |
| L | 437 ± 29 (396–473) | 420 ± 10 (400–440) | 430 ± 30 (400–460) | 457 (422–489) |
| a | 27.6 ± 1.7 (25.9–29.5) | 28.9 ± 2.9 (24.7–33.9) | 29.8 ± 2.3 (26.7–33.9) | 29.7–32.6 |
| c | 15 ± 1.06 (13.9–16.6) | 15.5 ± 1.1 (13.8–17.5) | 16.4 ± 1.2 (14.8–17.9) | 14.6 (14.6–15.1) |
| c′ | 2.37 ± 0.26 (2.14–2.61) | 2.6 ± 0.2 (2.2–2.8) | 2.6 ± 0.2 (2.1–2.7) | - |
| EP | 91 ± 7 (83–100) | 99 ± 5 (92–107) | 86 | |
| Max. body diam | 15.9 ± 1.24 (14.6–17.7) | - | - | - |
| Anal body diam. (ABD) | 12.5 ± 1.16 (11.5–13.6) | 10 ± 1 (10–11) | 10 ± 1 (10–11) | - |
| Tail length (T) | 29.3 ± 2.74 (26.2–33) | 27 ± 2 (24–31) | 26 ± 2 (23–29) | - |
| Spicule | 23.1 ± 1.81 (20.5–24.6) | 26.5 (n=4) (25.7–26.7) | 25.4 ± 1.1 (24.2–27.0) | 23.8 |
| Gubernaculum | 4.83 ± 1.16 (3.7–6.9) | - | - | - |
Morphometrics of Hemicriconemoides kanayaensis Nakasono & Ichinohe, 1961_ female_ All measurements are in μm and in the form: mean ± s_d_ (range)_
| Character | This study Iran | Maria et al., 2018 China | Nakasono & Ichinohe, 1961 Hangzhou Japan (Type Pop.) | Germani & Anderson, 1991 Taiwan | Chen et al., 2007 Taiwan |
|---|---|---|---|---|---|
| N | 10 | 15 | 20 | 12 | * |
| L | 494 ± 41.77 (409–560) | 601 ± 43.2 (500–663) | 571 (500–631) | 510 (470–540) | (430–600) |
| Rst | 24 ± 1.71 (21–27) | 21.6 ± 1.0 (20.0–24.0) | - | - | - |
| ROes | 35 ±1.89 (33–39) | 30.4 ± 1.3 (29.0–33.0) | - | - | - |
| Rex | 35 ± 2.2 (31–35) | 33.2 ± 1.3 (31.0–36.0) | 37 (30–44) | 35 (31–38) | (35–41) |
| Rv | 16.4 ± 14 (15–19) | 15.1 ± 1.1 (14.0–17.0) | 18 (16–21) | 17 (16–18) | (13–19) |
| Rvan | 5 ± 0.5 (5–6) | 4.8 ± 0.7 (4.0–6.0) | - | - | - |
| Ran | 12 ±1.7 (11–17) | 10.3 ± 0.6 (9.0–11.0) | 12 (11–15) | 10 (8–11) | (9–13) |
| a | 14.6 ± 1.3 (11.–16.8) | 20.5 ± 1.9 (17.6–24.4) | 21.5 (18.7–24.4) | 17.3 (15.8–18.4) | (14.8–20.7) |
| b | 4.6 ± 0.44 (3.9–5.2) | 5.0 ± 0.3 (4.6–5.8) | 4.8 (3.3–5.6) | 4.8 (4.4–5.3) | (3.6–5.3) |
| c | 14.5 ± 1.4 (13.6–16.6) | 15.7 ± 1.2 (13.9–17.8) | 14.3 (11.5–16.8) | 12.9 (12–14.7) | (11.8–18.3) |
| c’ | 1.84 ± 0.18 (1.5–2.1) | 2.0 ± 0.1 (1.8–2.3) | - | - | (1.5-2.5) |
| m | 83.2 ± (81.3–85) | 87.3 ± 1.3 (84.9–89.1) | - | - | - |
| V | 89.1 ± 1.5 (88.4–90.7) | 93.0 ± 0.6 (92.1–94.4) | 88.9 (87.5–91.5) | 88.3 (86.4–89.2) | (87.3–90.5) |
| VL/VB | 2.26 ± 0.23 (1.95–2.7) | 1.8 ± 0.1 (1.6–2.1) | - | - | - |
| Stylet | 70.9 ± 4.6 (65.9–78.2) | 76 ± 2.7 (72–82) | 74 (66–79) | 75 (66–78) | (69–79) |
| ST%L | 6.9 ± 1.2 (6.2–7.1) | 12.6 ± 0.9 (11.4–14.4) | - | - | - |
| Stylet knob length | 2.6 ± 0.41 (2–3.3) | 3.3 ± 0.4 (3.0–4.2) | - | - | - |
| Stylet knob width | 6.83 ± 0.79 (6–8.8) | 6.7 ± 0.4 (5.9–7.5) | - | - | - |
| DGO | 5.4 ± 0.3 (5.1–5.9) | 5.9 ± 0.5 (5.1–6.5) | - | - | - |
| Pharynx | 107 ± 9.8 (104.5–116.3) | 119 ± 6.3 (107–129) | - | - | - |
| Anterior to excretory pore | 98.9 ±10.7 (88.3–116) | 130 ± 6.4 (115–138) | - | - | (92–144) |
| Max. body diam | 33.9 ± 1.7 (31–36) | 29.5 ± 2.4 (26.0–34.0) | 27 (22–29) | - | - |
| Vulva body diam. (VD) | 22 ± 1.8 (20.7–25.5) | 22.8 ± 1.3 (20.0–25.0) | - | - | (18–29) |
| Vulva to tail tip | 49.9 ± 7.35 (42–69) | 42 ± 3.0 (36–46) | - | - | - |
| Anal body diam. (ABD) | 17.9 ± 1.59 (16–21.3) | 19.3 ± 1.1 (17.0–21.5) | - | - | - |
| Tail length (T) | 34.3 ± 4.16 (26.6–40) | 38 ± 3.0 (33–46) | - | - | - |