
The livestock sector is a vital part of the national economy. In rural areas, 35 – 40 million people rely on livestock for 30 – 40 % of their income. Animal husbandry in Pakistan significantly contributes to livelihoods and poverty alleviation (Rehman et al., 2017; Riaz, 2022). Pakistan has 82.5 million goats and 31.9 million sheep, with Khyber Pakhtunkhwa hosting 9.50 million goats and 3.30 million sheep. Small ruminants are critical to the sustainability of agriculture in developing countries and contribute to a variety of socioeconomic activities across the world (Ministry of Finance, Government of Pakistan, 2022; Herrero et al., 2013).
Open grazing and extensive farming methods in sheep and goats increase the risk of worm infection and helminthiasis because they graze on worm larvae and eggs that have adhered to the grasses and pastures (Ijaz et al., 2009). Haemonchus, a barber pole or wireworm, infects millions of sheep and goats worldwide, causing massive economic losses (Carson et al., 2023; Akkari et al., 2013). Haemonchosis also negatively impacts the economy and drastically lowers the productivity of the affected sheep. In animals of all ages, adult worms cause edema, diarrhea, anemia, and occasionally even death by sucking the blood from the abomasum of sheep and goats (Osman et al., 2018; Santos et al., 2012; Ahmad et al., 2024). Haemochus contortus worms sucks around 0.05 ml of blood each day, causing anemia, meat and wool loss, and biochemical disruptions (Khattak et al., 2023; Rowe et al., 2008). Haemonchus produces up to ten thousand eggs daily, expelled in host feces, infecting millions of small ruminants worldwide (Qamar & Alkheraije, 2023).
Haemonchus, a new anthelmintic-resistant parasite, must be accurately identified to be controlled. Genetic exchange between small and large ruminants and shared pastures contributes to the transmission of resistant genes (Amarante et al., 2017). The traditional coprological diagnostic techniques for parasite species are challenging. Although morphology is helpful, it might not always resolve the taxonomic issues regarding species identification (Roeber et al., 2013; Wyrobisz et al., 2016). Because of their specificity, sensitivity, and precision, molecular techniques like polymerase chain reaction (PCR) help detect parasite species, particularly strongyle nematodes, and are, therefore, a valuable tool for taxonomic studies (Arbabi et al., 2020; Grillo et al., 2007).
A molecular marker, ITS-2, is used to identify closely related species. Similarly, the Cox 1 gene, mitochondrial DNA (mtDNA), is another molecular marker used for studying parasite genetic diversity due to its strong maternal lineage and rate of mutation (Mishra et al., 2021; Chrisanfova et al., 2021; Kandil et al., 2018).
According to previous studies conducted in Pakistan, the prevalence of Haemonchus ranges from 6.5 % to 87 % (Lashari & Tasawar, 2011; Sajid et al., 2000) as shown in Table 1. Small ruminants in Pakistan are affected by haemonchosis, which causes severe economic losses. Effective control requires a full understanding of the epidemiological factors driving disease progression, although there are no microscopic, morphometric, or phylogenetic studies available in the Malakand division. Therefore, this study aims to provide the first comprehensive study on Haemonchus in the Malakand division, which includes epidemiology, morphometric characterisation, and phylogenetic analysis. The work will use molecular and microscopic methods to resolve taxonomic ambiguities, accurately differentiate Haemonchus from closely related species and improve parasite identification procedures. It will also provide insights into the genetic patterns and evolutionary relationships of Haemonchus populations in Pakistan. The current study will increase our knowledge of Pakistan’s haemonchosis epidemiology, including its origin, transmission, and population structure, which could drive targeted control strategies and guide government actions to reduce the spread of the disease.
Epidemiological spectrum of Haemochus contortus among sheep and goat populations in Pakistan.
| Study area | Host | Sample type | Method | Samples | Positive | Prev. % | QAS | Reference |
|---|---|---|---|---|---|---|---|---|
| KPK | Goat & Sheep | Fecal | Mic & Mol | 180 | 46 | 25.55 | 5 | Ahmad et al., 2024 |
| Sindh | Goat & Sheep | Fecal | Mic & Mol | 800 | 88 | 11 | 4 | Memon et al., 2024 |
| KPK | Goat & Sheep | Fecal | Mic, Floa & Sed | 1340 | 312 | 23.28 | 4 | Irshad et al., 2023 |
| KPK | Goat & Sheep | Fecal | Floa & MM | 400 | 57 | 14.25 | 5 | Khan et al., 2022 |
| KPK | Goat & Sheep | Blood & Fecal | Mic | 300 | 56 | 18.66 | 2 | Ali et al., 2022 |
| KPK | Goat & Sheep | Fecal | Mic | 480 | 63 | 13.12 | 3 | Aimen et al., 2022 |
| KPK | Goat & Sheep | Fecal | Mic | 315 | 71 | 22.53 | 3 | Ruhoollah et al., 2021 |
| Punjab | Goat & Sheep | Worm | Mic | 200 | 105 | 52.2 | 3 | Bibi et al., 2017 |
| KPK | Sheep | Fecal | Floa, sed & MM | 300 | 120 | 40 | 4 | Jamil et al., 2016 |
| Punjab | Goat & Sheep | Fecal | Mic & EPG | 646 | 178 | 27.55 | 5 | Qasim et al., 2016 |
| KPK | Goats | Fecal | Mic | 150 | 22 | 14.66 | 4 | Nabi et al., 2014 |
| Punjab | Goat & Sheep | Fecal | Floa & Sed | 1000 | 133 | 13.3 | 4 | Raza et al., 2014 |
| Punjab | Goat | Fecal | Floa & Sed | 100 | 20 | 20 | 3 | Ayaz et al., 2013 |
| Punjab | Goat & Sheep | Fecal | Floa, Sed & MM | 480 | 137 | 28.54 | 4 | Khalid et al., 2013 |
| Balochistan | Sheep | Fecal | McMaster | 1200 | 125 | 10.42 | 2 | Razzaq et al., 2013 |
| KPK | Sheep | Fecal | Mic & Floa | 356 | 190 | 53.37 | 3 | Ullah et al., 2013 |
| Punjab | Sheep | Fecal | Mic, Floa & Sed | 523 | 34 | 6.5 | 4 | Lashari & Tasawar, 2011 |
| Sindh | Goat | Fecal | Mic | 1065 | 156 | 14.65 | 3 | Akhter et al., 2011 |
| Punjab | Sheep | Fecal | MM | 333 | 259 | 77.7 | 3 | Tasawar et al., 2010 |
| Punjab | Goat & Sheep | Worm | K/Iden | 4740 | 1604 | 33.83 | 1 | Raza et al., 2009 |
| Punjab | Goat & Sheep | Fecal | Mic, Floa & Sed | 400 | 225 | 56.25 | 1 | Gadahi et al., 2009 |
| Punjab | Goat | Fecal | Mic, Floa & MM | 300 | 84 | 28 | 2 | Ijaz et al., 2008 |
| Punjab | Sheep | Worm | Mic & K/Iden | 960 | 591 | 61.5 | 3 | Lateef et al., 2005 |
| Punjab | Sheep | Fecal | Mic | 10000 | 5477 | 54.77 | 2 | Jabeen et al., 2000 |
| Islamabad | Sheep | Fecal | Floa & MM | 62 | 54 | 87 | 0 | Sajid et al., 2000 |
Mic: Microscopy; Floa: Floatation; Sed: Sedimentation; MM: McMaster; K/Iden: Key identification; KPK: Khyber Pakhtunkhwa
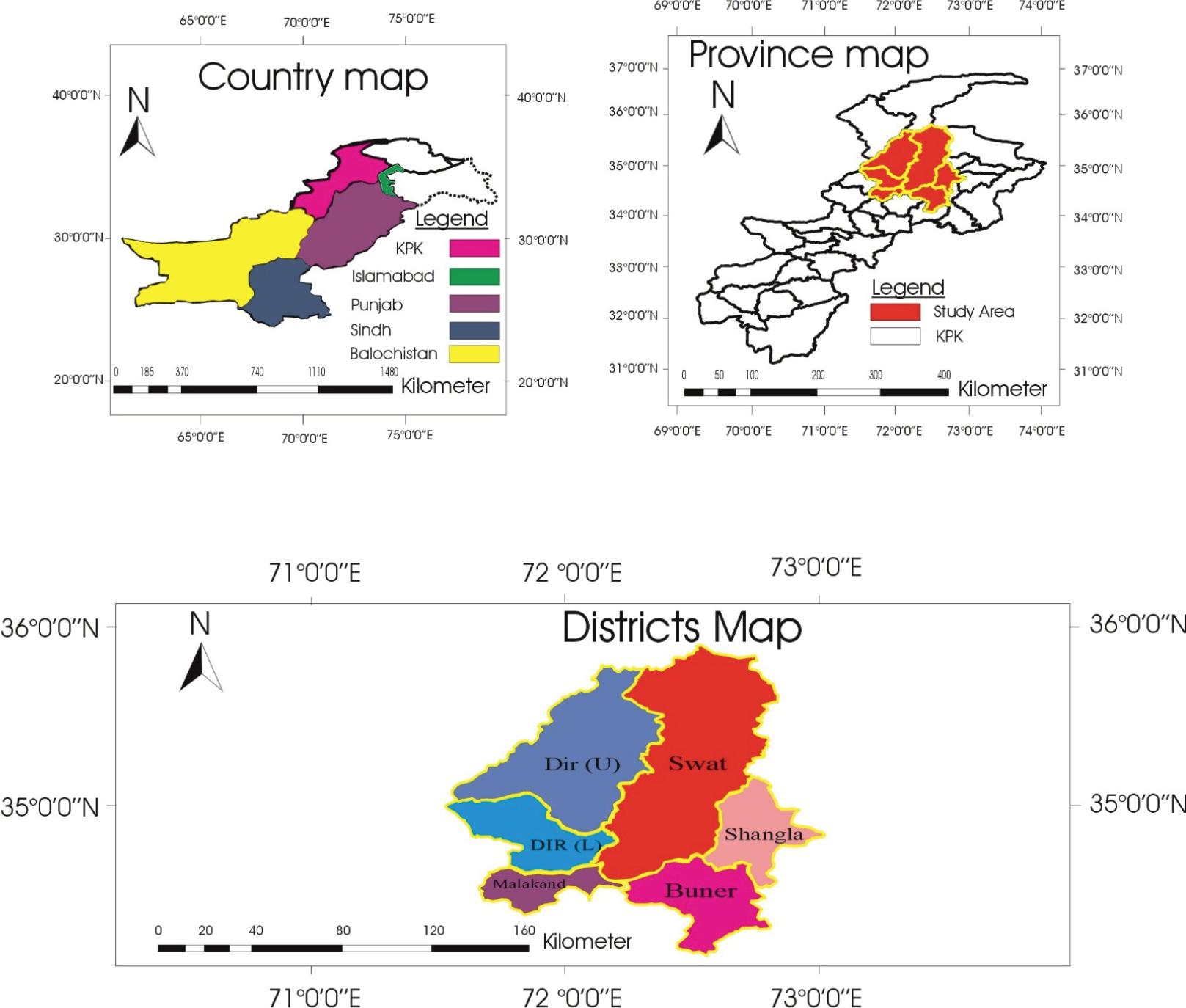
The study area map displays the geographical distribution of sampling locations in the Malakand Division of Khyber Pakhtunkhwa, Pakistan. It includes a national context map (country map), a provincial focus map (province map), and detailed district boundaries within the study area, highlighting Dir Upper, Dir Lower, Swat, Malakand, Shangla, and Buner. The map was designed and modified by the authors.
The study was conducted in Malakand division, Khyber Pakhtunkhwa, Pakistan (33° 59’ 59.89” N latitude and 72° 56’ 2.72” E longitude). Malakand Division is renowned for its extensive livestock farming and variety of agricultural activities. The region experiences average annual rainfall of 8.4 inches and temperatures ranging from -1°C in January to 42°C in July (Ali et al., 2021; Irshad et al., 2023).
In total, 878 (Goats: 398 and Sheep: 480) fecal samples were randomly gathered from sheep and goats across four seasons (January to December, 2024) from six districts of Malakand division, Pakistan, using a multistage random sampling design. Every fourth or eighth animal was sampled, depending on the flock size. To avoid repetition, no herd or animal was sampled more than once. Animals aged 3 months or older were included in the study; both healthy and unhealthy animals, regardless of treatment history, were sampled to allow comparative analysis. Pregnant, lactating and severely injured animals were excluded. Samples were collected directly from the rectum of sheep and goats. Comprehensive data encompassed animal category, time, gender, season, region, age, treatment history, dietary system, water source, and health status. Herd-level clustering was considered negligible, and no mixed effects modelling was applied due to the limited number of animals sampled per herd and a one-time visit design. Samples were examined at the Parasitology laboratory, Department of Zoology, University of Malakand, using the direct smear method under a microscope at 4X, 10X, and 40X. The Haemonchus eggs were identified based on the standard keys described by Mussa (2023) and Soulsby (1982).
A total of 100 abomasa (50 goats, 50 sheep) were collected from local abattoirs of the study area and processed immediately in the laboratory. A total of 1200 adult Haemonchus were recovered using the procedures described by Kuchai et al. (2012). The worms were then cleaned in normal saline and washed in phosphate-buffered saline (PBS; pH=7.4) containing ampicillin (100 mg/ml) and cloxacillin (50 mg/ml) to inhibit microbial growth. Subsequently, Haemonchus was identified based on morphological characteristics using the key described by Soulsby (1982). Parasites were stored at -20°C in 70 % glycerin alcohol solution for molecular analysis.
For morphometric characterization, 35 samples of Haemonchus eggs and 126 (76 females and 50 males) samples of adult worms were randomly selected for measurement. Parasitic eggs and adult worm measurements were conducted using a compound microscope (Olympus BX 53, Tokyo, Japan). Body length, body width, esophageal length, spicule length, gubernaculum length, vulva to posterior end distance, and anus to posterior end distance were measured in millimeters. In addition, the egg morphology, including the egg’s length, width, and shape, was evaluated as described by Lichtenfels et al. (1994) and Soulsby (1982).
Genomic DNA was extracted from individual Haemonchus samples using a commercial DNA extraction kit (Qiagen, Hilden, Germany), following the manufacturer’s guidelines.
To amplify the mitochondrial COX-1 gene for phylogenetic analysis, a polymerase chain reaction (PCR) was performed following Kandil et al. (2018). The primer sets used were LCO1490 (5' GGTCAACAAATCATAAAGATATTGG 3') as a forward primer and HCO2198 (5' TAAACTTCAGGGTGACCAAAAAATCA 3') as the reverse primer. Thermocycling conditions: 95°C for 5 minutes, 30 cycles of 95°C for 60 seconds, 55°C for 45 seconds, 72°C for 45 seconds, and a final extension at 72°C for 7 minutes. The amplified Cox-1 gene PCR products were visualized through agarose gel electrophoresis, as described by Ndosi et al. (2023).
The amplicons were confirmed through gel electrophoresis. Two amplified DNA products (one each from goats and sheep samples) were randomly selected, purified, and sequenced at Macrogen Inc. in Seoul, South Korea. The sequences were assembled and aligned using ClustalW multiple alignment implemented in MEGA11. Sequence analysis was performed using BLASTn algorithms and databases from the National Center for Biotechnology Information (NCBI) database. A phylogenetic tree was constructed using a neighbor-joining method implemented in MEGA 11.
Data were analyzed using SPSS 20 and GraphPad Prism V.7.0. Correlation between risk factors and Haemonchus prevalence was assessed through bivariate analysis using Chi-square tests. Descriptive statistics (standard deviation, error, and variance) were calculated. A p-value < 0.05 indicated statistical significance.
The University of Malakand’s advanced studies board approved the study protocols (UOM/Admin/2024/501). Informed consent was obtained from the owners of small ruminants that participated in the study. The authors declare that there are no conflicts of interest related to this study.
During the study period from January to December (2024), 878 fecal samples (Goats: 398 and Sheep: 480) were collected randomly from goats and sheep across six districts of the Malakand Division, namely (Swat, Shangla, Dir L, Dir U, Buner, and Malakand). Two hundred twenty-one (221) small ruminants tested positive for Haemonchus, resulting in an observed total prevalence of Haemonchus of 25.17 %. Sheep had a slightly greater prevalence of haemonchosis (26.04 %) than goats (24.12 %), although the difference was not statistically significant (p > 0.05) (Table 2).
Overall prevalence of H. contortus in goats and sheep in Malakand division.
| Host | Sample | Positive | Percentage | P-value |
|---|---|---|---|---|
| Goats | 398 | 96 | 24.12 | 0.51379ns |
| Sheep | 480 | 125 | 26.04 | |
| Total | 878 | 221 | 25.17 |
P-value is non-significant
In the current study, female sheep exhibited a higher prevalence of Haemonchus (27.86 %) compared to male sheep (23.5 %), as well as female goats (25.22 %) and male goats (22.62 %). However, this difference was not statistically significant (p>0.05).
Goats and sheep were categorized into three age groups: less than 1 year, 1 to 5 years, and greater than 5 years. Haemonchus infection rates varied significantly by age group (p < 0.05). Specifically: Younger animals (<1 year) had the highest infection rates, with 29.25 % in goats and 27.13 % in sheep, followed by older animals (>5 years) years Sheep 28.82 % and goats 27.03 % while animals aged (1 – 5) years had the lowest infection rates: 17.36 % in goats and 22.65 % in sheep.
This study was conducted across six districts of the Malakand division: Swat, Shangla, Dir (L), Dir (U), Malakand, and Buner. The prevalence of haemonchosis varied across districts, with Swat (26.22 %), Shangla (24.44 %), Dir (L) (26.28 %), Dir (U) (23.08 %), Malakand (26.25 %), and Buner (24.06 %). However, these differences were not statistically significant (p>0.05). Furthermore, the prevalence of haemonchosis was highest in grazing sheep (27.84 %) and goats (26.19 %) compared to stall-fed sheep (24 %) and goats (21.81 %), though the difference was not statistically significant (p>0.05).
Concerning health status, a statistically significant variation (p<0.05) was observed; the highest prevalence of haemonchosis was observed in poor goats (36.73 %) and sheep (35.71 %), followed by moderate sheep (30.11 %) and goats (26.97 %). The lowest prevalence rate was found in healthy goats (18.78 %) and sheep (20.59 %).
In addition, a high degree of Haemonchosis was observed in open water drinking goats (28.46 %) and sheep (28.39 %), followed by stream water sources drinking sheep (28.93 %) and goats (25.56 %) compared to other water sources like tap and bore water, the difference was not statistically significant (p>0.05). The prevalence of Haemonchosis in goats and sheep was further evaluated in different seasons. The highest prevalence was found in summer (27.69 %), while the lowest prevalence was recorded in winter (22.11 %), though this difference was not statistically significant (p>0.05).
Furthermore, statistically significant (p<0.05) variations between treated and untreated small ruminants were observed. The highest infestation was observed in untreated sheep (29.79 %) and goats (28.19 %), while the lowest prevalence was found in treated goats (16.55 %) and sheep (17.88 %), Table 3.
Prevalence of Haemonchus contortus in small ruminants concerning various risk factors in Malakand division.
| Risk Factors | Variables | Total samples | Total positive cases | Total prevalence (%) | Goats Positive (%) | Sheep Positive (%) | P-value |
|---|---|---|---|---|---|---|---|
| Gender | Male | 368 | 85 | 23.1 | 22.62 | 23.50 | 0.2292ns |
| Female | 510 | 136 | 26.7 | 25.22 | 27.86 | ||
| Age | <1 years | 235 | 66 | 28.1 | 29.25 | 27.13 | 0.0391* |
| 1-5 years | 325 | 66 | 20.3 | 17.36 | 22.65 | ||
| >5 years | 318 | 89 | 28 | 27.03 | 28.82 | ||
| Area | Swat | 164 | 43 | 26.2 | 23.19 | 28.42 | 0.9815ns |
| Shangla | 135 | 33 | 24.4 | 23.61 | 25.40 | ||
| Dir (L) | 156 | 41 | 26.3 | 25.00 | 27.50 | ||
| Dir (U) | 130 | 30 | 23.1 | 23.21 | 22.97 | ||
| Malakand | 160 | 42 | 26.2 | 25.76 | 26.60 | ||
| Buner | 133 | 32 | 24.1 | 23.73 | 24.32 | ||
| Grazing system | Grazing | 465 | 126 | 27.1 | 26.19 | 27.84 | 0.1629ns |
| Stall-fed | 413 | 95 | 23 | 21.81 | 24.00 | ||
| Health condition | Healthy | 435 | 86 | 19.8 | 18.78 | 20.59 | 0.0003* |
| Moderate | 338 | 97 | 28.7 | 26.97 | 30.11 | ||
| Poor | 105 | 38 | 36.2 | 36.73 | 35.71 | ||
| Drinking water source | Open | 285 | 81 | 24.4 | 28.46 | 28.39 | 0.1729ns |
| Bore | 187 | 40 | 21.4 | 20.45 | 22.22 | ||
| Stream | 211 | 58 | 27.5 | 25.56 | 28.93 | ||
| Tap water | 195 | 42 | 21.5 | 20.00 | 22.86 | ||
| Season | Autumn | 205 | 48 | 23.4 | 21.98 | 24.56 | 0.5016ns |
| Winter | 190 | 42 | 22.1 | 20.51 | 23.21 | ||
| Spring | 223 | 59 | 26.5 | 25.23 | 27.59 | ||
| Summer | 260 | 72 | 27.7 | 27.05 | 28.26 | ||
| Treatment | Treated | 290 | 50 | 17.2 | 16.55 | 17.88 | 0.0004* |
| Untreated | 588 | 171 | 29.1 | 28.19 | 29.79 |
P-value is non-significant
statistically significant
The present study randomly selected 35 positive samples from different small ruminants for egg morphometric characterization. Haemonchus eggs were oval, with equal poles and divided cells that did not fill the egg cavity (Fig. 2A). The average length of the eggs was 80.98μm, ranging from 63μm to 99μm. Egg width ranged from 36μ to 61μ, with an average width of 45.04μ. (Table 4).
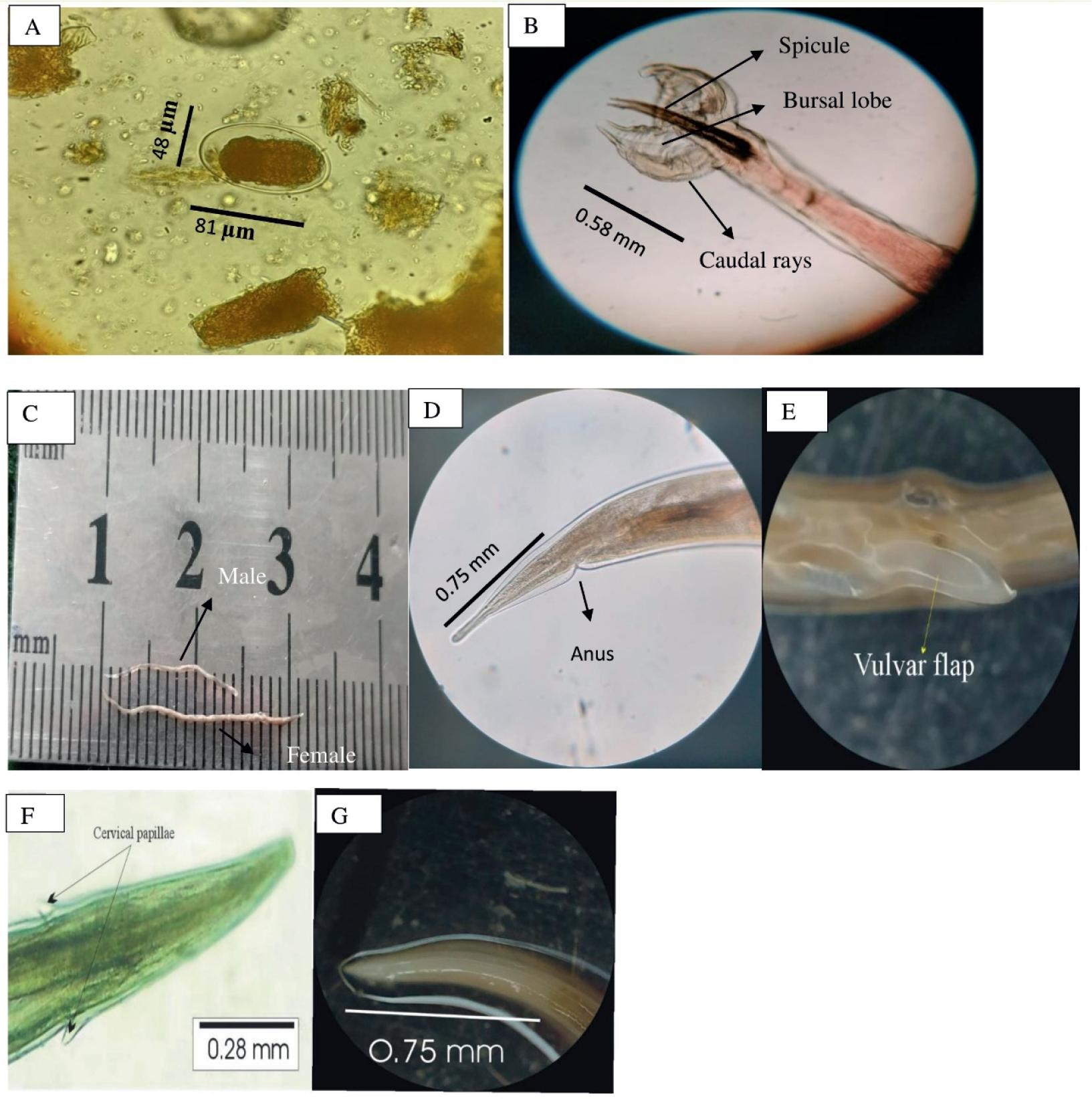
Morphological features of Haemonchus contortus.
(A). Egg containing morula, showing measured length and width (μm); (B). Male bursa highlighting spicules and caudal rays, with spicule length (mm); (C). Adult male and Female H. contortus aligned on a calibrated scale to show total body length (mm); (D). Posterior end of female showing anus and measured tail length (mm); (E). Vulvar flap of female Haemonchus; (F). Cervical papillae with recorded length (mm); (G). Oesophagus of H. contortus showing total length (mm)
Morphometric characteristics of H. contortus egg sizes (in μm).
| Shape | Statistical index | Length | Width |
| μm | μm | ||
| Oval | Average | 80.98 | 45.05 |
| Maximum | 99 | 61 | |
| Minimum | 63 | 36 | |
| Standard error | 1.6016 | 1.43 | |
| Standard deviation | 9.4749 | 8.49 | |
| Sample variance | 89.7754 | 72.14 | |
| N | 35 | 35 |
N: Number of eggs examined
A total of 1200 Haemonchus worms were recovered from 100 abomasums collected from the different abattoirs of the study area. One hundred twenty-six worms (76 females and 50 males) were randomly selected (from goats and sheep) for morphometric analysis. The results are summarized in Table 5. The anterior end was broad and blunt, with a small buccal cavity featuring a visible tooth protruding from the dorsal wall. The cervical papillae of both male and female worms possessed a spine-like protrusion on its outside that ran almost parallel to the longitudinal cuticular ridges. The body also had transverse striations, which were visible.
Morphometric measurement (in mm) of H. contortus isolated from goats and sheep.
| Particulars | Male worms (n= 50) | Female worms (n= 76) |
|---|---|---|
| Body length (mm) | 11.9 – 13.8 | 15 – 23.8 |
| Body width (mm) | 0.23 – 0.38 | 0.29 – 0.38 |
| Cervical papillae (mm) | 0.28 – 0.38 | 0.24 – 0.42 |
| Spicule length (mm) | 0.44 – 0.78 | – |
| Length of esophagus (mm) | 0.61 – 0.92 | 0.72 – 1.22 |
| Gubernaculum length (mm) | 0.09 – 0.18 | – |
| Distance from vulva to posterior end (mm) | – | 0.68 – 0.82 |
| Distance from Anus to posterior end (mm) | – | 0.29 – 0.35 |
Male worms were smaller than females, measuring 11.9 to 13.8 mm in length and 0.23 to 0.38 mm in width (Fig. 2C). The cervical papillae ranged from 0.28 to 0.38 mm, whereas the esophagus measured 0.61 to 0.92 mm. A bursa with two large lateral lobes and a small dorsal lobe was observed at the tail (Fig. 2B). The asymmetrically branched dorsal ray divided near the tip, creating a short external branch. Two 0.44 to 0.78 mm long spicules with a bulb and pore toward the end were supported by a spindle-shaped gubernaculum tapered toward the tail end and measured 0.09 to 0.18 mm.
The length of female Haemonchus measured 15 – 23.8 mm while the width was 0.29 – 0.38 mm (Fig. 2C). The cervical papillae measured 0.24 – 0.42 mm (Fig. 2F), while the esophagus was lengthy and club-shaped, measuring 0.72 – 1.22 mm (Fig. 2G). Female Haemonchus has a slender, spineless, straight, and pointed tail. The characteristic ‘barber pole’ appearance, resulting from the coiled uterus and intestines, was observed. The vulva is located in the posterior portion of the body, approximately 0.68 – 0.82 mm from the tail end, and is covered by a large vulvar flap (Fig. 2E). The anus is located 0.29 – 0.35 mm from the posterior end (Fig. 4D).
Following DNA extraction and COX-1 gene amplification, the PCR products of two Haemonchus isolates from Malakand Division were examined under UV light, producing an amplicon of approximately 666 bp (Fig. 3). The Neighbor-Joining method was utilized to construct a phylogenetic tree comparing the two isolates from the current study with 19 published sequences from various regions, including Pakistan, Bangladesh, Iran, Brazil, USA, New Zealand, Australia, and the UK, as well as out-group nematode species.

Agarose gel electrophoresis showing PCR amplification of COX region of H. contortus.
Phylogenetic analysis revealed two distinct haplotypes in the present study isolates. One isolate (5047657) was grouped with isolates from Pakistan (724353), Bangladesh (LC716224), and Iran (MW051258), suggesting genetic similarity with the isolates from neighboring regions. The other isolate (5047660) formed a separate haplotype (Fig. 4). These findings indicate the presence of two distinct genetic variants of Haemonchus, which may reflect intraspecies variation or localized adaptation within Haemonchus populations in the Malakand division.
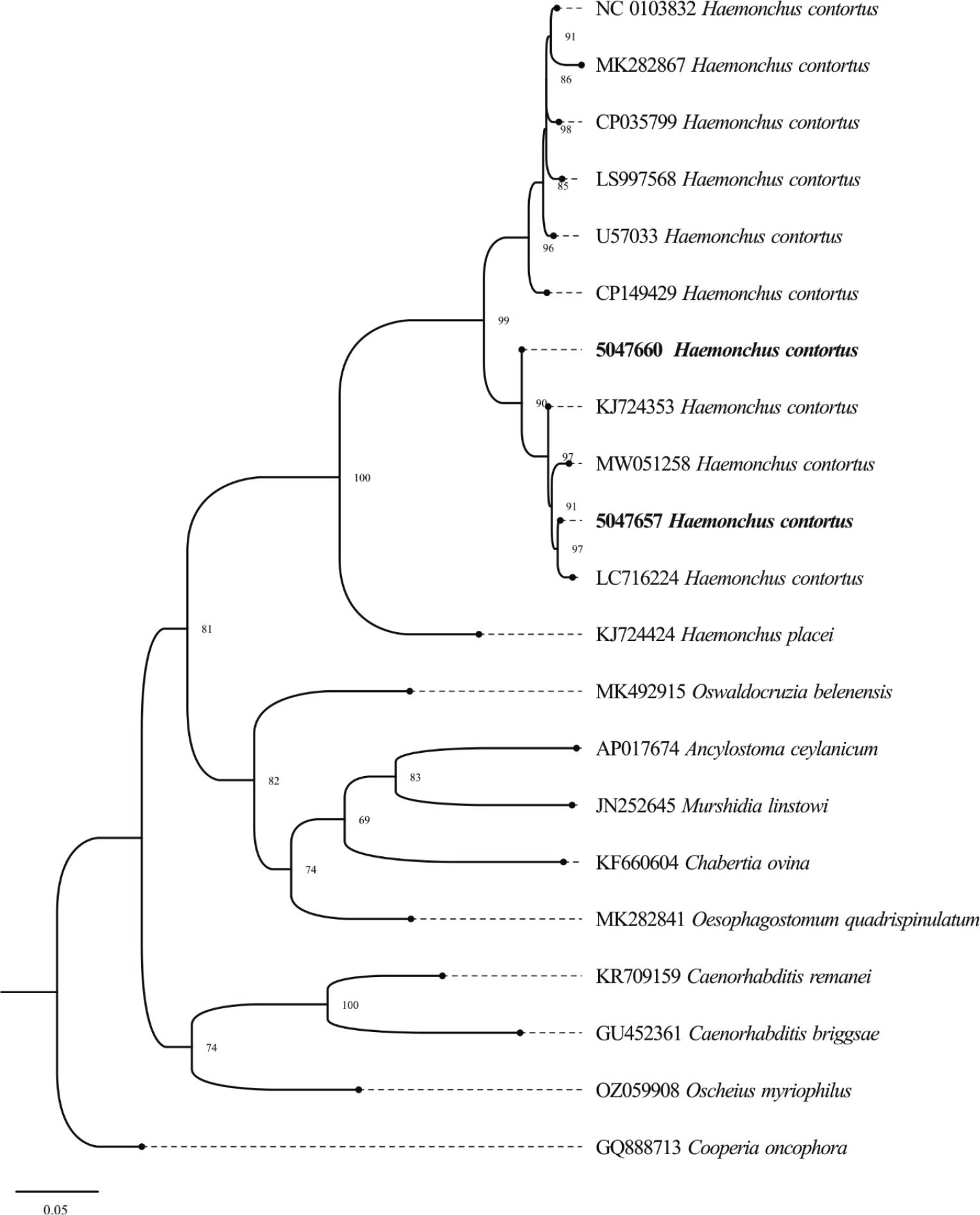
Phylogenetic tree of H. contortus in Goats and Sheep using Cox-1 marker. The constructed tree illustrates the genetic links between the isolates of the current study and reference sequences from other areas. Bootstrap values are given at branch points representing the group’s confidence levels.
Haemonchus, a parasite that affects both the health and production of animals, poses a serious challenge to the livestock industry and results in substantial economic losses (Arsenopoulos et al., 2021). Given the significance of haemonchosis, this study aimed to evaluate the prevalence, risk factors, morphometric characteristics, and phylogenetic analysis of Haemonchus in small ruminants of the Malakand Division. Previous studies were reviewed and summarized to provide a contextual background on the epidemiology of haemonchosis in Pakistan (Table 1). The present study revealed that the prevalence of Haemonchus in small ruminants of Malakand division was 25.17 %, which falls within the broad range reported in earlier studies (6.5 % to 87 %; Table 1). Various factors, such as management strategies, environmental variability, sample size differences, and seasonality in the research region, could be responsible for the observed fluctuation in prevalence. The parasite density may be affected by grazing patterns, socioeconomic factors, and overuse of anthelmintics (Lashari & Tasawar 2011).
The current study found that the overall sex-wise prevalence of haemonchosis in small ruminants was higher in females (26.66 %) compared to males (23.09 %), p>0.05. According to similar studies conducted by Ahmad et al. (2024), Raza et al. (2014), and Brik et al. (2019), females are more prone to parasitism due to reproductive stress and compromised immune systems, which may account for the greater occurrence of haemonchosis. However, the current study contradicts the findings of Rinaldi et al. (2015) and Mohamed et al. (2024), who reported that males had a higher prevalence of Haemonchus than females, suggesting that local factors may also influence the susceptibility.
Age-specific prevalence showed higher infection rates in the oldest (>5 years) (27.98 %) and youngest (<1 year) (25.53 %) age groups. These results align with the findings of Ullah et al. (2013) and Adduci et al. (2022), who also reported higher rates of Haemonchus infection in younger and older age groups. The higher infection rates in older and younger age groups may be attributed to weaker immunity. Targeted control measures for these vulnerable age groups could help reduce the parasite burden.
Risk factor analysis highlighted the grazing system as a significant determinant of infection, with a higher prevalence (27 %) in grazing small ruminants than in stall-fed small ruminants (23 %). Our results align with those of Shahzad (2016). Greater exposure to infectious larvae on contaminated pastures may be the cause of the higher prevalence of haemonchosis in grazing animals.
Similarly, access to contaminated water sources and poor body condition were associated with higher infection rates. The highest infection rate was found in small ruminants with poor body condition (36.19 %) and those drinking from streams and open water sources (28.42 % and 27.48 %, respectively). These results highlight the role of nutrition, water hygiene, and grazing management in controlling haemonchosis. The present study is consistent with earlier findings of Shahzad (2016), Sangma et al. (2012), and Irshad et al. (2023).
In sheep and goats, seasonal prevalence rates of Haemonchus showed that the highest infection rates were observed in the spring (27.59 %) and summer (28.26 %). Monsoon rains, dense vegetation, and frequent temperature changes may be the cause of the increased prevalence of Haemonchosis in small ruminants during these seasons. These findings align with those of Irshad et al. (2023).
According to the study, the prevalence of Haemonchus was significantly higher in untreated small ruminants (29 %) than in treated ones (17 %). These results are in line with those of Raza et al. (2007), Irshad et al. (2023), and Ijaz et al. (2009), who similarly reported lower infestation rates in treated animals. The high prevalence of Haemonchosis in the research area’s animal population could result from negligence and mistreatment.
The eggs of H. contortus in the current study were oval, with an average length of 80.98 μm and a width of 45.0 μm. The findings of the current study are consistent with Soulsby (1965), who found H. contortus eggs to measure around 74×44 μm. The findings of the current study contrast with those of Thienpont (2003) and Inder et al. (2010), who reported larger sizes of H. contortus eggs, measuring 83.7×45.2 μm and 94.7×58.3 μm, respectively. Variations in egg size among these studies may have been due to host species, geographical location, environmental conditions, or the methodology employed for measurement.
The findings of the current study provide an essential understanding of Haemonchus’ morphometric features, which are necessary for parasite identification and classification. Morphometric measurements of H. contortus in the current study showed both consistency and variations with prior studies. The body length of male worms (11.9 – 13.8 mm) was slightly greater than reported by Nath et al. (2021) and Kuchai et al. (2012), although shorter than that documented by Bibi et al. (2017). Male spicule lengths (0.44 – 0.78 mm) were consistent with the findings of Soulsby (1982) and Bibi et al. (2017), although gubernaculum and esophagus lengths were typically shorter than previously documented. The body lengths of females (15 – 23.8 mm) were comparable to the findings of Kuchai et al. (2012) but were shorter than those reported by Bibi et al. (2017). The lengths of the esophagus and the distances from the vulva and anus to the posterior end were also lower than those reported by Nath et al. (2021).
The consistency in morphometric parameters between the current study and prior studies verifies that Haemonchus is accurately identified in small ruminants of the Malakand Division. The minor variations between the present study and previous findings may be due to host-related and geographic factors that influence the morphology of parasites. However, differences have little impact on the overall identification of Haemonchus.
Phylogenetic analysis of Haemonchus isolates of the Malakand Division, using COX-1 gene sequences, revealed genetic variation. Genetic divergence between isolates 5047657 and 5047660, as revealed by phylogenetic analysis, may suggest intraspecies diversity that is influenced by host differences, geographic location, or environmental stress in the Malakand. The same findings have been reported by Yin et al. (2013), Parvin et al. (2024), and Moudgil et al. (2022), who observed that Haemonchus often forms two to three clades based on host-associated and geographic variations.
The primary limitation of this study was time constraints, which resulted in restricted sampling across districts and limited sequencing for genetic diversity analysis. Future studies should focus on expanding sampling and sequencing efforts to understand better the genetic diversity of Haemonchus in the area.
The current study concluded that Haemonchus is prevalent in Pakistan, particularly in the Malakand Division and is significantly associated with health status, prior anthelmintic treatment and host age. Phylogenetic analysis revealed two genetically distinct isolates forming separate clades, indicating genetic diversity within the isolates. This study contributes to a better understanding of epidemiological and genetic patterns of Haemonchus, which may aid future research on parasite management. Future studies should explore additional genetic markers and geographic regions in order to enhance the understanding of Haemonchus diversity and epidemiology.