
Charles Darwin (1) closed On the Origin of Species with this famous passage:
There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one; and that, whilst this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.
Darwin's ‘endless forms most beautiful’ was his testimony to the creative power of natural selection. Creativity, by definition, implies novelty: bringing into existence something which had never before existed. Novelty constitutes the grist for the mill of natural selection to refine those endless forms most beautiful.
To Darwin, the source of that novelty was simple variation of form, which conferred on some forms advantage over others in the ‘struggle for existence’. Combined with Alfred Russell Wallace's commentary on the ‘tendency of varieties to depart indefinitely from their original type’ (2), the Darwin–Wallace theory provided the first coherent naturalistic explanation for the generation of life's remarkable diversity and seemingly endless ingenuity. No longer need life's endless forms most beautiful be taken as evidence of the creative power of an omnipresent deity. It now could be explained as a natural phenomenon, like the shaping of landscapes.
Both Darwin and Wallace had trouble sticking to their naturalistic programme, though. The stumbling block for Darwin was heritability of adaptation. A well-adapted creature would fare better in the struggle for existence than a poorly-adapted one. For natural selection to generate those endless forms most beautiful, adaptation had to be heritable. Darwin's abortive answer to this problem was his Lamarckian ‘mad dream’ of pangenesis. Wallace, for his part, carved out an exception for humans, which he regarded as having somehow risen above the animal kingdom whence humans arose.
More challenges confronted the Darwinian idea in the early 20th century. The rise of Mendelian genetics and Morgan's evolutionary doctrine of mutationism threatened to upend the entire Darwinian programme. Ronald Fisher's reconciliation of natural selection with Mendelism breathed new life into the Darwinian idea, but with Darwinism now recast in radically new form: Neodarwinism. Where Darwin looked to selection of favored races as the principal driver of evolution, Neodarwinism looked to selection of favored genes, and mutation as the source of evolutionary novelty (3).
Following the Neodarwinist synthesis, the mid to late 20th century was consumed by the quest for the material nature of the gene. The stage was set by the elucidation of the structure of DNA, and Francis Crick's statement of his Central Dogma of Molecular Biology (Figure 1). The central dogma aligned closely with the Neodarwinian idea of gene selection and mutation as the source of evolutionary novelty. New function, in Crick's dogma, could only arise from new hereditary memory: mutation.
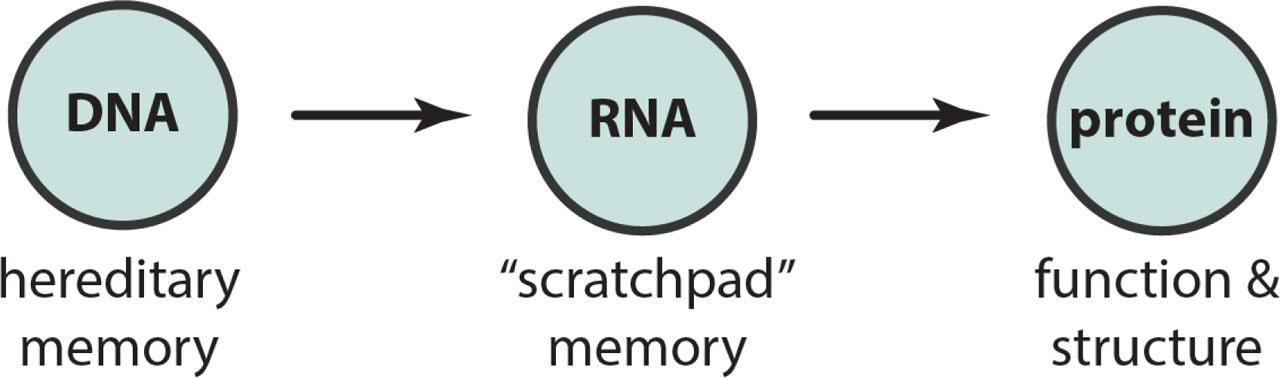
The central dogma of molecular biology.
The central dogma's simplistic picture of the translation of DNA sequence code into coherent function has mostly been undermined by our growing knowledge of the complexity of the genome and its expression. Epigenetic mechanisms have introduced syntax, grammar, post-transcriptional editing, physiological feedbacks, and other nuances, which have blurred the central dogma's neat distinctions between DNA, transcribed RNA, and translation into protein, structure, and function (Figure 2) (4). Not only has the translation of sequence code into coherent function become extraordinarily rich and nuanced, the heritability of adaptation, and even Darwin's mad dream of pangenesis has taken on new life, through epigenetic markers transmitted through gametes now taking on the role of Darwin's gemmules (5).
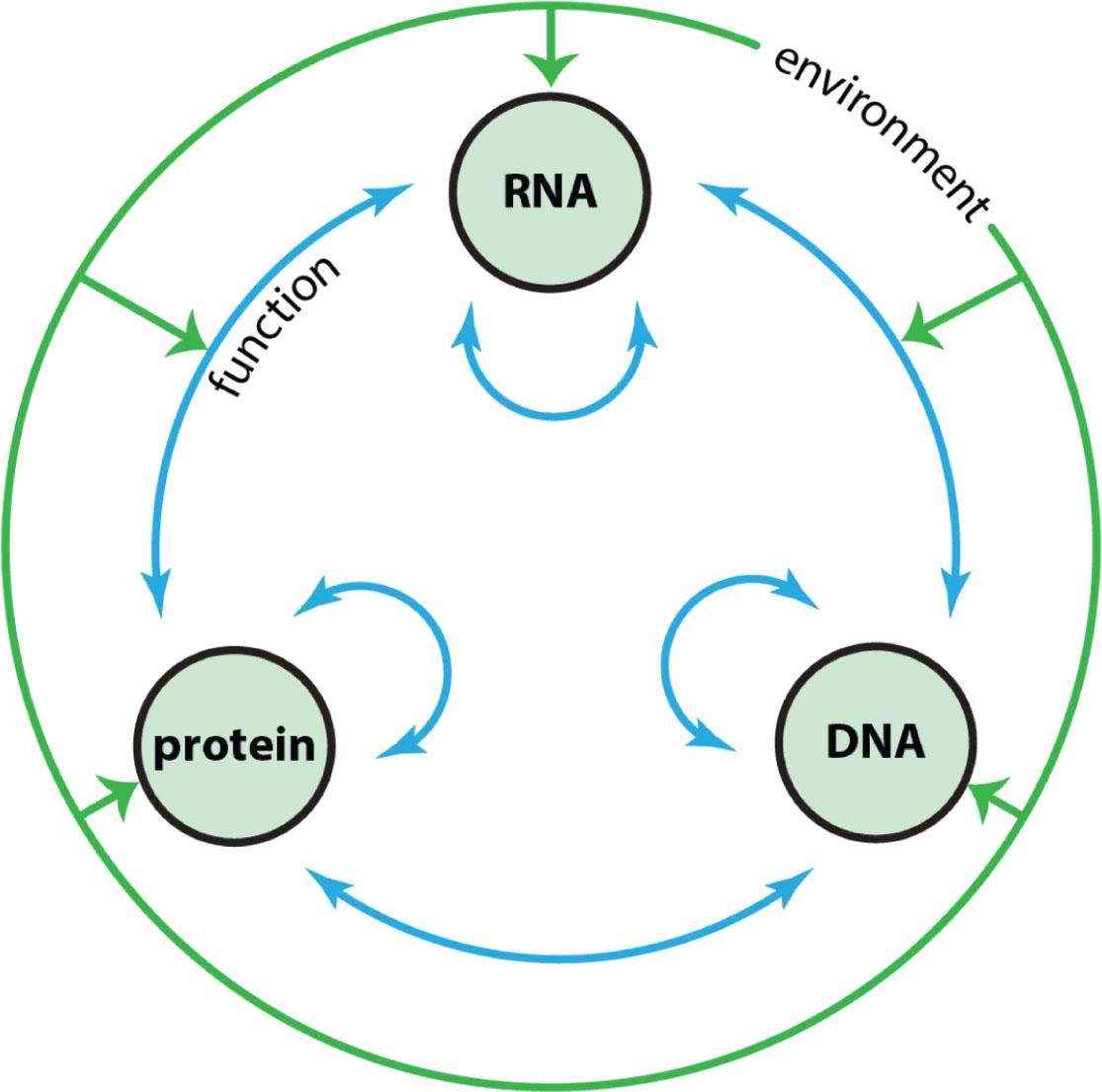
The blurred lines of heredity and function. All components of the central dogma are embedded in multiple feedbacks which blur the distinctions inherent in the central dogma. Among these include direct influence of the environment on all of the ‘-omes’ of cell function – genome, transcriptome, and proteome.
Along with these discoveries has come the opportunity to rethink the nature of evolutionary novelty. The distinction of genotype from phenotype, for example, has driven a conceptual wedge through the organism. Barbara McClintock's characterization of the genome as an ‘organ of the cell’ undermines that division: genes are now participants in coherent function rather than determinants of function, which folds the genotype and phenotype back together into a unitary whole (6). The distinction of genotype and phenotype dissolves, revealed as an artifice which exists primarily to sustain dogmas of gene selectionism and gene determinism. With that has come a host of new questions and ways of looking at the phenomenon of evolutionary novelty, the nature of the organism and of hereditary memory, and the primacy of the Darwinian idea.
‘Organism’ is a maladroit word. Grammatically, ‘organism’ parses as a noun, an object. In reality, ‘organism’ is more a verb, specifically the manifestation of a thermodynamic process. The living organism is a highly ordered, unstable, and highly improbable transient assemblage of matter. Cast in the language of thermodynamics, the living organism is a highly specified low entropy disequilibrium. As all low-entropy systems do, the organism tends spontaneously to degrade to thermodynamic equilibrium. The disequilibrium that is the organism persists nevertheless because it does work to create new specified orderliness as fast as it degrades to disorder (Figure 3). The work rate required to do this is the metabolic rate, expressed in watts. When there is no metabolic rate, the orderly assemblage of matter that the organism comprises degrades to disorderly equilibrium, which is death.
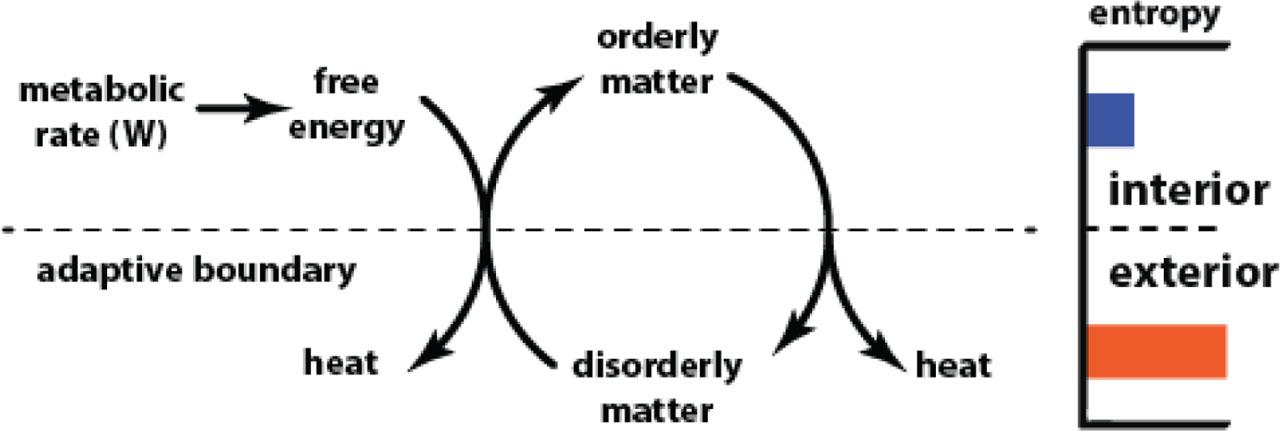
The persistent dynamic disequilibrium. Free energy drives a movement of disorderly matter across an adaptive boundary and does work to organize it into the specified dynamic disequilibrium of the organism. At the same time, the orderly organism degrades spontaneously to disorder.
The process that is the living organism nevertheless has an objective appearance, that is to say, it appears to us to be an object, because the organism is bounded by a skin, integument, membrane, or built structure. In the thermodynamic organism, these constitute adaptive boundaries, which manage the flows of matter and energy that sustain the open thermodynamic system of the organism (Figure 3).
The fundamental adaptive boundary is the cell membrane, which sequesters an intracellular environment from an ambient extracellular environment, and which controls flows of matter and energy across it. Adaptive boundaries can build, nest and interact, however. Coalitions of cells can form sheet-like epithelia, which divide environments on a larger scale, one environment sequestered within the epithelial sheet from the other environment without. The earthworm nephridium, which manages the flow of water and solutes between the earthworm body and its environment, illustrates this concept well (Figure 4).
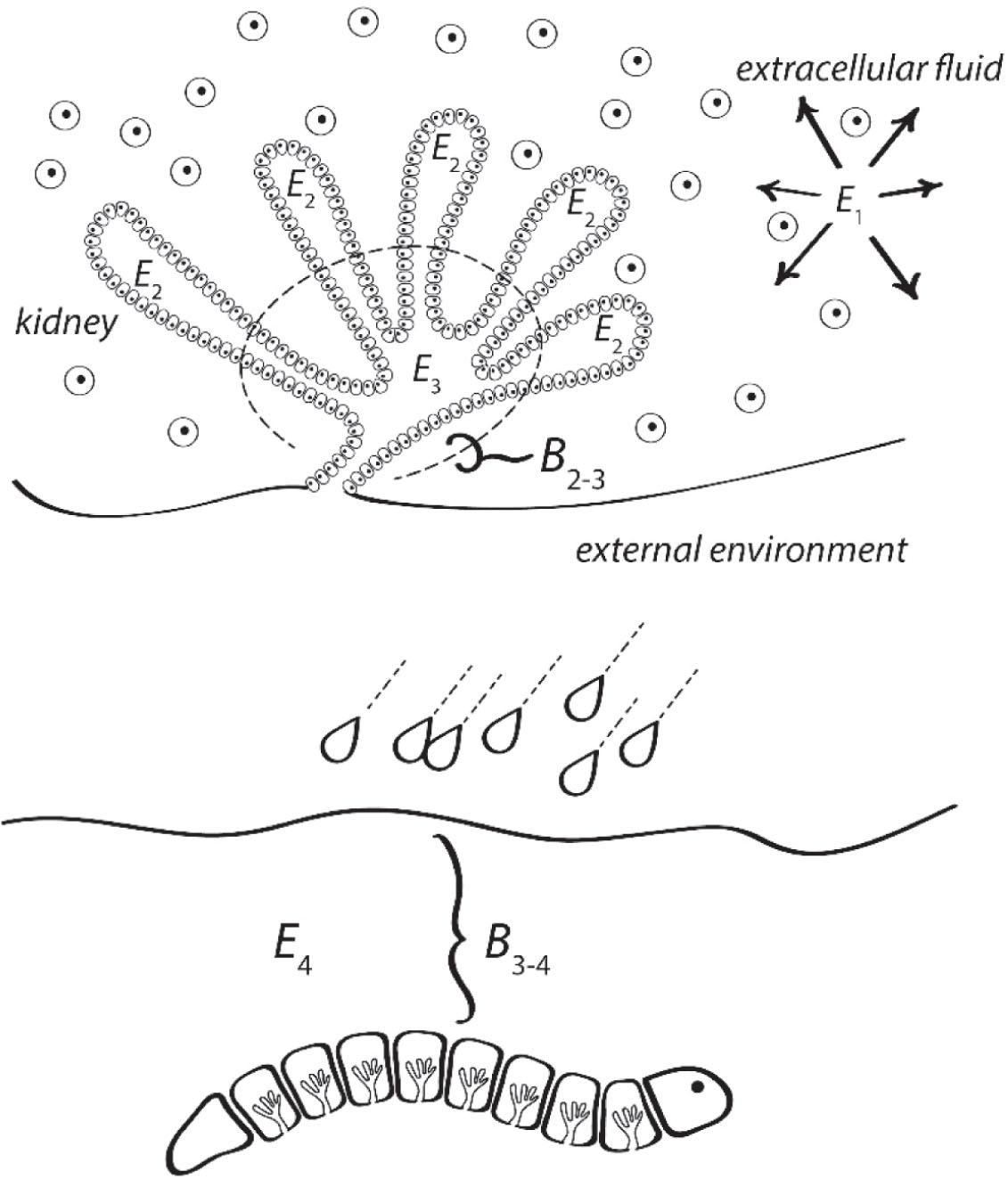
The nested adaptive boundaries and segregation of environments that comprise the earthworm and management of water and solute balance.
The nephridium manages the earthworm's internal water balance in the face of variation in the external environment's water availability. It does so by creating a hierarchy of nested sub-environments, each delimited by an adaptive boundary. The cells living within the worm's coelomic fluid are bounded by the cell membrane (designated B0–1), dividing the intracellular environment (designated E0) from the environment of the extracellular fluid (designated E1). The nephridium epithelium itself is structured to form additional sub-environments within the nephridial tubules (E2), which together create a new environment (E3) within the nephridium bladder. The ultimate segregation of environments occurs at the integument, which is the interface of the earthworm organism and the soil environment in which it lives (E4). Each sequestration of environments (E0, E1, E2, E3, E4) is through a hierarchy of nested adaptive boundaries (B0–1, B1–2, B2–3, B3–4).
The whole organism consists of elaborate networks of such nested adaptive boundaries, which accounts for the hierarchical scales of organization typical of the animal body. Adaptive boundaries can also extend beyond the conventional organism to include animal-built structures, like the soil environment constructed by earthworm populations, nests of social insect colonies, and others. There is no limit to the nesting and layering of adaptive boundaries, save for the biosphere itself (7, 8). The complexity of nested adaptive boundaries does not negate the organism's fundamental nature as an open thermodynamic system. It does, however, complicate the nature of the organism.
The organism as a thermodynamic disequilibrium managed by adaptive boundaries can be conceptualized in different ways. Figure 3 represents the organism as an open system with respect to energy, but a closed-loop system with respect to matter (10). As long as the organism lives, matter continually cycles across the adaptive boundary, with a stream of high-entropy matter drawn across the adaptive boundary as work is done to organize it into the transient low-entropy stream of matter that constitutes the organism. In accordance with the second law, this low entropy state degrades spontaneously to equilibrium as it returns to the environment whence it came, until it is cycled back through the adaptive boundary's thermodynamic engine.
Applying the conservation of mass principle to this model yields a counterintuitive conclusion: that physiological function cannot be confined to a ‘living’ environment within an adaptive boundary. Mass brought into a cell, for example, is also mass taken from the environment on the other side of the cell membrane. The action of an adaptive boundary thus affects environments on both sides of the boundary, so that physiology – living function – is necessarily ‘extended’. (8) Life is, in essence a conspiracy of environments sequestered by adaptive boundaries.
The thermodynamic organism may also be represented as a standing wave of specified low entropy (Figure 5). The organism (or more accurately, the process that the organism embodies) rides along on the crest of a standing wave of specified low entropy, metabolic work being done to draw matter up the trailing face of the wave, followed by a second-law-driven cascade of matter down the wave's leading face on its way to thermodynamic equilibrium. The crest of the wave represents a disequilibrium state that stands as long as metabolic work creates order at the same rate as it degrades. If metabolic work fails, the standing wave degrades to a uniform level of thermodynamic equilibrium: again, death.
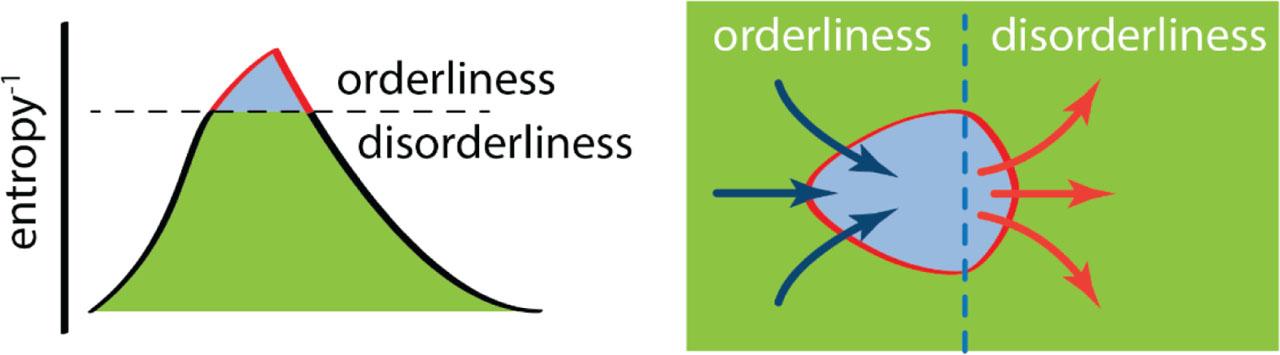
The organism as thermodynamic standing wave. Left: The organization of matter into the organism by the adaptive boundary. Right. A top down look at the persistent dynamic disequilibrium. The blue-shaded region bounded by red represents the organism. The green-shaded region represents the environment.
The thermodynamics illustrated in Figures 3 and 5 can apply both to living systems and physical systems. The formation of orderly arrays of Bénard cells in layers of heated fluids is but one example of the spontaneous emergence of orderliness in open thermodynamic systems, so called dissipative thermodynamic systems (11, 12).
A question immediately arises. If life is a phenomenon distinct from the purely physical, what is life's distinctive quality? I have argued elsewhere (13, 14) that life's distinctive quality is homeostasis, a concept expressed by Charles Darwin's French contemporary and physiologist, Claude Bernard. Bernard's usual sobriquet is the ‘father of experimental medicine’. The sobriquet is apt: Bernard was a skilled experimentalist and empiricist, who pioneered the modern science of physiology. As an aside, the word homeostasis was not coined by Bernard, but was introduced into the lexicon by the early 20th century physiologist, Walter B. Cannon, in his 1932 book The Wisdom of the Body (16).
Usually, Claude Bernard and the concept of homeostasis are introduced to modern students by citing Bernard's famous aphorism:
The steadiness of the internal environment is the requirement for a free and independent life.
There is more to Bernard's scientific philosophy than this aphorism. The concept of homeostasis has a complicated and protean history, making it perhaps the most misunderstood and thoroughly trivialized concept in modern biology (13, 18). Contrary to Bernard's usual image as a strict materialist, his concept of homeostasis has deep roots in the vitalist traditions of Hippocratic medicine, of health comprising a balance of opposing tendencies. These concepts were themselves inspired by pre-Socratic notions of nature comprising a balance of opposing properties, specifically earth, air, fire, and water. The thermodynamic organism described above fits comfortably within that concept of balance, albeit expressed in the language of thermodynamics, disequilibrium, and equilibrium. The language is essential. Homeostasis is often misconstrued as some form of equilibrium. In terms of the thermodynamic organism, homeostasis is rather a precariously perched disequilibrium, sustainable only so long as work is done to offset the inevitable and ongoing degradation of low entropy states to maximum entropy.
Our modern conception of homeostasis, which I shall henceforth call the cybernetic metaphor, was largely inspired by the engineer Norbert Wiener. Bernard was greatly admired by Wiener, who sought to create self-regulating machines that modeled Bernard's self-regulating organisms (19, 20). In Wiener's cybernetic metaphor, homeostasis is attained through the operation of machines, such as feedback controllers, that regulate particular physical properties, such as temperature, similar to the regulation of temperature by a negative feedback thermostat (21).
The cybernetic metaphor for homeostasis is more than an analogy: it is a particular statement of cause and effect. The regulated property is the effect, and the cybernetic machine is the cause. Bernard's conception of homeostasis is in fact the inverse of Wiener's (Figure 6). To Bernard, homeostasis was the fundamental principle that distinguishes life from non-life. While Bernard's great skills as an experimentalist and empiricist allowed him to tease out the details of the mechanisms behind the steadiness of the internal environment, Bernard was nevertheless a ‘small-v vitalist’, in that he regarded life as distinctive in some fundamental way from non-life. His famous aphorism expresses this: the ‘steadiness of the internal environment’ is the predicate for the ‘free and independent life’ (13). The inversion is crucial: being life's fundamental quality, homeostasis is not derivable from mechanism. Rather, the mechanism is the outcome of life's fundamental quality: homeostasis is now the cause, and the mechanism is the effect.
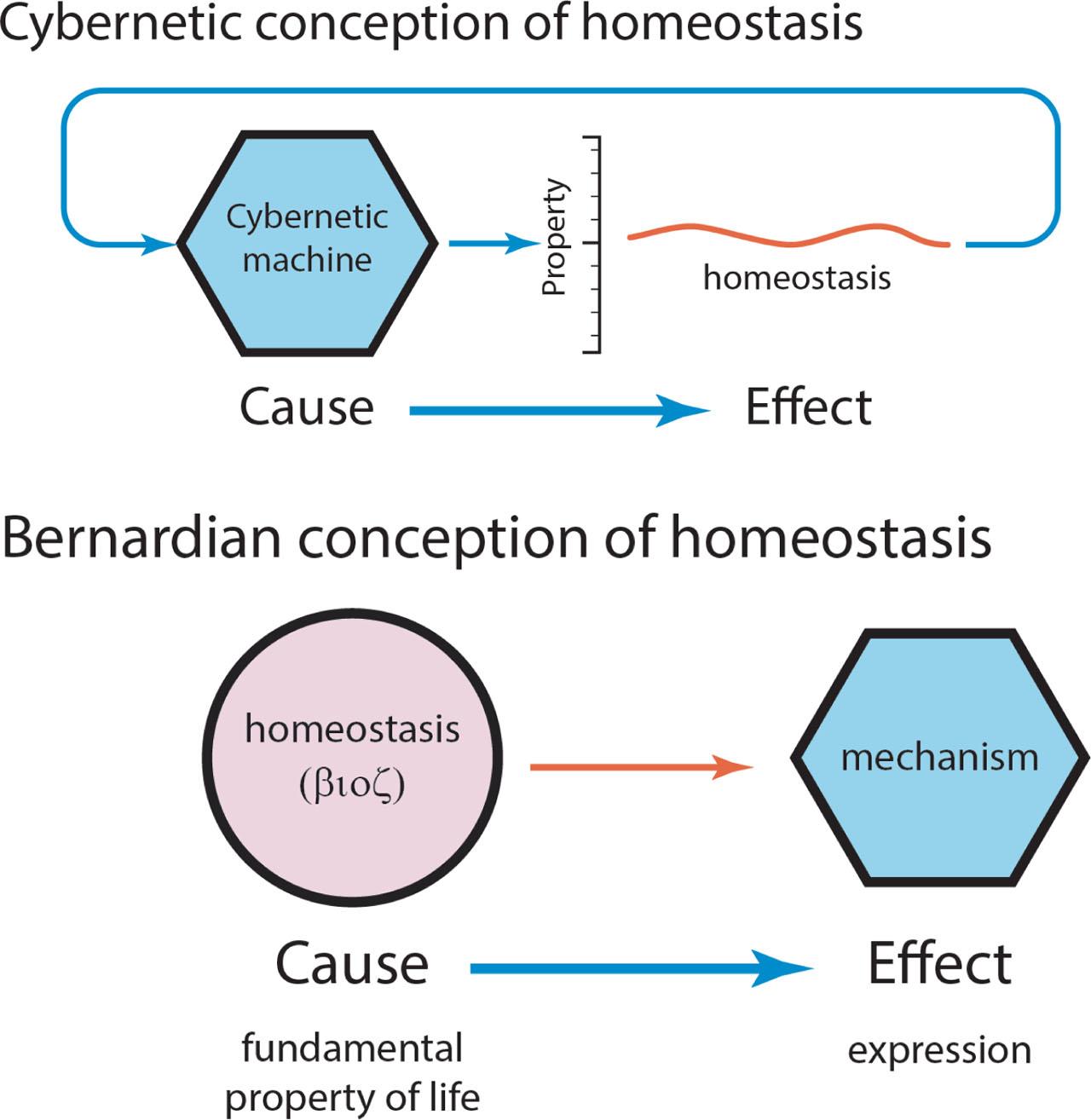
Cause and effect in the cybernetic conception of homeostasis and its inversion in the Bernardian conception.
Bernard's small-v vitalism derived from a school of thought that emerged in the late 18th century known as process vitalism. This vitalist philosophy emerged from the medical faculty of Montpellier University, largely through the work of the physician Théophile Bordeu. For many centuries prior, medicine had been dominated by doctrines of essentialist vitalism, which looked to ineffable vital essences or vital forces to impart life to inanimate matter. Lamarck's posited ‘adaptive force’ and ‘complexifying force’ were drawn from that well.
Essentialist vitalism had largely fallen from favor by the end of the 18th century (13). In its place, process vitalism attributed life's distinctive nature not to an ineffable force, but to a process of endless negotiation and mutual accommodation among an organism's ‘many little lives’ – cells, organs, even organisms – in assemblage (13, 14). Bordeu himself was inspired by the organism-like behavior of bee swarms. Bernard's experimental philosophy derived from the concept of the ‘many little lives’, whose negotiations and mutual accommodation he teased out through the language of the chemistry of the processes.
Bernard's conception of homeostasis is close cousin to the Aristotelian concept of the βioς (Figure 6), an idealized state toward which the organism strives (22). Each type of organism strives toward its own βioς. The material form and function of the organism is the physical embodiment of the βioς. Form and function may adapt to a range of environments – plumage or pelage may thicken in cold conditions, worked muscles may enlarge, for example. I return to this below.
An adaptive boundary works to sustain an entropy gradient across it (Figure 7), doing metabolic work that organizes fluxes of matter and energy across the boundary, creating specified orderliness as rapidly as it degrades. The entropy difference represents a difference of potential energy (ΔPE), which is proportional to the work needed to sustain the internal low entropy state.
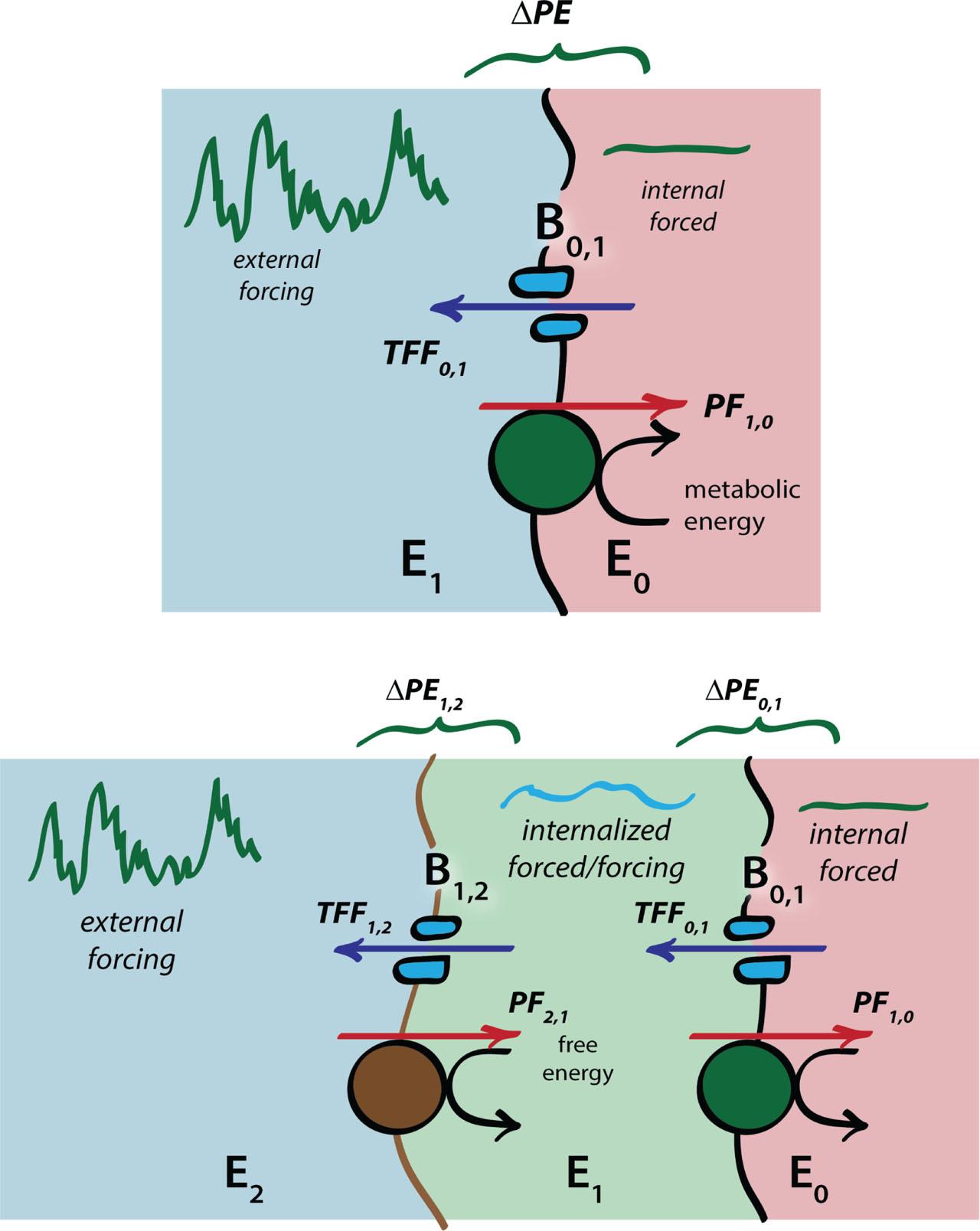
Extended homeostasis and nesting of adaptive boundaries to internalize external environments. Numbering scheme for boundaries and environments follow Figure 4.
Homeostasis of the internal environment is attained when work is done to drive a physiological flux (PF) against the PE gradient sufficient to offset the thermodynamically-favored flux (TFF) demanded by the second law. Because both the PF and TFF are tied to a flux of matter, it follows from the conservation of mass principle that homeostasis on one side of an adaptive boundary necessarily imposes homeostasis on the other side as well: extended homeostasis, in a phrase (8). The modification of soils by the earthworm (Figure 4) is an example of extended homeostasis. The ‘signal’ of extended homeostasis here is the adaptive modification of the worms' soil environment.
The signal of extended homeostasis can be difficult to discern if it is overwhelmed by strong forcing by external environments (Figure 6). If the forcing from the external environment fluctuates rapidly and chaotically, as it would likely do if the environmental forcing is strong, this will impose commensurate large and unpredictable metabolic demands on the adaptive boundary (Figure 7, top). These can be ameliorated by nesting adaptive boundaries within one another. By nesting B0,1 inside a new adaptive boundary (B1,2), E1 now becomes an internalized ‘external’ environment, managed by B1,2, so that the metabolic demands on B0,1 are ameliorated and damped (Figure 7, bottom).
A compelling example of extended homeostasis may be found among the animal-built structures of the Macrotermitinae, a family of termites that, among other attributes, organize their societies into a consolidated subterranean colony, embedded deep in soil, and located below a large epigeal soil mound constructed by the colony's workers (23,24,25,26,27). The mound, which stands several meters tall, is the most prominent physical evidence of the colony's presence. The mound is also a dynamic structure, cycling roughly 250 kg of soil through it annually as worker termites transport soil from underground up into the mound to offset soil eroded away by wind and rain. It has been described as a slow-motion ‘soil fountain’ (28), a literal expression of the standing thermodynamic wave described in Figure 5.
The construction of the mound and its architecture represent at least two forms of extended homeostasis. The simplest involves nest humidity, which is maintained at 60%–70% throughout the annual rainfall cycle of summer torrential rains and winter drought. In the dry winters, workers mine water from soil aquifers, and physically transport it up into the colony (PF), offsetting loss of water from the colony to the dry environment (TFF). In the wet summers, water is actively transported in the form of wet soil from the nest up into the mound (PF) and deposited on the mound surface, offsetting the percolation of water into the nest from the rain-soaked soil (TFF). In this way, the 12-month cycle of winter drought and summer rainfall is damped to produce the nest's steady humidity through the year.
A more complex form of extended homeostasis arises from the mound's function as a wind-driven lung. The colony has a considerable metabolic rate, in the tens of watts, which is sustained by exchange of the respiratory gases (oxygen, carbon dioxide, and water vapor) with the atmosphere (29). The driver for the colony's respiratory gas exchange is the interception of turbulent wind energy across the mound's porous surface. Velocity of turbulent wind varies chaotically which means the forcing of respiratory gas flux is also chaotic. For their part, termites work to maintain a steady environment within the nest, and respond to any temporal variation of air movements, or to temporal variation of partial pressures of the respiratory gases, by building barriers that impede air movements, and consequently impede respiratory gas exchange. The mound is the compromise that balances these conflicting processes. The complex internal tunnel network within the mound gradually damps the chaotic wind energy intercepted at the surface to the air spaces around the colony, while still allowing respiratory gas exchange to proceed. Remarkably, this is similar to how the architecture of the alveolar lung of mammals damps the cyclical variation of ventilation to maintain a steady environment within the alveoli and gas exchange between air and blood.
Extended homeostasis is essentially the physiological dimension of niche construction, to which I shall return below (30, 31). Where niche construction theory was inspired by Richard Dawkins' metaphor of the extended phenotype (32), and was initially built upon population genetics models that accounted for accrual of fitness from modification of environments, the extended organism metaphor introduces essential attributes of organisms, specifically extended homeostasis (8).
Compared to Wiener's cybernetic metaphor, Bernard's conception of homeostasis does not sit comfortably in modern biological thought, for two reasons. First, it subordinates mechanism to a fundamental quality of life that itself cannot be reduced to mechanism. Second, and related to the first, Bernard's conception opens the door for admitting frank purposefulness to the phenomenon of life itself, and (more controversially) to shaping the history of life – evolution. Both are dimensions of the same overarching phenomenon, namely adaptation.
Adaptation is literally ‘tendency to aptitude’, which is an organism's ability to adjust form and function to better fit the organism's circumstances. Adaptation can also shape lineages of organisms, presumably through natural selection of genomes that produce apt function and form, although this is a point of some dispute (33).
It is possible to construct a cybernetic model for adaptation (Figure 8). A cognitive engine senses the state of the internal environment and compares it against some internalized set point. The cognitive engine may consist of various proxy measures of the trait under control. For cell salt and water balance, for example, stretch of the cell membrane is the proxy measure of changes of cell volume, which are themselves influenced by gradients in salt concentration and water potential across the membrane (34, 35).
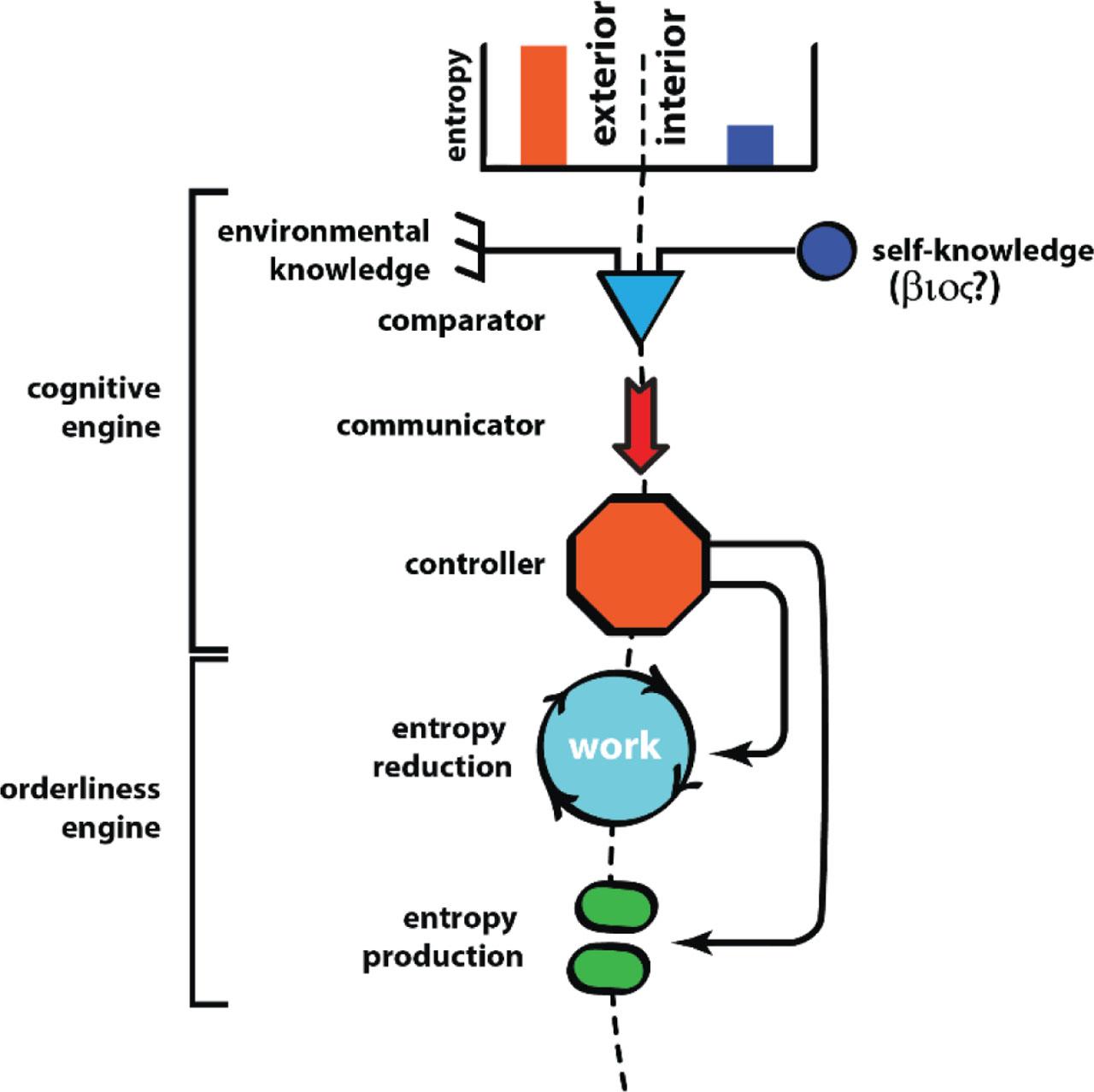
Homeostasis and adaptation in a cybernetic framework.
The comparison directs an orderliness engine, which mediates the order producing work needed to restore the internal state toward the set point. Continuing the example of cell volume regulation, the energy feed to cation ATPases may be changed, or aquaporin permeability may be adjusted. The result will be maintenance of cell volume in the face of a range of environmental salinities that otherwise would cause the cell to swell or shrink.
At higher scales of organization, the cybernetic metaphor becomes increasingly problematic (36). The organism comprises innumerable regulatory processes (‘-stats’), such as for temperature (thermostat), electrolyte balance (osmostat), or metabolic rate (metabolostat), each presumably requiring its own cybernetic system to operate, and which overlap and interact. The current fashion for dealing with this complexity is to treat the unitary organism as an assemblage of ‘-omes’ – the genome, transcriptome, proteome, metabolome, and so forth – which interact to produce the well-functioning organism (37).
Whether it is multiple ‘-stats’ or multiple ‘-omes’, the cybernetic metaphor by its nature anatomizes the organism into a proliferating assemblage of cybernetic systems. At increasing levels of complexity, the mechanistic aspirations of the cybernetic metaphor become shakier, and at the organism level, break down utterly. The organism is itself a form of homeostasis, a dynamic disequilibrium that persists through time by continually balancing order-producing work against thermodynamic decay. One could conceivably construct a cybernetic uber-system (an organismostat?) that manages the organism-level fluxes of matter and energy that sustain the organism's form and function, as well as all the organism's sub-divisions. But what would the elements of such a cybernetic system be? What, for example, would be the putative ‘organismostat set point’ (Figure 8). At increasing levels of complexity, the cybernetic metaphor converges onto something akin to the frankly purposeful Aristotelian concept of the βioς (Figures 6 and 8). The ‘set point’ of the cybernetic metaphor becomes indistinguishable from a kind of self-knowledge of what the system intends to be, with the orderliness engine being purposefully directed to realizing the system's self-knowledge. For extended organisms with extended homeostasis, the cybernetic metaphor becomes even more tenuous.
At root, homeostasis is a cognitive phenomenon. The adaptive boundary is a sensory interface, which senses environmental circumstances, and maps that information onto some internal cognitive representation of the environment. This cognitive map is then compared to the organism's self-knowledge of what it intends to be (Figure 9). The adaptive boundary is also a motor interface, doing physical work on the external environment, shaping it so that the ‘real world’ is brought into conformity with the βioς. This is intentionality: the organism intends to create an external environment that is suitable for sustaining the βioς (15, 38,39,40). Simple adaptation follows when the cognitive map tracks the ‘real world’ as it changes, and directs the motor interface to shape the real world, again with the aim to sustain the βioς, that is, the persistent thermodynamic disequilibrium that is the organism. This is adaptation as frank, not apparent, intentionality.
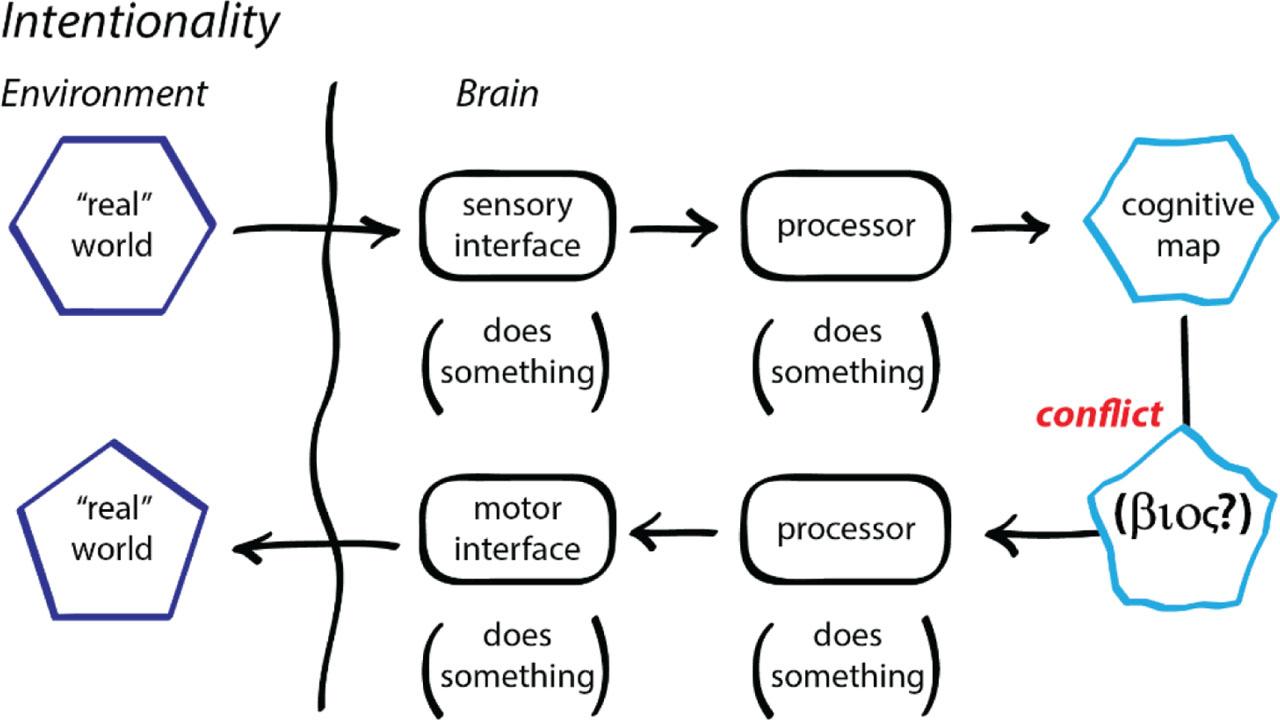
Cognition, homeostasis and intentionality.
Cognitive maps also may be shaped independently of direct inputs from the sensory interface. There may be inputs from other cognitive and adaptive systems within the organism (Figure 10). In this circumstance, cognitive maps can arise that are decoupled from a direct sensory inputs: imagined, in a word. Creativity emerges when the motor interface works to create a ‘real world’ that conforms to the imagined cognitive representation. Novelty is the product of intentional creativity.
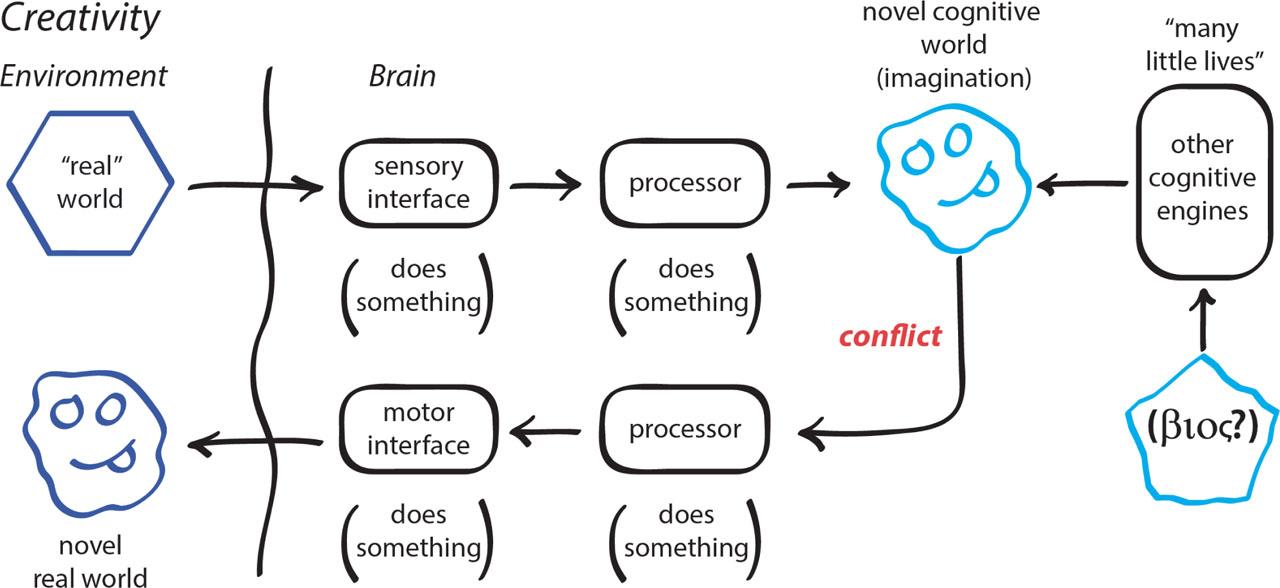
Cognition, homeostasis, and creativity.
The creation of an entirely new soil environment by populations of earthworms exemplifies this type of creativity. Populations of termite colonies do as well, creating new ecosystem wide systems of hydrology that would not exist in their absence (41). In both instances, and more, new worlds are created driven by the organisms' intention and creativity.
Evolutionary adaptation has been construed as present function informed by the inherited memory of past apt function (42, 43). Gene selectionism asserts that the sole, or at least the most important, repository of hereditary memory is the gene, or more precisely, the nucleotide sequences in nucleic acids (3). These sequences are passed on with high fidelity with each replication, and from generation to generation. This gene-centered view imposes a strong distinction between organismal adaptation and evolutionary adaptation. The organism may adapt during its lifetime, but those adaptations die with the organism. Only adaptations encoded in the genotype endure across generations.
McClintock's concept of the genome as an organ of the cell dissolves that distinction. The nucleotide sequence code of DNA becomes a type of memory token, an object memory, in itself capable of nothing. It is the expression of object memory as function, a process memory that is embedded in the complex physiological milieu that is governed by an adaptive boundary. Process memory includes a dimension of time that does not apply to object memory. Object memory exists: it either is, or is not. Process memory, in contrast, persists.
Memory is not only a remembrance of the past, it is a means of biasing the future. The strength of a memory is determined both by how strong the future bias is, and by how far into the future the bias extends. Like life itself, process memory is subject to the same dynamics of disequilibrium, of creation and expression balanced against thermodynamic decay. Process memory therefore operates on a continuum, from very short, which may be termed physiological memory, to long, which if long enough, may be termed hereditary memory (Figure 11). Where the one transitions into the other will depend upon context. The cell, for example, is a low entropy state that is subject to a particular rate of thermodynamic decay (the TFF, Figure 3), and that persists as long as order-producing work can be done to offset it (the PF, Figure 3). An enzyme complex may shape function for only a few seconds, for example, less persistent than the cell itself. Within that context, this is effectively a form of physiological memory. If, on the other hand, the cell constructs an environment that persists for longer than the cell itself, that process memory effectively becomes a form of hereditary memory for the cells that construct it (Figure 11). Working up through the levels of the nested adaptive boundaries that comprise the organism, process memories can be simultaneously physiological and hereditary. If, for example, an earthworm's nephridial epithelium (B1,2, Figure 4) creates an environment (E1) that persists beyond the lifetimes of the cells (E0) living there, the maintenance of E1 is a form of hereditary process memory. Just as the nesting of adaptive boundaries creates newly internalized environments (Figure 7), process memories can be simultaneously physiological and ‘hereditized’ by context.
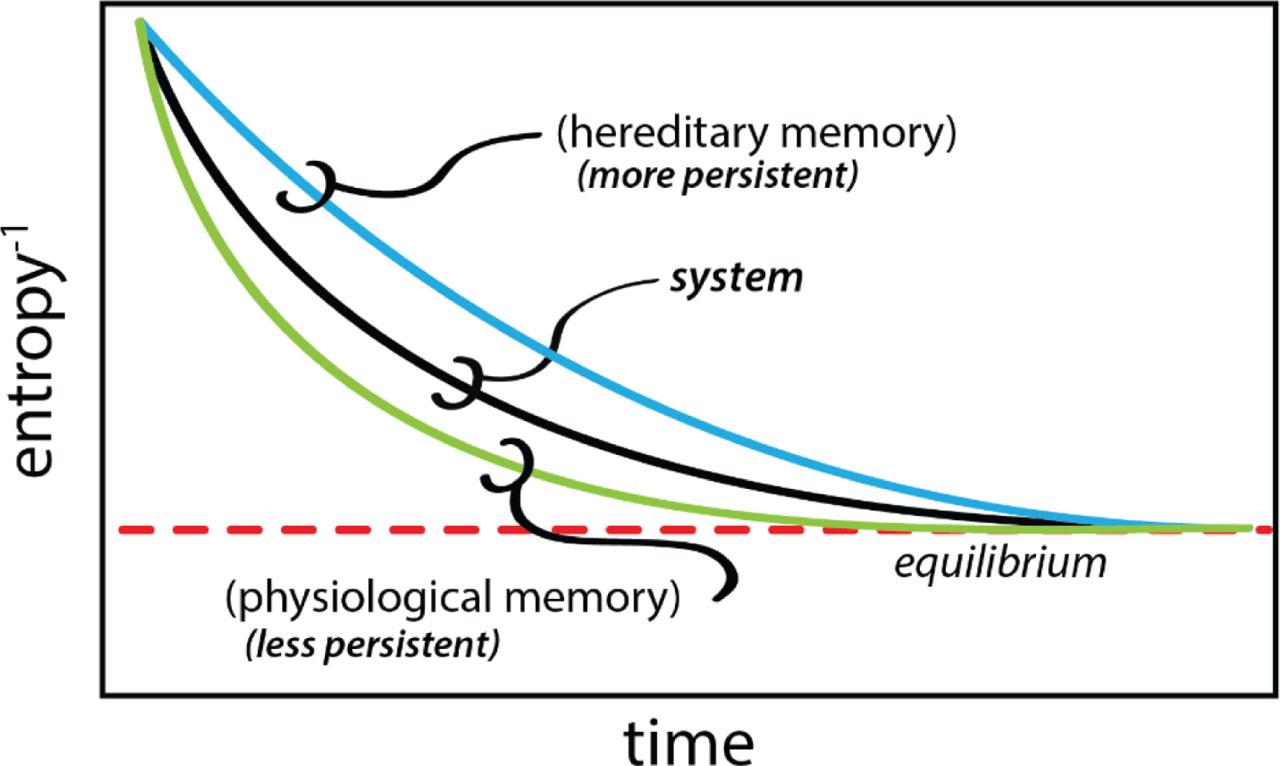
Process memory and the continuum of physiological and hereditary memory.
All process memories subordinate to the context of the organism are, by this criterion, physiological. Organisms are, however, extended, and this means that organism-created environments can also embody process memory. This may be physiological memory if the organism-created environment does not persist beyond the organism's lifespan. If the organism-created environment persists beyond the organism's lifespan, the process memory counts as hereditary: it is a persistent constructed environment that informs the function of future generations.
The Macrotermes colony provides a useful illustration of this. The mound represents an extended environment created by the workers, making the constructed environment a form of physiological memory for the workers. A focal deposition of soil by a worker termite serves as physiological memory for other workers in that cohort, generating a form of collective building known as stigmergy (38, 44, 45). Worker termites have lifespans of a few weeks, whereas the mound can persist for periods of months. The constructed environment of the mound therefore serves as a form of hereditary memory for subsequent cohorts of workers. The mound persists as long as the queen lives, who has a typical lifespan of 10–20 years (41, 46, 47), making the mound environment a form of physiological memory for her. When she dies, the mound gradually disintegrates over the course of a few years, as the population of workers dies out. However, the colony's activities remodel soil composition and hydrology on regional and ecosystem wide scales, with effects that can last for time scales ranging from centuries to millennia (48), creating a form of heritable memory for subsequent generations of termite queens (41). These long-lasting environmental modifications enable Macrotermes colonies to extend their range into otherwise inhospitable environments.
This expands the range of heritable memory far beyond the narrow gene-centered form that constitutes Neodarwinism. And because these are mediated by extended homeostasis/βioς, it introduces a cognitive/purposeful dimension to evolutionary adaptation that the Neodarwinist idea excludes a priori (49).
A fundamental question arises from this expansive conception of hereditary memory, to wit: what drives the course of a lineage's evolution? Richard Dawkins has argued that it is ultimately the gene – the object memory – that is the driver, on the grounds that the information in DNA sequence code endures far longer than any conceivable effects that phenotypes might bring about (50). By this logic, it is object memory – the genome and its slow modifications – that is evolution's default driver.
However, the genome itself is embedded in a physiological milieu that is defined and shaped by a complex network of epigenetic feedbacks, which can include heritable modifications of the genome (Figures 2 and 12). This raises the fundamental question. Is it genomes and their modifications that drive lineages into the future? Or are genomes largely drawn along in the epigenetic wake of the action of adaptive boundaries? If the former, the Darwinian idea of natural selection is vindicated. If it is the latter, the Darwinian idea is undermined, because selection is no longer natural, but is the expression of the frank intentionality of adaptation.
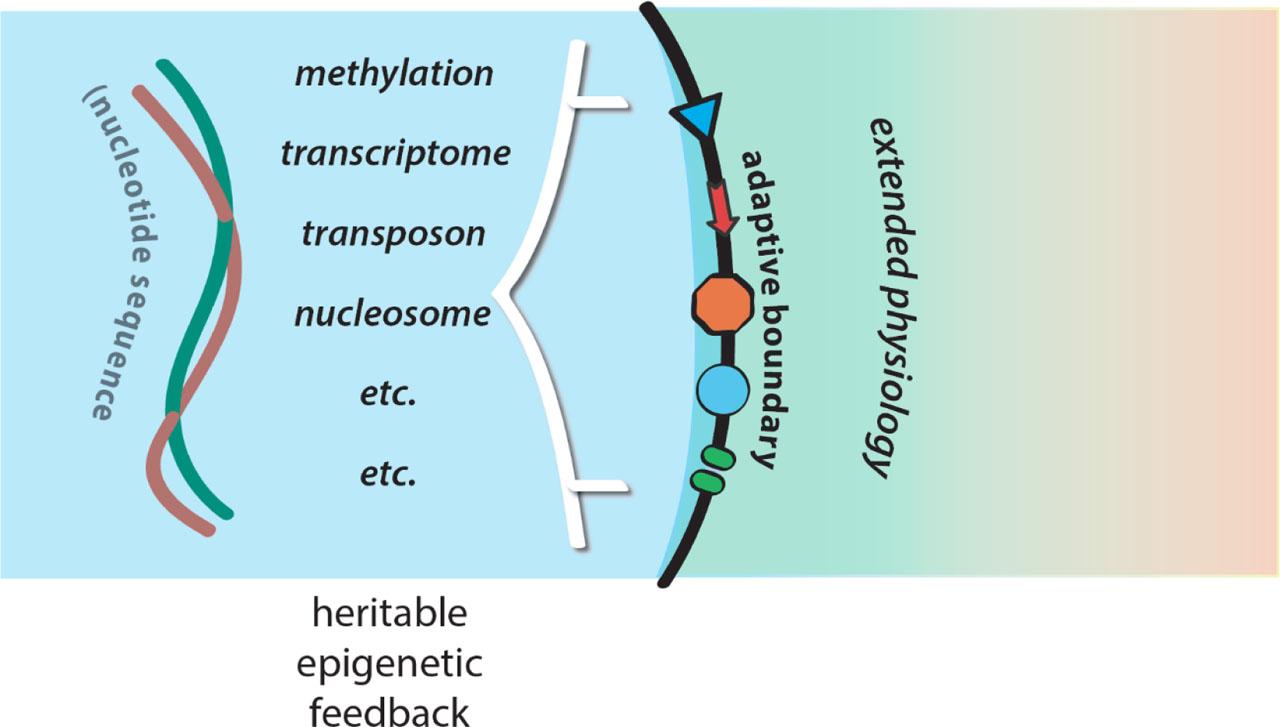
Epigenetic feedbacks on the memory token of nucleotide sequence code.
I may now return to the question I posed at the beginning of this essay: whence evolutionary novelty, the source of Darwin's ‘endless forms most beautiful’? The prevailing answer is Neodarwinist: mutation filtered by gene selection. I argue that a better answer is the inherent purposefulness of homeostasis combined with the dynamics of the extended organism.
In a regime of gene selectionism, evolutionary novelty is a form of environmental determinism. A physical environment presents a range of variation in some property, say temperature (Figure 13). Organisms living in that environment possess a variation in genomic aptitudes. Natural selection will favor genomes that match aptitude to prevailing environments, and their numbers will thereby increase (Figure 13, top). At the extremes, say too warm or too cold, genomic aptitudes diminish, these variants will not be selected, and their numbers will decline. Natural selection therefore is centralizing, acting to conform genomes to the most prevalent environmental conditions (Figure 13, top).
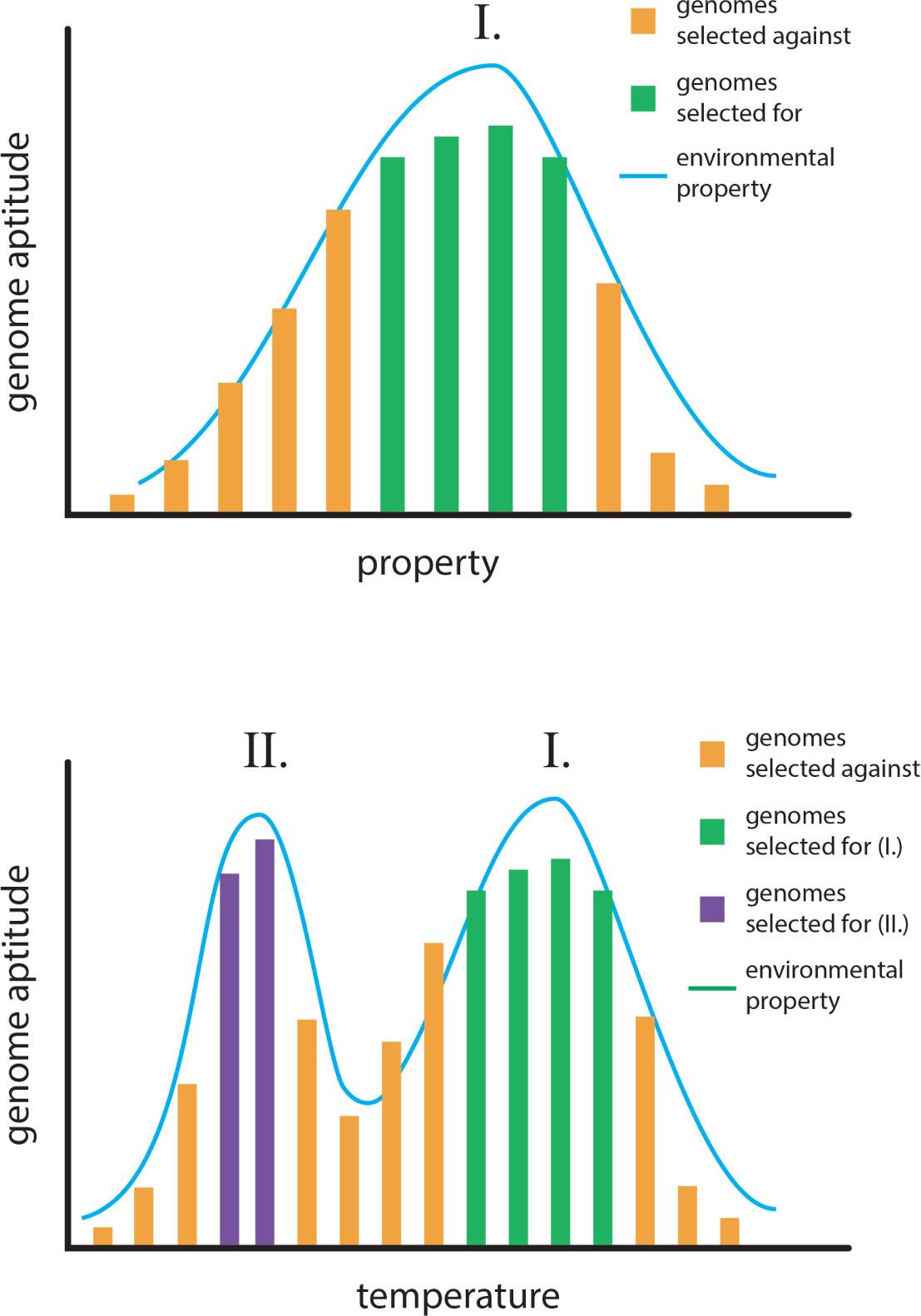
The Neodarwinist model of evolutionary novelty. Top: The centralizing tendency of gene selectionism. In an environment that varies in some property (e.g., temperature), some genomes will confer high aptitude and be selected for (green bars), while some will be less apt and be selected against (orange bars). Bottom: Some genomes that confer inaptitude in one environment (I.) may confer high aptitude in another (II.), and will be selected for (purple bars), producing a novel genome.
In this conception, evolutionary novelty arises through environmental novelty. If a new environment presents itself (II, in Figure 13, bottom) where genomic variants that were inapt in environment I are now apt for environment II, those genomes selected against in environment I are now favored in environment II (Figure 13, bottom). This is essentially Sewall Wright's ‘adaptive peaks’ model as refined by G Evelyn Hutchinson's ecological niche concept (51, 52).
This model for evolutionary novelty needs no agency on the part of the living organism to operate. It is simply a sorting mechanism that is governed by the variations and vicissitudes of environments (54). In all imaginable scenarios, genomes (or their expression as organisms) are at the mercy of the environment, which renders the organism as either apt for the environment, or inapt.
This model, which we may call crude gene selectionism, is now widely regarded as too narrow in scope to account for the broad scope of evolutionary process, which includes disparate concepts like evo-devo, chromosomal dynamics and epigenetics (51, 55, 56). The extended evolutionary synthesis (EES) seeks to unify these under a broad Darwinian umbrella (Figure 14). Gene selection still exists, but it is embedded in this complex network of broader types of selection. Notably absent from the EES, however, is the phenomenon of adaptation (Figure 14). This is excluded perhaps because adaptation introduces a frankly purposeful form of agency into evolution. This fundamentally undermines the Darwinian idea: if organisms purposefully strive to adapt, rather than are simply selected because they are apt, species in some sense ‘want to evolve’ (57), an inference that can elicit strident disapproval (58, 59).
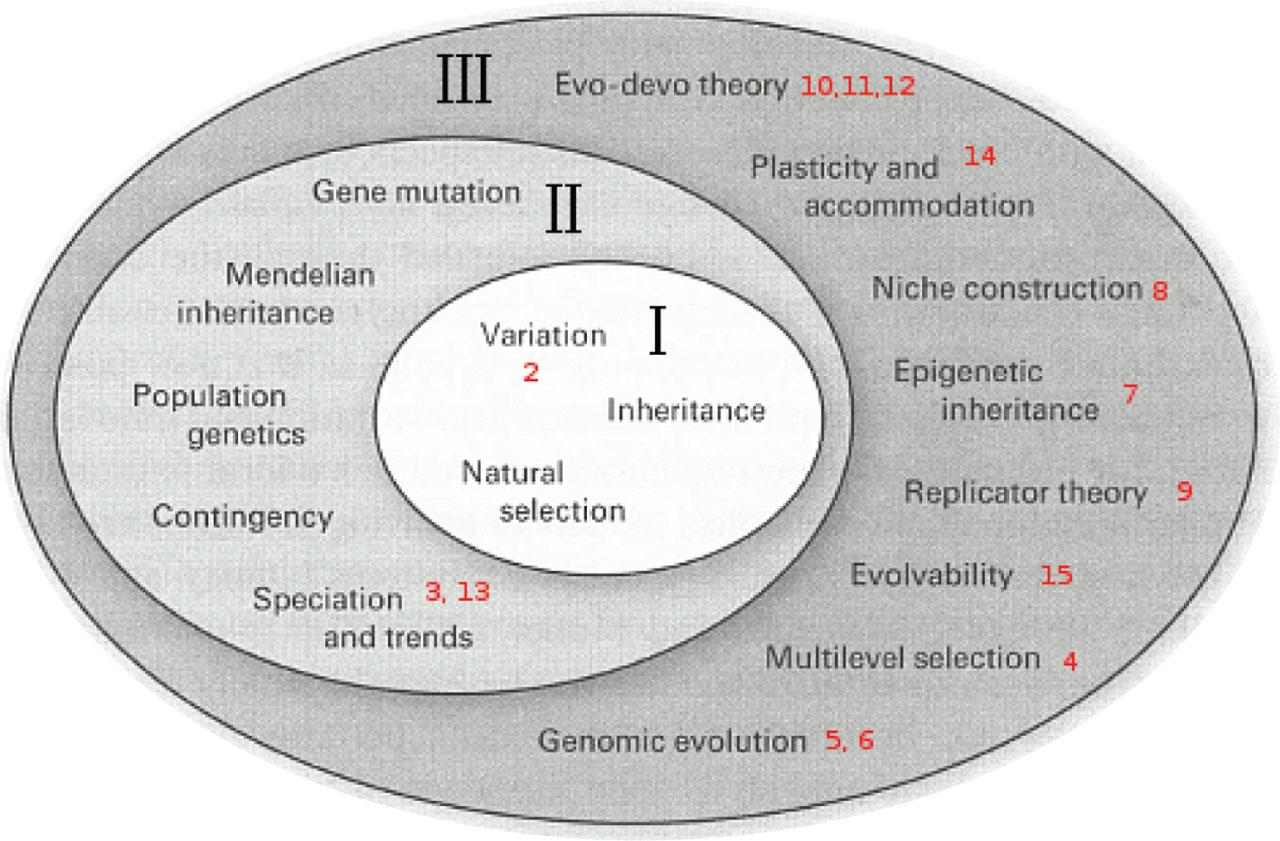
The EES. From: Pigliucci, Müller (2010) Evolution, The Extended Synthesis, page 11. EES, extended evolutionary synthesis.
The extended organism idea, which melds the Bernardian conception of homeostasis with niche construction (61), and the related Aristotelian concept of the βioς, offers an alternative model for evolutionary novelty, which rests on four supports: the thermodynamics of adaptation; broadening the scope of hereditary memory; dissolution of the distinction between genotype and phenotype; and dissolution of the distinction between organismal and evolutionary adaptation.
Imagine an extended organism bounded by an adaptive boundary that creates a surrounding region of extended homeostasis (Figure 15, top). Functional aptitude derives from the work of homeostasis, that is the work rate needed to sustain the standing wave of persistent thermodynamic disequilibrium that is the organism. Functional aptitude is maximized when the work of homeostasis is minimized. Persistence of the extended organism is governed by an interaction of process and object memory, and organized around the organism's self-knowledge of what it intends to be (the βioς, for convenience).
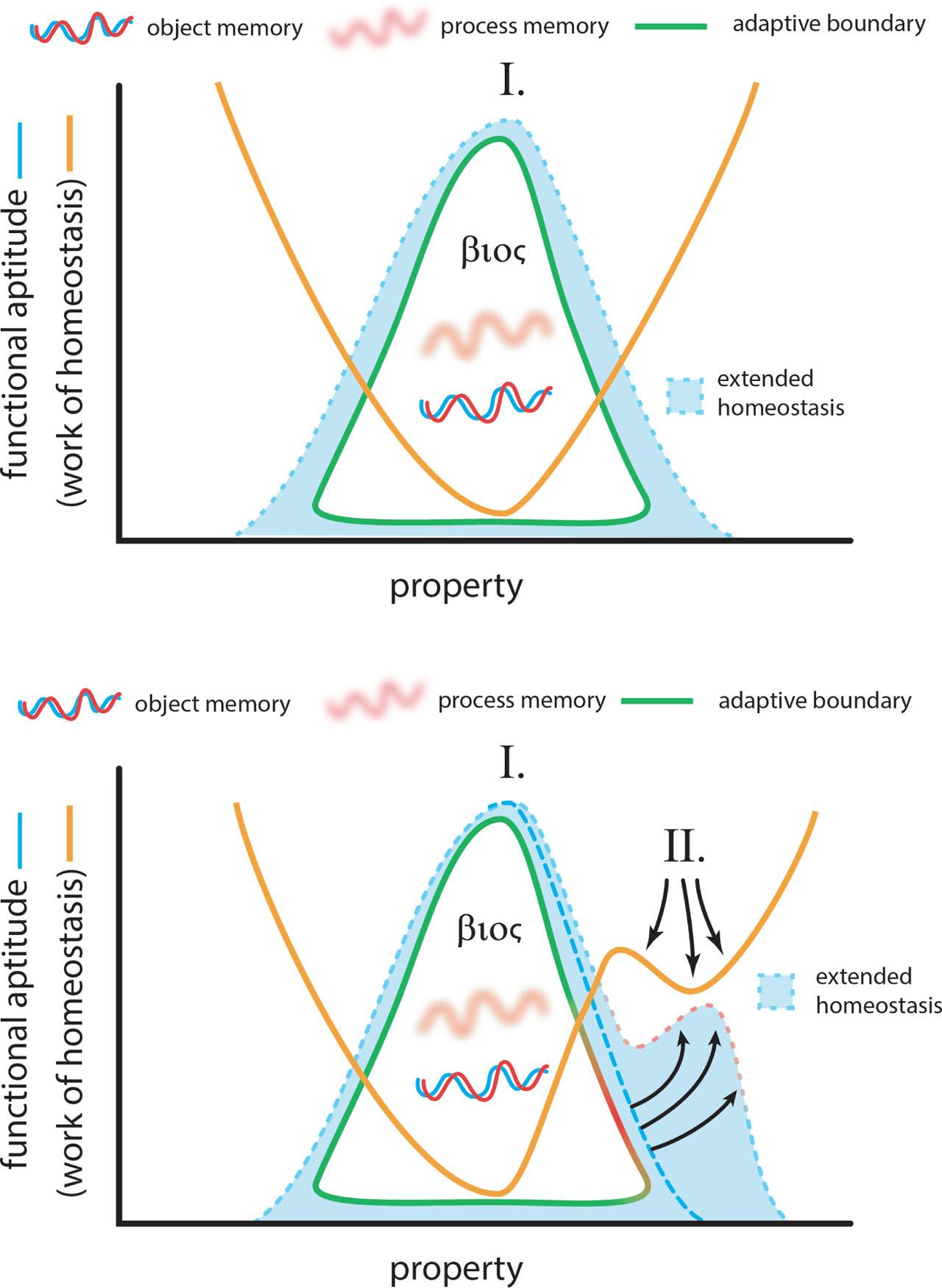
Extended homeostasis and evolutionary novelty.
Similar to the Neodarwinist model, functional aptitude in the extended organism model can gravitate toward a minimum for work of homeostasis (labeled I. in Figure 15, top). The difference comes in at the environmental extremes. In the Neodarwinist model, environmental extremes select against inapt genomes. Novelty only arises by the introduction of a new sorting filter, a new environment that selects genomes that were inapt in the original environment. In the extended organism model, environmental extremes offer new opportunities for purposeful striving. By intentionally creating new environments that channel matter and energy toward the organism's specified thermodynamic disequilibrium, the extended organism effectively creates anew a local minimum in the cost of homeostasis (labelled II. in Figure 15, bottom). Novelty in this circumstance arises at the environmental extremes, in the form of intentional goal-directed modification of environments, the goal being persistence of the βioς. Novelty becomes evolutionary novelty when these intentional modifications outlast the extended organism and verge into hereditary memory (Figure 11).
The extended organism idea liberates the organism from the vicissitudes of an indifferent environment, in which the organism either adapts or dies. The tools of liberation are those fundamental qualities of life – purposefulness, internal intelligence and self-knowledge, striving toward a goal – that underpin life's remarkable ingenuity. It also liberates lineages of organisms from the same constraints, and it does so through the same ingenuity that creates novel forms and functions of organisms – evolutionary novelty.