
Allocreadiidae is a family of trematodes with a cosmopolitan distribution, whose species in their adult stage parasitize mainly freshwater fish, although some parasitize marine fish and other vertebrates in freshwater habitats such as amphibians and snakes, and some species may also be progenetic in arthropods (Caira & Bogéa, 2005). Caira and Bogéa (2005) recognized 15 genera in the family and an additional 15 genera that were controversial. With new revisions on the morphology and the addition of genetic data for phylogenetic comparisons, several of the genera recognized by Caira and Bogéa (2005) have been synonymized or relocated to other families, and some new genera have been described; additionally, Megalogonia Surber, 1928 and Wallinia Pearse, 1920 were placed within the Allocreadiidae (Perez-Ponce de León et al., 2007). Therefore, 18 valid genera are currently recognized in Allocreadiidae (WoRMS 2022; Caira & Bogéa, 2005; Pérez-Ponce de León et al., 2013).
In the present century, four new genera have been described in the Americas: Auriculostoma Scholz, Aguirre-Macedo & Choudhury, 2004; Paracreptotrema Choudhury, Pérez Ponce de León, Brooks & Daverdin, 2006; Paracreptotrematoides Pérez Ponce de León, Pinacho-Pinacho, Mendoza-Garfias, Choudhury & García-Varela, 2016; and Pseudoparacreptotrema Pérez Ponce de León, Pinacho-Pinacho, Mendoza-Garfias, Choudhury & García-Varela, 2016. However, more recently it was found that Auriculostoma is a synonym of Creptotrema (Franceschini et al., 2021). The discovery in recent decades of new genera, their validity, and taxonomic changes, in principle, have been due to the boom in combining DNA sequence analysis with traditional morphological characters. Thanks to the use of molecular phylogenies, it was possible to determine that some morphological and host differences between the species provided strong evidence that they belonged to different genera (e.g. Pseudoparacreptotrema and Paracreptotrematoides from Paracreptotrema), but also made it possible to synonymize seemingly distinct genera (e.g. Auriculostoma with Creptotrema). In the case of this study, trematodes of Allocreadiidae were collected in the intestine of Atherinella alvarezi (Díaz-Pardo) (Atheriniformes), and we immediately noticed morphological differences that we had never observed before in allocreadiids. These differences were complemented with DNA sequences of the 28S and ITS from the ribosomal RNA region (rRNA)and the phylogenies obtained corroborated that the trematodes we found are a new genus and species. Therefore, the objective of this study is the description of this new taxon found in Mexico, to offer a hypothesis about its phylogenetic relationships, and to highlight the importance of integrative taxonomic studies in recognizing and understanding the diversity of trematodes.
As a part of the study of helminth parasites of fishes in the Lacantún River from the Biosphere Reserve of Montes Azules, Chiapas, Mexico, 53 individuals of Atherinella alvarezi were caught using by electrofishing and cast nets in some tributaries and the main channel of the Lacantún River, next to the Chajul Station (16°06’38.4’’ N, 90°56’23.6’’ W) during January 2018. The fish were transported alive to a temporary field necropsy station, where they were necropsied for metazoan parasites by dissecting them and checking all the organs. Particularly, the gastrointestinal tract of each fish was placed in Petri dishes with some 6.5 % saline solution for examination under a stereomicroscope.
The trematodes of the same morphotype were separated into two groups: some were killed and fixed with 4 % hot (nearly boiling) formaldehyde and
stored in vials for morphological study; others were placed directly into vials with absolute ethanol while still alive for DNA extraction and molecular studies.
Of the specimens fixed in 4 % formaldehyde for morphology, 28 were stained with Gomori´s trichrome and Mayer’s paracarmine, cleared in methyl salicylate, and mounted in Canada balsam. These specimens were observed, measured, and drawn using an Olympus DIC Nomarski-BX50 microscope (Olympus Corporation, Tokyo, Japan) equipped with a drawing tube; measurements are presented in micrometers (μm) with the range followed by the mean in parentheses. Two other specimens were processed for scanning electron microscopy (SEM) as follows: specimens were dehydrated in a graded ethanol series (30 %, 50 %, 70 %, 96 % and 100 %), dried at the critical point with carbon dioxide; these specimens were mounted on a metal stub with carbon adhesive tabs, then gold coated, and examined at 10 kV with a Hitachi Stereoscan SU1510 SEM (Hitachi Ltd., Chiyoda, Tokyo, Japan). Specimens were deposited in the Colección Nacional de Helmintos UNAM (CNHE), in the Colección Helmintológica from CINVESTAV Mérida (CHCM), in Natural History Museum of Geneva (MHNG).
To extract genomic DNA, five specimens were digested separately, using the REDExtract-N-Amp Tissue PCR kit (Sigma, St. Louis, Missouri, USA) following the manufacturer’s instructions. Proteinase was denatured at 95º C for 3 min. The ITS (ITS1+5.8S+ITS2) region and 28S rRNA gene (Domains D1+D2+D3) were amplified using polymerase chain reaction (PCR). For ITS, the primers BD1 5′–GTCGTAACAAGGTTTCCGTA–3′, and BD2 5′–TATGCTTAAATTCAGCGGGT–3′ (Luton et al., 1992) were used; for the 28S, the primers 391 5′-AGCGGAGGAAAAGAAACTAA-3′ (Nadler & Hudspeth, 1998) and 536 5’-CAGCTATCCTGAGGGAAAC-3’ (Stock et al., 2001) were used. The PCR was performed in a final volume of 25μl containing 2 μl of genomic DNA, 14.25 μl of ultrapure water, 1 μl of each primer, 2.5 μl of 10x buffer A, 1.5 μl of MgCl2 at 25 mM, 2.5 μl of dNTPs at 2 mM, and 0.25 μl (1 U) of Taq DNA polymerase (Vivantis, Shah Alam, Selangor Darul Ehsan, Malaysia). All PCR reactions were run in an Axygen® MaxyGene™ II thermocycler. Thermal cycling conditions for both ITS and 28S consisted of 95°C for 5 min followed by 35 cycles of 94°C for 1 min, 50 °C for 1 min and 72°C for 1 min, with a final extension at 72 °C for 10 min. Sequencing was performed using the amplification primers plus the internal primers: 503 5’-CCTTGGTCCGTGTTTCAAGACG-3’ (Stock et al., 2001) and 504 5’-CGTCTTGAAACACGGACTAAGG-3’ (García-Varela & Nadler, 2005) for 28S; BD3 5′-GAACATCGACATCTTGAACG-3′ and BD4 5′-ATAAGCCGACCCTCGGC-3′ (Hernández-Mena et al., 2014) for ITS. PCR products were sequenced using an ABI 3500xL Genetic Analyser (Applied Biosystems, Waltham, Massachusetts, USA) in the Laboratorio de Secuenciación Genómica de la Biodiversidad y de la Salud, Instituto de Biología, Universidad Nacional Autónoma de México. Finally, from the resulting sequences, a consensus sequence was obtained for each gene of each extracted specimen using the Geneious Pro 4.8.4 software (Biomatters Ltd., Auckland, New Zealand). Sequences were submitted to GenBank.
Phylogenetic analyzes were performed for the 28S gene, because it is the gene with most sequences for species of Allocreadidae. To build this dataset, most of the sequenced species available in Genbank were added (see Table 1) using the Mesquite 3.62 software (https://www.mesquiteproject.org/). Callodistomidae species were used as outgroup for rooting the trees. The alignment of the sequences was performed with the SATé software, using the following configuration: Aligner = MAFT, Merger = MUSCLE, Tree Estimator = RAXML, Model = GTRCAT, Iteration limit= 200, Tree Return= Best (Liu et al., 2009, 2012). The best alignment was selected for phylogenetic analyses. Nucleotide substitution was estimated with jModelTest v2 (Darriba et al., 2012). Phylogenetic trees were generated with maximum likelihood (ML) and Bayesian inference (BI). For ML analysis, RaxmlGUI v. 1.3 software (Silvestro & Michalak, 2012) was employed to estimate the best tree under the GTR+I+G model and where 1,000 bootstrap repetitions (Bt) were run to estimate the support of the nodes. For the BI analysis we used MrBayes v. 3.2.1 (Ronquist et al., 2012), in which four independent MCMC runs of 20 million generations each were performed. For each run, four chains with a heating parameter value of 0.9 were employed, and the tree topologies were sampled every 1000 generations (printfreq = 1000 sample-freq = 1000 diagnfreq=10,000). Burn-in periods were set to the first 1500 generations. Nodal support was estimated as posterior probability values (PP) and a 50 % majority-rule consensus tree was obtained. The phylogenetic trees obtained were visualized in FigTree v. 1.4.2. Finally, genetic distances were calculated as uncorrected p-distance in MEGA v6 Software (Tamura et al., 2013).
The research followed approval from the Ethics Committee of the Autonomous University of Nuevo Leon (UANL), and after obtaining a permit from the Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT), Mexico (permit numbers: FAUT-017 and SGPA/DGVS/03492).
Family: Allocreadiidae Looss, 1902
Genus: Mesoamericatrema Hernández-Mena, Mendoza-Garfias and Vidal-Martínez
Diagnosis. Body oval, widest at the level of ventral sucker. Tegument smooth, aspinose. Eyespot pigment present and scattered in forebody. Forebody with tegumental papillae on lateral sides.
Trematode species included in phylogenetic analyses with the 28S rRNA gene. The accession number of the Genbank sequences, the host and the locality where the sequenced specimens were collected are shown.
| Species | Author | GenBank | Host | Locality | Sequence Reference |
|---|---|---|---|---|---|
| ALLOCREADIIDAE | Looss, 1902 | ||||
| Acrolichanus auriculatum | (Wedl, 1858) Skvortsov, 1927 | MN524579 | Acipenser schrenckii | Russia | Atopkin et al., 2020 |
| Allocreadium gotoi | (Hasegawa & Ozaki, 1926) Shimazu, 1988 | LC215274 | Misgurnus anguillicaudatus | Japan: Nagano, liyama, Midori | Shimazu, 2017 |
| Allocreadium isoporum | (Looss, 1894) Looss, 1902 | GU462125 | Alburnus alburnus | Russia: Lake Oster, Karelia | Petkeviciüte et al., 2010 |
| Allocreadium lobatum | Wallin, 1909 | EF032693 | Semotilus corporalis | USA | Curran et al., 2006 |
| Allocreadium neotenicum | Peters, 1957 | JX977132 | Hydroporus rufifrons | United Kingdom: Lake District, Cumbria | Bray et al., 2012 |
| Bunodera acerinae | Roitman & Sokolov, 1999 | GU462122 | Gymnocephalus cernua | Russia: River Tvertsa, upper Volga River basin | Petkeviciüte et al., 2010 |
| Bunodera eucaliae | (Miller, 1936) Miller, 1940 | DQ029329 | Culaea inconstans | Canada: Brokenhead River, Manitoba | Choudhury and Leon-Regagnon, 2005 |
| Bunodera inconstans | (Lasee, Font & Sutherland, 1988) Brooks, 1992 | DQ029330 | Culaea inconstans | Canada: Brokenhead River, Manitoba | Choudhury and Leon-Regagnon, 2005 |
| Bunodera luciopercae | (Müller, 1776) Lühe, 1909 | GU462113 | Sphaerium rivicola | Russia: River Tvertsa, upper Volga River basin | Petkeviciüte et al., 2010 |
| Bunodera mediovitellata | Tsimbalyuk & Roitman, 1966 | DQ029332 | Gasterosteus aculeatus | Canada: Little Campbell River, B. C. | Choudhury and Leon-Regagnon, 2005 |
| Bunodera sacculata | Van Cleave & Mueller, 1932 | DQ029333 | Perea fluviatilis | Canada: Lake Sasajewun, Algonquin Park, Ontario | Choudhury and Leon-Regagnon, 2005 |
| Bunodera vytautasi | Atopkin, Sokolov, Shedko, Vainutis & Orlovskaya, 2018 | MG262545 | Pungitius pungitius | Canada: Magadan region, Chernoe Lake | Atopkin et al., 2018 |
| Crepidostomum affine | Tkach, Curran, Bell & Overstreet, 2013 | KF250358 | Hiodon tergisus | USA: Pearl River,Mississippi | Tkach, et a/., 2013 |
| Crepidostomum auritum | (MacCallum, 1919) | KF250357 | Aplodinotus grunniens | USA: Pearl River,Mississippi | Tkach, et a/., 2013 |
| Crepidostomum cooperi | Hopkins, 1932 | DQ029328 | Percopsis omiscomaycus | Canada: Lake Winnipeg, Manitoba | Choudhury and Leon-Regagnon, 2005 |
| Crepidostomum cornutum | (Osborn, 1903) Stafford, 1904 | EF032695 | Lepomis gulosus | USA | Curran et al., 2006 |
| Crepidostomum farionis | (Müller, 1780) Lühe, 1909 | FR821399 | Oncorhynchus masou | Russia | Atopkin and Shedko, 2014 |
| Crepidostomum illinoiense | Faust, 1918 | KF356372 | Hiodon alosoides | USA: Red Lake River,Minnesota | Tkach, et a/., 2013 |
| Crepidostomum metoecus | (Braun, 1900) Braun, 1900 | FR82140 | Salvelinus leucomaensis | Russia | Atopkin and Shedko, 2014 |
| Crepidostomum nemachilus | Krotov, 1959 | FR821408 | Salvelinus leucomaensis | Russia | Atopkin and Shedko, 2014 |
| Crepidostomum oschmarini | Zhokhov & Pugacheva, 1998 | MH159994 | Pisidium casertanum | Lithuania: River Nedzinge | Petkeviciüte et al., 2018 |
| Creptotrema astyanace | (Scholz, Aguirre-Macedo & Choudhury, 2004) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | HQ833707 | Astyanax aeneus | Costa Rica: Guanacaste | Curran etal., 2011 |
| Creptotrema stenopteri | (Mane-Garzon & Gascon, 1973) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | MN822005 | Charax stenopteri | Argentina: Buenos Aires, Punta Lara | Montes et al., 2021 |
| Creptotrema conconae | Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | OK044374 | Imparfinis mirini | Brazil: Upper Parana River basin | Franceschini et al., 2021 |
| Creptotrema creptotrema | Travassos, Artigas & Pereira, 1928 | OK044371 | Megaleporinus elongatus | Brazil: Mogi-Guacu River, Upper Parana River basin | Franceschini et al., 2021 |
| Creptotrema funduli | Mueller, 1934 | JQ425256 | Fundulus notatus | USA: Mississippi, Biloxi River, Harrison County | Curran et al., 2012 |
| Creptotrema guacurarii | (Montes, Barneche, Croci, Balcazar, Almiron, Martorelli & Perez-Ponce de Leon, 2021) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | MN822004 | Characidium heirmostigmata | Argentina: Misiones, Parque Nacional Iguazu | Montes et al., 2021 |
| Creptotrema lobata | (Hernändez-Mena, Lynggaard, Mendoza-Garfias & Perez- Ponce de Leon, 2016) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | KX954170 | Brycon guatemalensis | Mexico: Tenosique, Chiapas | Hernändez-Mena etal., 2016 |
| Creptotrema megacetabularis | Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | OK044375 | Auchenipterus osteomystax | Brazil: Upper Parana River basin | Franceschini et al., 2021 |
| Creptotrema schubarti | Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | OK044373 | Characidium schubarti | Brazil: Upper Parana River basin | Franceschini et al., 2021 |
| Creptotrema tica | (Hernandez Mena, Pinacho-Pinacho, Garcia-Varela, Mendoza- Garfias & Perez-Ponce de Leon, 2018) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | MH997001 | Gymnotus maculosus | Costa Rica: Creek at Pitaya, Guanacaste | Hernändez-Mena etal., 2019 |
| Creptotrema totonacapanensis | (Razo-Mendivil, Mendoza-Garfias, Perez- Ponce de Leon & Rubio-Godoy, 2014) Franceschini, Aguiar, Zago, de Oliveira Fadel Yamada, Bertholdi Ebert & da Silva, 2021 | KF631417 | Astyanax mexicanus | Mexico: Filipinas, Veracruz | Razo-Mendivil et al., 2014b |
| Creptotrematina aguirrepequenoi | Jimenez-Guzman, 1973 | HQ833708 | Astyanax aeneus | Costa Rica: Guanacaste | Curran etal., 2011 |
| Creptotrematina batalhensis | Dias & Abdallah in Alves Dias, Perez-Ponce de Leon, Almeida Camargo, Müller, Silva, Kozlowiski de Azevedo & Abdallah, 2020 | MT512642 | Astyanax lacustris | Brazil: Batalha River | Dias et al., 2020 |
| Margotrema bravoae | Lamothe-Argumedo, 1970 | KT833278 | Allotoca dugesii | Mexico (Central region) | Perez-Ponce de Leon et al., 2016 |
| Margotrema resolanae | Perez-Ponce de Leon, Martinez-Aquino & Mendoza-Garfias, 2013 | KT833271 | Xenotaenia resolanae | Mexico (Central region) | Perez-Ponce de Leon et al., 2016 |
| Megalogonia ictaluri | Surber, 1928 | EF032694 | Ictalurus punctatus | USA: Pearl River, Mississippi | Curran et al., 2006 |
| Mesoamericatrema magnisacculus | This study | OP279638- OP279642 | Atherinella alvarezi | Mexico: Arroyo San Jose, Chiapas | This study |
| Paracreptotrema blancoi | Choudhury, Perez Ponce de Leon, Brooks & Daverdin, 2006 | KT833285 | Priapichthys annectens | Costa Rica: Orosi River | Perez-Ponce de Leon et al., 2016 |
| Paracreptotrema rosenthali | Bautista-Hernandez, Monks, Pulido-Flores & Miranda, 2015 | KT833287 | Xiphophorus malinche | Mexico: Malila River, Hidalgo | Perez-Ponce de Leon et al., 2016 |
| Paracreptotrematoides heterandriae | (Salgado-Maldonado, Caspeta-Mandujano & Vazquez, 2012) Perez Ponce de Leon, Pinacho-Pinacho, Mendoza-Garfias, Choudhury & Garcia-Varela, 2016 | KF697693 | Heterandria bimaculata | Mexico: Agua Bendita, Xico, Veracruz | Razo-Mendivil et al., 2014a |
| Pseudoparacreptotrema axtlaensis | Mendoza-Garfias & Choudhury, 2020 | MT180832 | Agonostomus monticola | Mexico: Axtla de Terrazas, San Luis Potosi | Perez-Ponce de Leon et al., 2020 |
| Pseudoparacreptotrema falciformis | Hernändez-Mena & Pinacho-Pinacho, 2020 | MT180829 | Agonostomus monticola | Mexico: Matias Romero, Oaxaca | Perez-Ponce de Leon et al., 2020 |
| Pseudoparacreptotrema macroacetabulata | Perez Ponce de Leon, Pinacho-Pinacho, Mendoza-Garfias, Choudhury & Garcia-Varela, 2016 | KT833322 | Profundulus kreiseri | Guatemala: Puente Sansare | Perez-Ponce de Leon et al., 2016 |
| Pseudoparacreptotrema pacificum | Sereno-Uribe & Garcia-Varela, 2020 | MT180810 | Agonostomus monticola | Costa Rica: Rio Lajas | Perez-Ponce de Leon et al., 2020 |
| Pseudoparacreptotrema profundulusi | (Salgado-Maldonado, Caspeta-Mandujano & Martinez- Ramirez, 2011) Perez Ponce de Leon, Pinacho-Pinacho, Mendoza-Garfias, Choudhury & Garcia-Varela, 2016 | KT833290 | Profundulus sp. | Mexico: Templo River, San Juan del Rio, Oaxaca | Perez-Ponce de Leon et al., 2016 |
| Wallinia anindoi | Hernändez Mena, Pinacho-Pinacho, Garcia-Varela, Mendoza- Garfias & Perez-Ponce de Leon, 2018 | MH997003 | Astyanax aeneus | Mexico: San Juan del Rio, Oaxaca | Hernändez Mena et al., 2018 |
| Wallinia brasiliensis | Dias, Müller, Almeida, Silva, Azevedo, Perez-Ponce de Leon & Abdallah, 2018 | MH520995 | Astyanax fasciatus | Brazil: Upper Paranä River Basin | Dias et al., 2018 |
| Wallinia caririensis | Silva, Dias, Silva & Yamada, 2020 | MW024866 | Astyanax bimaculatus | Brazil: Batateiras River | da Silva et al., 2021 |
| Wallinia chavarriae | Choudhury, Daverdin & Brooks, 2002 | HC833703 | Astyanax aeneus | Costa Rica: Animas River, Guanacaste | Curran et al., 2011 |
| Wallinia mexicana | Perez-Ponce de Leon, Razo-Mendivil, Mendoza-Garfia, Rubio-Godoy & Choudhury, 2015 | KJ535504 | Astyanax mexicanus | Mexico: Covadonga River, Durango | Perez-Ponce de Leon et al., 2016 |
| CALLODISTOMIDAE | Odhner, 1910 | ||||
| Prosthenhystera caballeroi | Jimenez-Guzmän, 1973 | KM871185 | Astyanax aeneus | Costa Rica: Tampisquto River, Guanacaste | Tkach and Curran, 2015 |
| Prosthenhystera obesa | (Diesing, 1850) Travassos, 1922 | AY222206 | Hoplias sp. | Peru: Itaya River, Iquitos | Olson et al., 2003 |
| Prosthenhystera oonastica | Tkach & Curran, 2015 | KM871181 | Pylodictis olivaris | USA: Pearl River, Mississippi | Tkach and Curran, 2015 |
Oral sucker spherical, subterminal, with oral papillae, 6 or more anterior external, 4 on inner surface (2 near mouth) and 5 posterior external. Oral lappets or lobes lacking. Ventral sucker spherical, muscular, slightly larger than oral sucker. Mouth subterminal. Prepharynx absent. Pharynx rounded, muscular. Esophagus short. Intestinal bifurcation in mid-forebody; caeca terminating at level of posterior testis. Testes two, rounded, entire, tandem, overlapping or slightly separate, in posterior half of hindbody. Cirrus sac very large in proportion to body length, elongate, median, claviform, extending to ovary. Seminal vesicle unipartite, continuous. Pars prostatica ovoid. Ejaculatory duct longer than pars prostatica. Cirrus not observed. Genital atrium indistinct. Genital pore median, between intestinal bifurcation and ventral sucker. Ovary globular, smooth, sinistral, post-equatorial, pre-testicular, posterior to ventral sucker. Seminal receptacle flask-shaped. Mehlis’ gland between ovary and anterior testis. Laurer’s canal opening on dorsal surface of body. Vitelline follicles large, in two ventro-lateral fields not confluent, from intestinal bifurcation to posterior end of body. Uterus not extending posteriorly beyond anterior margin of anterior testis, with few eggs (from 2 to 5). Metraterm similar in length to cirrus sac. Eggs oval and operculate. Excretory vesicle I-shaped, extending to anterior testis. Excretory pore terminal. Parasites in the intestine of atherinopsids (Atheriniformes: Atherinopsidae), in Middle America.
Type species: Mesoamericatrema magnisacculus Hernández-Mena, García-Teh and Caspeta-Mandujano n. sp.
Type host: Atherinella alvarezi (Díaz-Pardo, 1972), Gulf silverside (= Plateadito de Tacotalpa in spanish) (Atheriniformes: Atherinopsidae).
Infection site: Intestine.
Type locality: Arroyo San José, Chiapas, Mexico (16°06’50.0” N, 90°56’03.3” W).
Other localities: Arroyo San Pablo, Chiapas, Mexico (16°06’10.0” N, 91°00’52.2” W).
Etymology: The name of the genus reflects the fact that the type species is distributed in Mexico, in a portion of the cultural region known as “Mesoamerica” that includes from the southern half of Mexico to Nicaragua.
Zoobank Life Science Identifier: urn:lsid:zoobank. org:act:733A83FD-3C04-4304-A1D4-507F2CD12090
Mesoamericatrema n. gen. possesses all the adult characteristics of the family Allocreadiidae as stated by Caira and Bogéa (2005), such as an aspinose tegument, unarmed cirrus (when observable), well developed cirrus sac, lack of an external seminal vesicle, and the general disposition of the vitelline follicles and gonads. The new genus can be easily distinguished from Acrolichanus Ward, 1917, Bunodera Railliet, 1896, Bunoderella Schell, 1964, Crepidostomum Braun, 1900, Creptotrema Travassos, Artigas & Pereira, 1928, Creptotrematina Yamaguti, 1954, Megalogonia Surber, 1928, Paracrepidostomum Lü & Wu, 1996, Rastridostomum Bilqees, Khatoon, Bibi & Mutiur-Rehman, 2007, Trematichtys Vaz, 1932, because Mesoamericatrema n. gen. does not have oral lappets or oral lobes on the oral sucker (Caira & Bogéa, 2005; Franceschini et al., 2021), while the other genera have lobes or lappets protruding from the oral sucker.
On the other hand, Mesoamericatrema n. gen. is easily differentiated from Allocreadium Looss, 1900, Caudouterina Martin, 1966, Margotrema Lamothe-Argumedo, 1970, Paracreptotrema, Paracreptotrematoides, Pseudallocreadium Yamaguti, 1971, Pseudoparacreptotrema, and Wallinia because the new genus has a very elongated cirrus sac that is large in proportion to body size (1: 2.1–2.9) and extends posteriorly beyond the ventral sucker to the ovary, while in the other genera the cirrus sac is small or slightly elongated and does not extend posteriorly beyond the ventral sucker (with the exception of Wallinia, where the sac extends only slightly beyond the ventral sucker). Additionally, the new genus can be distinguished from the others by the combination of some morphological characters, such as the distribution of testes, vitelline follicles, the uterus and the number of eggs: Mesoamericatrema n. gen. presents testes in tandem, non-confluent vitelline follicles extending posteriorly to the posterior end of the body, a pre-testicular uterus, with very few eggs; Margotrema has oblique testes, and non-confluent vitelline follicles that do not reach the testes, post-testicular uterus, with numerous eggs; Caudouterina has oblique testes, vitelline follicles confluent in the anterior region of the ventral sucker and extending posteriorly near the posterior end of body, and a post-testicular uterus with numerous eggs; Wallinia has oblique testes, non-confluent vitelline follicles extending posteriorly between the posterior testis and the end of caeca, and a post-testicular uterus with numerous eggs; Pseudallocreadium and Allocreadium have testes that are oblique or in tandem respectively, vitelline follicles that are confluent in the post-testicular region and extend posteriorly to the posterior end of the body, and a pre-testicular uterus with numerous eggs; Paracreptotrema and Paracreptotrematoides have symmetrical testes, non-confluent vitelline follicles extending posteriorly only to the testes, uterus extending into testicular space, with relative few eggs; finally, Pseudoparacreptotrema has symmetrical or oblique testes, vitelline follicles that are confluent in post-testicular region and testicular region extending posteriorly to the posterior end of the body, uterus extending into the testicular space, and relatively few eggs.
Description (Based on 19 adult specimens from Arroyo San José, and 9 adult specimens from Arroyo San Pablo, Chiapas, Mexico; Figs 1, 2 and 3). Body aspinose, oval, small, 465–710 (574) long, maximum width 155–230 (201) at posterior region of ventral sucker; anterior end rounded. Tegument smooth. Eyespot pigment scattered in forebody at level of pharynx. Forebody, 140–200 (162) long; forebody length to total body length ratio 1: 3.3–3.6 (3.5). Hindbody with ventro-lateral tegumental papillae in the first half of the segment, 232–400 (314) long; forebody length to hindbody length ratio 1:1.6–2.3 (1.9), first half of hindbody with ventro-lateral tegumental papillae. Oral sucker spherical, subterminal, 62–85 (69) long, 67–75 (70) wide, with oral papillae: 6 or more anterior external, 4 on inner surface (2 near mouth) and 5 posterior external (Fig. 2). Oral lappets or lobes lacking (Fig. 2). Mouth subterminal. Ventral sucker spherical with circular opening, larger than oral sucker, muscular, 88–110 (97) long by 85–102 (96) wide. Oral sucker length to ventral sucker length ratio 1: 1.3–1.6 (1.4); Oral sucker width to ventral sucker width radio 1:1.3–1.5 (1.4Prepharynx absent. Pharynx rounded, muscular 30–45 (36) long by 33–40 (36) wide. Esophagus very short, almost imperceptible and narrow. Intestinal bifurcation in mid-forebody, 95–120 (103) from anterior end of body; caeca long, narrow, terminating at level of the posterior testis, 332–462 (380) long, 15–25 (19) wide. Testes two, symmetrical, rounded, smooth, entire, medial, tandem, overlapping or slightly separate, located in posterior half of hindbody. Anterior testis 50–55 (52) long by 45–57 (52) wide. Posterior testis 49–62 (54) long by 49–60 (54) wide, to 65–125 (84) from posterior end of body. Cirrus sac elongate, median, dorsal, claviform, narrower anteriorly and much broader posteriorly, extending from genital pore to anterior half of ovary, 176–332 (224) long, 83–112 (100) wide; cirrus sac length to body length ratio 1: 2.1–2.9 (2.6). Seminal vesicle unipartite, folded, continuous, in posterior part of cirrus-sac, 50–100 (62) long, 50–80 (61) wide. Pars prostatica ovoid 45–62 (52) long by 12–30 (24) wide, surrounded by prostatic cells. Ejaculatory duct longer than pars prostatica, 67–117 (91) long by 10–20 (13) wide. Cirrus not observed. Genital atrium indistinct. Genital pore slit-like, median, between the intestinal bifurcation and the ventral sucker. Ovary globular, unlobed, margin smooth, 52–75 (64) long, 50–75 (61) wide, sinistral, post-equatorial, pre-testicular, between ventral sucker and anterior testis, at 37–78 (56) from posterior margin of ventral sucker. Seminal receptacle flask-shaped, 35–46 (39) long, 55–75 (62) wide, between ovary and anterior testis. Mehlis’ gland composed of scattered gland cells, between ovary and anterior testis. Laurer’s canal indistinct in most specimens, in one specimen opening on dorsal surface of body lateral to the outer border of caecum, at level of ovary. Ootype not observed. Vitellarium formed by large follicles, distributed in two ventro-lateral fields not confluent, extending from intestinal bifurcation level to near posterior extremity of body; follicles overlapping caeca ventrally; vitelline ducts uniting near seminal receptacle to form vitelline reservoir. Uterus intercaecal, forming loops between anterior testis and cirrus sac. Metraterm thick-walled, similar in length to cirrus sac. Eggs oval, operculate, few (2–5), 37–52 (47) long, 32–42 wide (38). Excretory vesicle I-shaped, 140–238 (177) long, 10–18 (14) wide, extending anteriorly to middle of anterior testis. Excretory pore terminal.
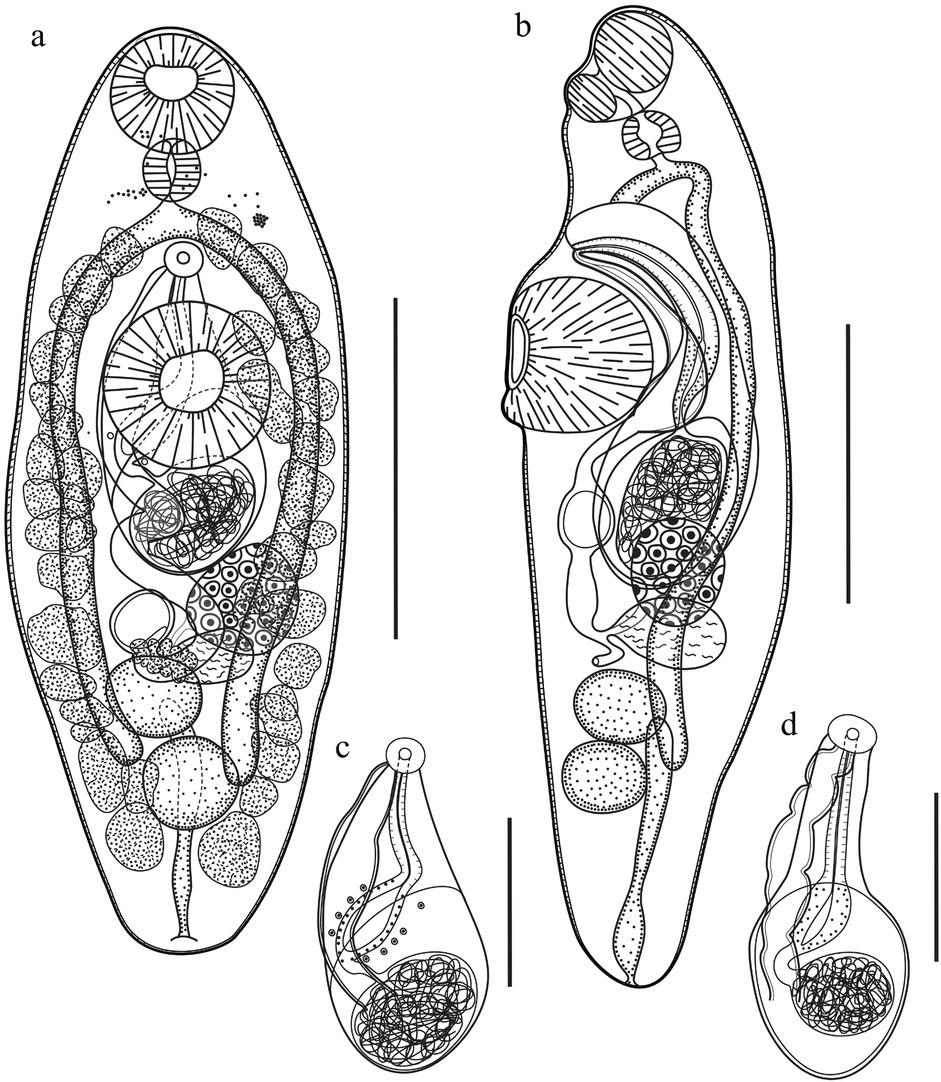
Mesoamericatrema magnisacculus n. sp.: a) Dorso-ventral view of the holotype (scale bar= 200 μm); a) Lateral view of a paratype (scale bar= 200 μm); c) and d) Ventral view of two different male genitalia, showing the cirrus sac containing the seminal vesicle, the pars prostatica, and the ejaculatory duct (scale bar= 100 μm). The seminal vesicle is embedded in a very thick sac.
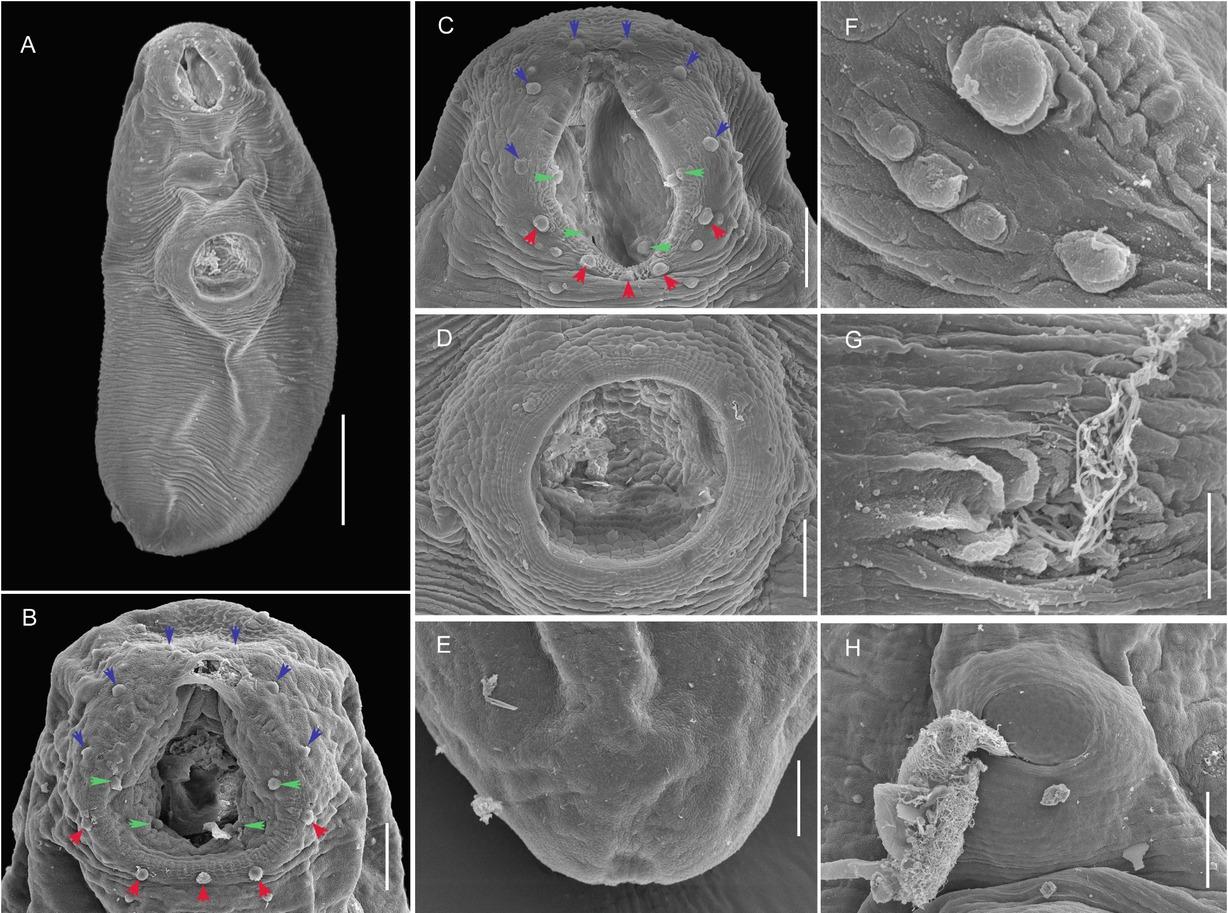
Scanning electron micrographs of Mesoamericatrema magnisacculus n. sp.: A) ventral view of the entire body (scale bar= 100 μm); B) and C) ventral view of two different oral suckers showing the oral papillae (6 external anterior in blue arrows, 4 internal in green arrows and 5 external posterior in red arrows; scale bar= 25 μm); d) ventral view of the ventral sucker (scale bar= 25 μm); e) ventral view of terminal excretory pore at posterior end of hindbody (scale bar= 25 μm); f) close-up of an external oral papilla (scale bar= 5 μm); g) sperm leaving the genital pore (scale bar= 5 μm); h) egg released through the genital pore (scale bar= 25 μm).
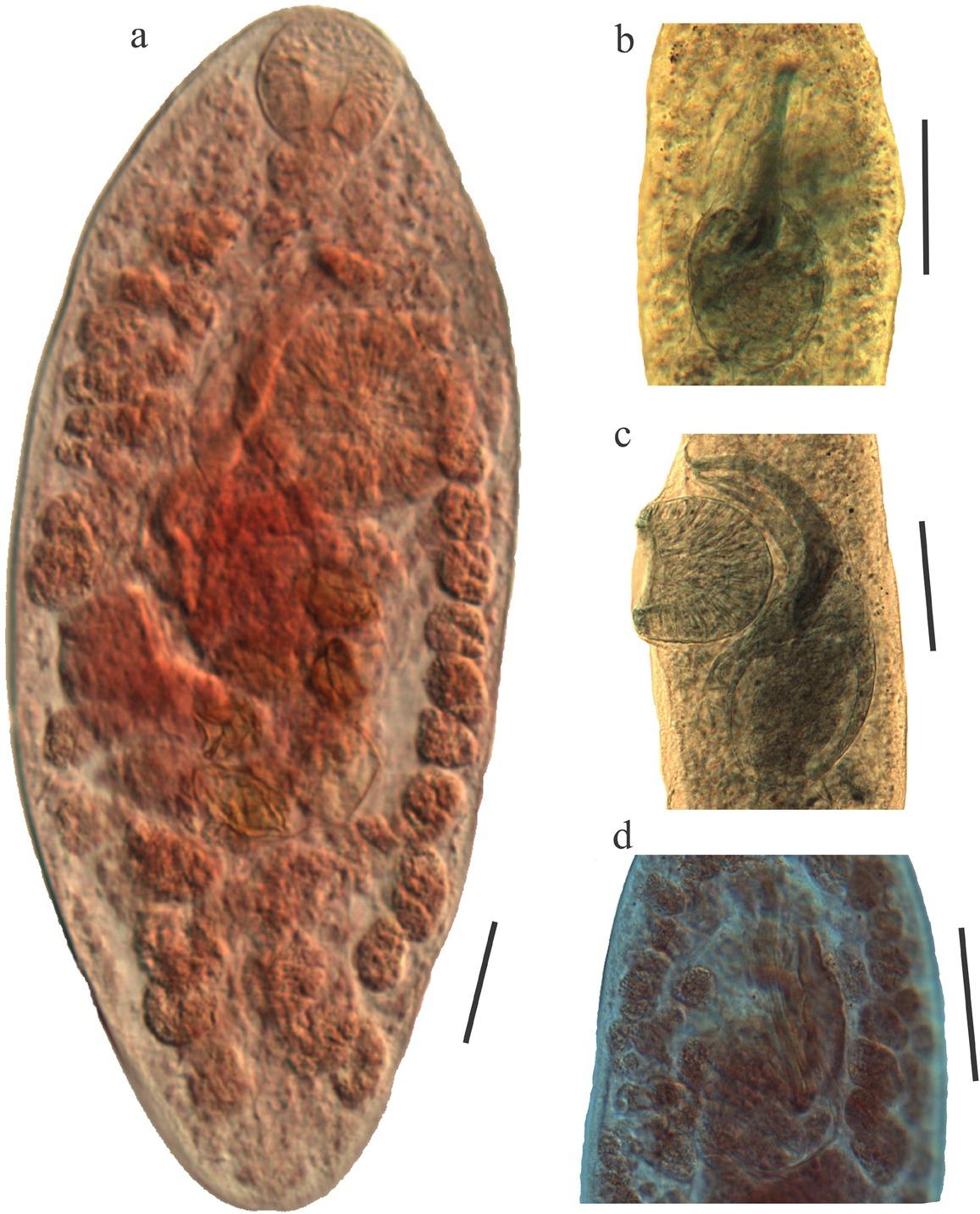
Photographs where the size of the cirrus bag is observed with respect to the size of the body. The length of the ejaculatory duct, the pars prostatica and the seminal vesicle, which is surrounded by a thick wall of the sac, are also observed; a), b) and c) paratypes stained with Mayer’s paracarmine; d) paratype stained with Gomori’s trichrome. Scale bar= 50 μm.
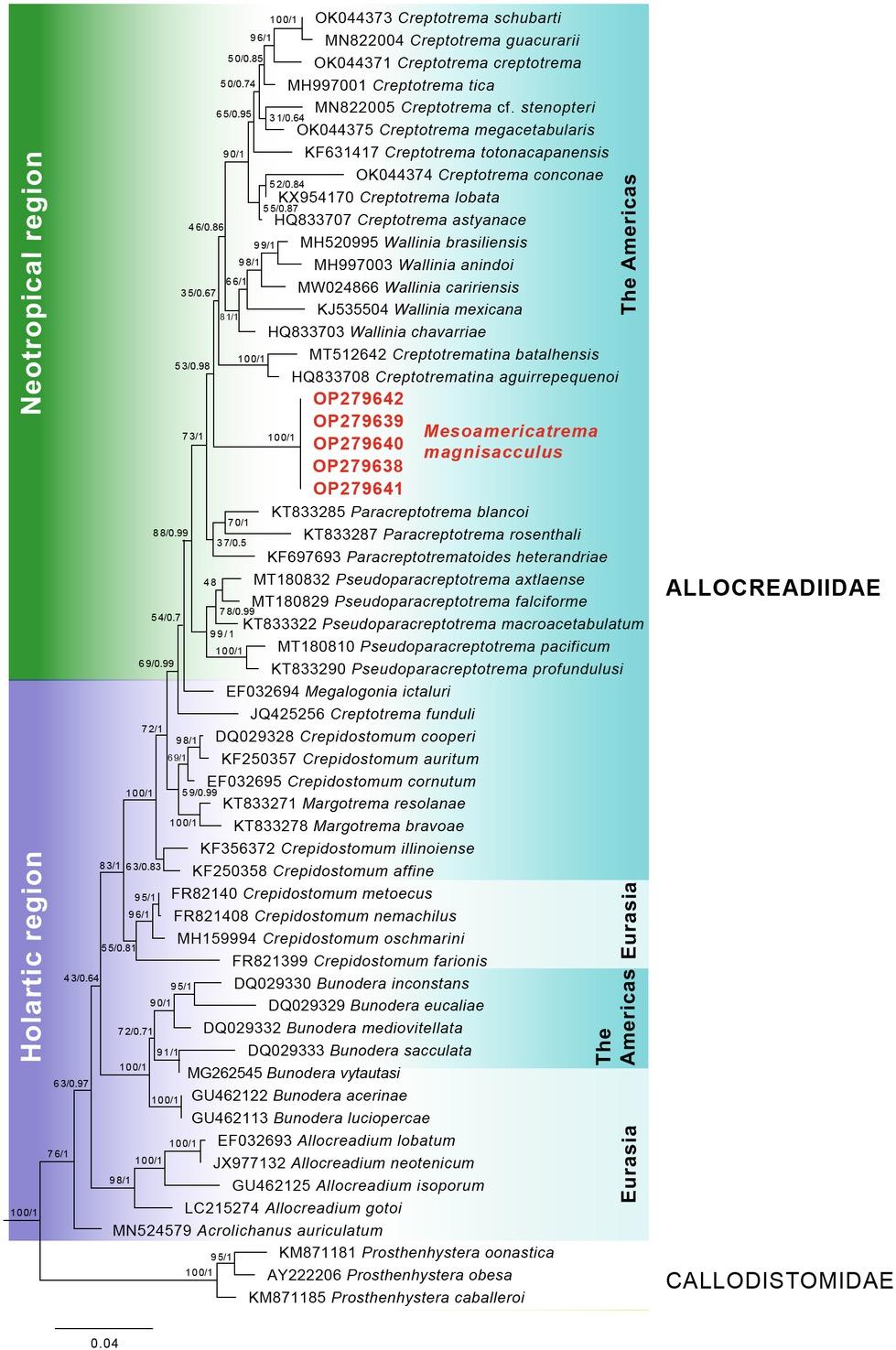
Maximum Likelihood phylogenetic tree of the 28S gene for Allocreadiidae species, showing the position of the new genus that is highlighted in bold red font (–ln= 9888.861897). Numbers near internal nodes indicate bootstrap support and posterior probabilities values.
Type host: Atherinella alvarezi (Díaz-Pardo, 1972), Gulf silverside (= Plateadito de Tacotalpa in spanish) (Atheriniformes: Atherinopsidae).
Type locality: Arroyo San José, Chiapas, México (16°06’50.0” N, 90°56’03.3” W).
Other localities: Arroyo San Pablo, Chiapas, Mexico (16°06’10.0” N, 91°00’52.2” W).
Type material: Holotype CNHE 11671; Paratypes CNHE 11672 (n=5) and CNHE 11663 (n=5); Paratypes CHCM 670 (n=4) and CHCM 671 (n=3); Paratypes MHNG PLAT-0144002 (n=4).
Representative DNA sequences: 28S rRNA (OP279638 – OP279642); ITS rRNA (OP279696 – OP279697).
Etymology: The specific name reflects the fact that this species has a cirrus sac that is huge in proportion to its body length.
Zoobank Life Science Identifier: urn:lsid:zoobank.org:act:0520E4B6-B22F-433E-AF28-C6E32A4D84A9
Phylogenetically, the new species is related to species of genera that are distributed in the Neotropical region: Creptotrema, Creptotrematina, Paracreptotrema, Paracreptotrematoides, Pseudoparacreptotrema, and Wallinia (see below for results of the phylogenetic analyses). Mesoamericatrema magnisacculus n. sp. is easily distinguishable from all species of Creptotrema and Creptotrematina, and from some species of Pseudoparacreptotrema (i. e. P. falciformis Hernández-Mena & Pinacho-Pinacho, 2020 and P. axtlaensis Mendoza-Garfias & Choudhury, 2020) because the new species lacks lappets or lobes protruding from the oral sucker.
The new species is distinguished from all species of Wallinia, Paracreptotrema, Paracreptotrematoides, and from the remaining species of Pseudoparacreptotrema, because M. magnisacculus n. sp. has a very large cirrus sac (1: 2.1–2.9) that posteriorly surpasses the ventral sucker and extends to the ovary, whereas in Wallinia species it is
is moderately elongated and only slightly extends past the ventral sucker, and in the species of Paracreptotrema, Paracreptotrematoides and Pseudoparacreptotrema is short and never extends beyond the posterior level of the ventral sucker. Additionally, the new species has testes in tandem and a pre-testicular uterus with very few eggs; Wallinia species have oblique testes and a post-testicular uterus with numerous eggs; Paracreptotrema and Paracreptotrematoides species have symmetrical testes and a uterus that contains relatively few eggs and extends into the testicular space; and Pseudoparacreptotrema species have symmetrical or oblique testes and uterus that also contains relatively few eggs and extends into the testicular space. Furthermore, M. magnisacculus n. sp. possesses non-confluent vitelline follicles while Pseudoparacreptotrema species possess confluent vitelline follicles in the post- testicular and testicular regions.
The 28S data set was formed of a matrix of 51 species in 13 genera of Allocreadiidae (see Table 1). The length of the sequences in the aligned matrix was 1500 bp. The substitution model was GTR+G+I and the nucleotide frequencies were: A= 0.212, C= 0.214, G=0.318, T=0.256. The ML tree value was –ln= 9888.861897. Both the ML tree and the BI consensus tree presented practically the same topology. Regarding the phylogenetic relationships, all genera, except for Crepidostomum and Creptotrema, were monophyletic. In the case of Creptotrema, previous studies have already suggested that C. funduli, due to its phylogenetic position and geographical distribution, is a different genus and that this species needs a new morphological and taxonomic review (Pérez-Ponce de Léon et al. 2020); therefore, Creptotema sensu stricto included the rest of the species that formed a monophyletic group. In the tree, it was observed that the phylogenetic relationships of the species were consistent with the regions where they are distributed, i.e. the Holarctic and Neotropical regions. The species of the Neotropical region formed a monophyletic clade that was made up of seven genera with high support values (Bt=88; PP=0.99), where the new genus also nested: Creptotrema, Creptotrematina, Mesoamericatrema n. gen., Paracreptotrema, Paracreptotrematoides, Pseudoparacreptotrema and Wallinia. Mesoamericatrema, was placed as the sister group of the subclade (Creptotrematina + (Wallinia + Creptotrema)) (Bt=53; PP=0.98). The genetic distances in the 28S rDNA between Mesoamericatrema magnisacculus n. sp. and the species of the genera of the sister subclade were variable: from 4.59 to 5.46 % with Creptotrematina species, from 3.94 to 6.24 % with Wallinia species and from 3.75 to 6.42 % with Creptotrema species (Table 2). On the other hand, the genetic distances in the ITS gene between Mesoamericatrema magnisacculus n. sp. and species of the genera of the sister subclade were also variable, varying from 4.81 to 5.65 % with Creptotrematina species, 5.12 % with Wallinia chavarriae and from 4.40 to 4.94 % with Creptotrema species (Table 2).
Genetic distances between Mesoamericatrema magnisacculus n. gen. n. sp. and the Allocreadiidae species of the Neotropical clade in the 28S and ITS ribosomal genes (na= sequences not available in Genbank).
| Species | Mesoamericatrema magnisacculus | |
|---|---|---|
| 28S | ITS | |
| Creptotrema schubarti | 5.63 | na |
| Creptotrema guacurarii | 5.37 | na |
| Creptotrema creptotrema | 5.53 | na |
| Creptotrema tica | 3.75 | na |
| Creptotrema cf. stenopteri | 5.01 | na |
| Creptotrema megacetabularis | 4.71 | na |
| Creptotrema totonacapanensis | 3.83 | 4.94 |
| Creptotrema conconae | 6.42 | na |
| Creptotrema lobata | 3.75 | 4.63 |
| Creptotrema astyanace | 3.79 | 4.40 |
| Wallinia brasiliensis | 5.20 | na |
| Wallinia anindoi | 5.43 | na |
| Wallinia caririensis | 6.24 | na |
| Wallinia mexicana | 5.76 | na |
| Wallinia chavarriae | 3.94 | 5.12 |
| Creptotrematina batalhensis | 5.46 | 4.81 |
| Creptotrematina aguirrepequenoi | 4.59 | 5.65 |
| Paracreptotrema blancoi | 3.91 | 7.29 |
| Paracreptotrema rosenthali | 5.69 | 8.27 |
| Paracreptotrematoides heterandriae | 4.62 | 4.34 |
| Pseudoparacreptotrema axtlaense | 5.03 | na |
| Pseudoparacreptotrema falciforme | 4.86 | na |
| Pseudoparacreptotrema macroacetabulatum | 3.93 | 7.56 |
| Pseudoparacreptotrema pacificum | 5.61 | na |
| Pseudoparacreptotrema profundulusi | 5.29 | 7.56 |
The results of our study revealed the existence of a new genus of Allocreadiidae that parasitizes the fish Atherinella alvarezi. The discovery of the new genus named Mesoamericatrema n. gen. was based on the comparative study of internal and external morphological characters (using SEM), on the host association, and on the phylogenetic analysis of the 28S region (domains D1, D2, and D3) of the ribosomal RNA unit. Morphologically, the new genus presents a very large cirrus sac in proportion to the length of its body (1: 2.1–2.9), a characteristic that easily differentiates it from the other genera of Allocreadiidae, since generally the species of the other genera have a cirrus sac that does not extend beyond the posterior margin of the ventral sucker, and when it does, the sac is not as large in proportion to the length of the body (Scholz et al. 2004; Caira & Bogéa, 2005; Choudhury et al., 2006; Pérez Ponce de León et al., 2016; Franceschini et al., 2021).
Scanning Electron Microscopy (SEM) images show that the external surface of the body of Mesoamericatrema n. gen. does not have spines, but it does have oral and ventro-lateral tegumental papillae, features it shares with other species of allocreadiids. In particular, the number of oral papillae can be variable, however, the papillae that are constant in number are the internal papillae close to the mouth (4) and the posterior external papillae (5). This peculiarity of the number of internal and posterior oral papillae had already been observed in other genera such as Creptotrema (Razo-Mendivil et al., 2014; Hernández-Mena et al., 2016; 2019; Montes et al., 2021), Paracreptotrema (Choudhury et al., 2006; Perez-Ponce de León et al., 2016), Paracreptotrematoides (Perez-Ponce de León et al., 2016), Pseudoparacreptotrema (Perez-Ponce de León et al., 2016; 2020) and Wallinia (Hernández-Mena et al., 2019), which usually also have 4 internal papillae and 5 external posterior papillae, which suggests that so this character may be associated with this group of allocreadiids and may even be a synapomorphy for this large Neotropical clade. As a result, we suggest that it be included in the general diagnosis of these aforementioned genera. Unfortunately, not all recent descriptions of new species have SEM images, which makes it difficult to know if the number of oral papillae is a constant character in all species, so we propose that future descriptions include SEM observations to document the distribution of tegumental papillae and other surface features of these trematodes.
Mesoamericatrema n. gen. was nested within a clade that contains species and genera that geographically have a Neotropical and Middle American distribution, which extends from Argentina and Brazil to Mexico, passing through Central America. These genera are Creptotrema, Creptotrematina, Paracreptotrema, Paracreptotrematoides, Pseudoparacreptotrema, and Wallinia. More specifically, the new genus was grouped as the sibling taxon of the clade formed by Creptotrema + Creptotrematina + Wallinia with low bootstrap values but high posterior probabilities in clade support. The composition of the genera that make up the clade of Neotropical allocreadiids had also been previously reported by other authors (Pérez-Ponce de León et al., 2020; Franceschini et al., 2021), who mentioned that the grouping of these trematodes is related to their hosts which are freshwater fishes of Neotropical origin.
Species of all Neotropical genera have been recorded in Mexico, but other genera of allocreadiids have also been previously recorded, such as Allocreadium Looss, 1900, Crepidostomum Braun, 1900, Megalogonia Surber, 1928 and Margotrema Lamothe-Argumedo, 1970 (Perez-Ponce de León et al., 2007; Galaviz-Silva et al., 2013), which have rather a Neartic affinity, so that Mexico is clearly a country where historical geological processes have favored the coexistence of parasites and hosts (and other animals) with different biogeographical affinities (Aguilar-Aguilar et al., 2003; Miller et al., 2005; Pérez-Ponce de León & Choudhury, 2005; Chakrabarty & Albert, 2011; Choudhury et al., 2017; Morrone, 2014; 2020). The hydrological region and the host where the new genus was found have a Neotropical biogeographical affinity, so we expected that Mesoamericatrema n. gen. would nest in the clade of Neotropical allocreadiids instead of grouping with Neartic allocreadiids, a hypothesis that has been supported by the result of phylogenetic analyses. Therefore, the phylogenetic position of Mesoamericatrema n. gen. does correspond to the host and regional affinities of the Neotropical allocreadiids and increases the diversity of genera that have this affinity.
Neotropical allocreadiids parasitize freshwater fish of Neotropical origin (Albert & Reis, 2011) of the following orders (fig. 5): Creptotrema is in Characiformes, Gymnotiformes and Siluriformes; Creptotrematina and Wallinia have only been recorded in Characiformes; Paracreptotrema and Paracreptotrematoides have been found in Cyprinodontiformes; and Pseudoparacreptotrema has been found in Cyprinodontiformes and Mugiliformes. In this study, we found Mesoamericatrema magnisacculus n. sp. as an adult parasite of Atherinella alvarezi, a species of Atheriniformes endemic to the region (Albert & Reis, 2011). Previously, Salgado-Maldonado et al. (2014) had already reported an allocreadiid in A. alvarezi, which they only identified as “Allocreadidae gen. sp. b”. A review of this material allowed us to conclude that it is the taxon that we describe in this study. Thus, this is the first genus and species of allocreadiid that has an affinity for this group of fish. The host affinity of the new genus is consistent with the other genera with which it is phylogenetically grouped, and while their respective hosts do not have a common phylogenetic origin, all of them have evolved in the same biogeographical region. The fact that all Neotropical allocreadiids do not necessarily share the same group of fish (genus, family, or order), may imply that host-switching events have occurred during the evolution of this group of parasites due to the possible sympatry of the different groups of fish that have inhabited the same biogeographical zones.
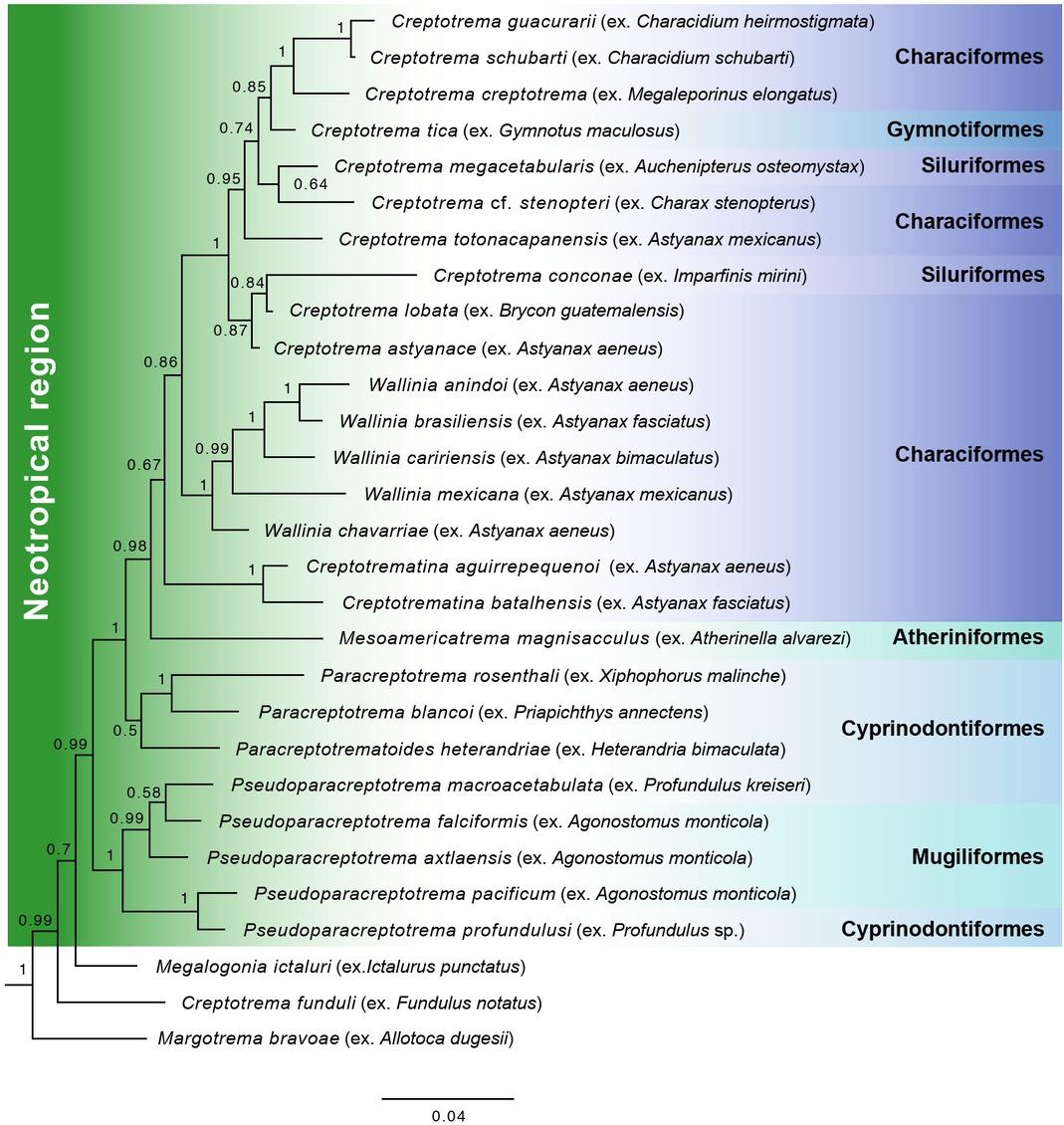
Phylogenetic tree of the species of Allocreadiidae with Neotropical distribution, indicating their definitive hosts and the families of fish to which they belong with Neotropical affinity.
The following parasites have previously been recorded in A. alvarezi: the trematodes “Allocreadidae gen. sp. b”, and metacercariae of Posthodiplostomum minimum MacCallum, 1921 and Centrocestus formosanus Nishigori, 1924; the acanthocephalan Octospiniferoides chandleri Bullock, 1957; and the nematodes Spinitectus osorioi Choudhury and Pérez-Ponce de León, 2001, Rhabdochona sp., Contracaecum sp. and Spiroxys sp. (Moravec et al., 2010; Salgado-Maldonado et al., 2011; 2014). M. magnisacculus n. sp. represents the first species of trematode recorded to date that completes its life cycle in the intestine of A. alvarezi.
Mesoamericatrema n. gen. is the fifth genus of Allocreadiidae to be described in the Americas since 2004 and the third that has been discovered in Mexico since 2016, so it is important to mention that the discovery of this new genus increases parasite diversity both locally and regionally, providing support to the hypothesis that Middle America is indeed a very important region where notable speciation events have taken place in freshwater fish parasites (Choudhury et al., 2016, 2017).