
Figure 1.
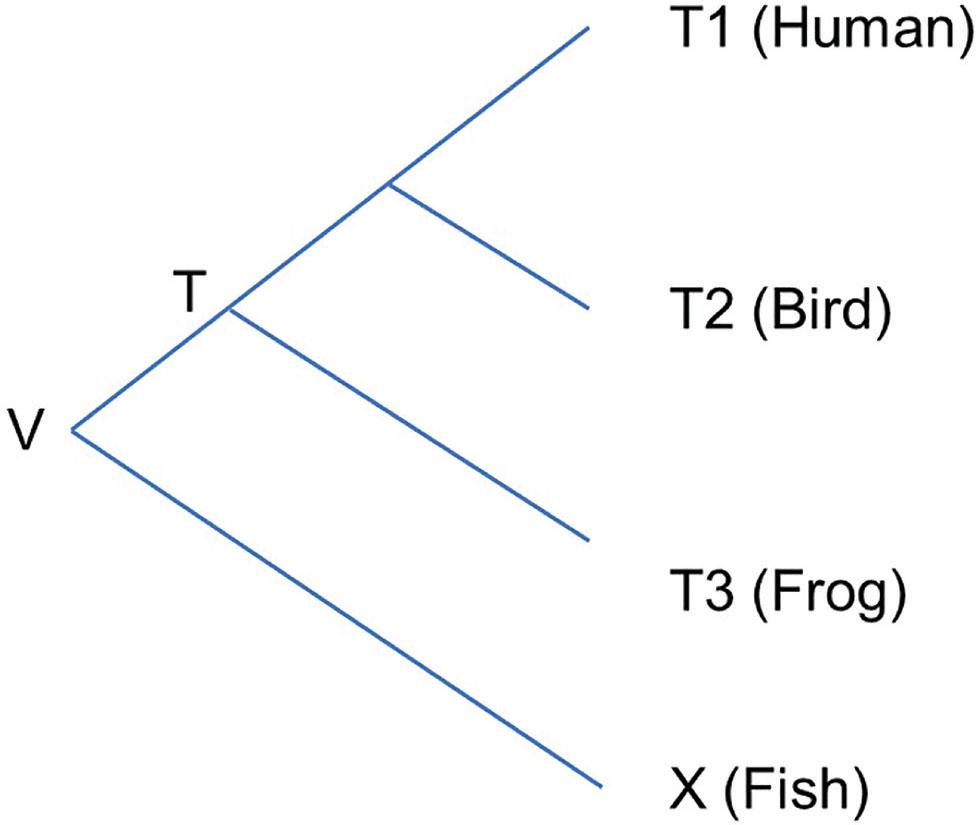
Figure 2.
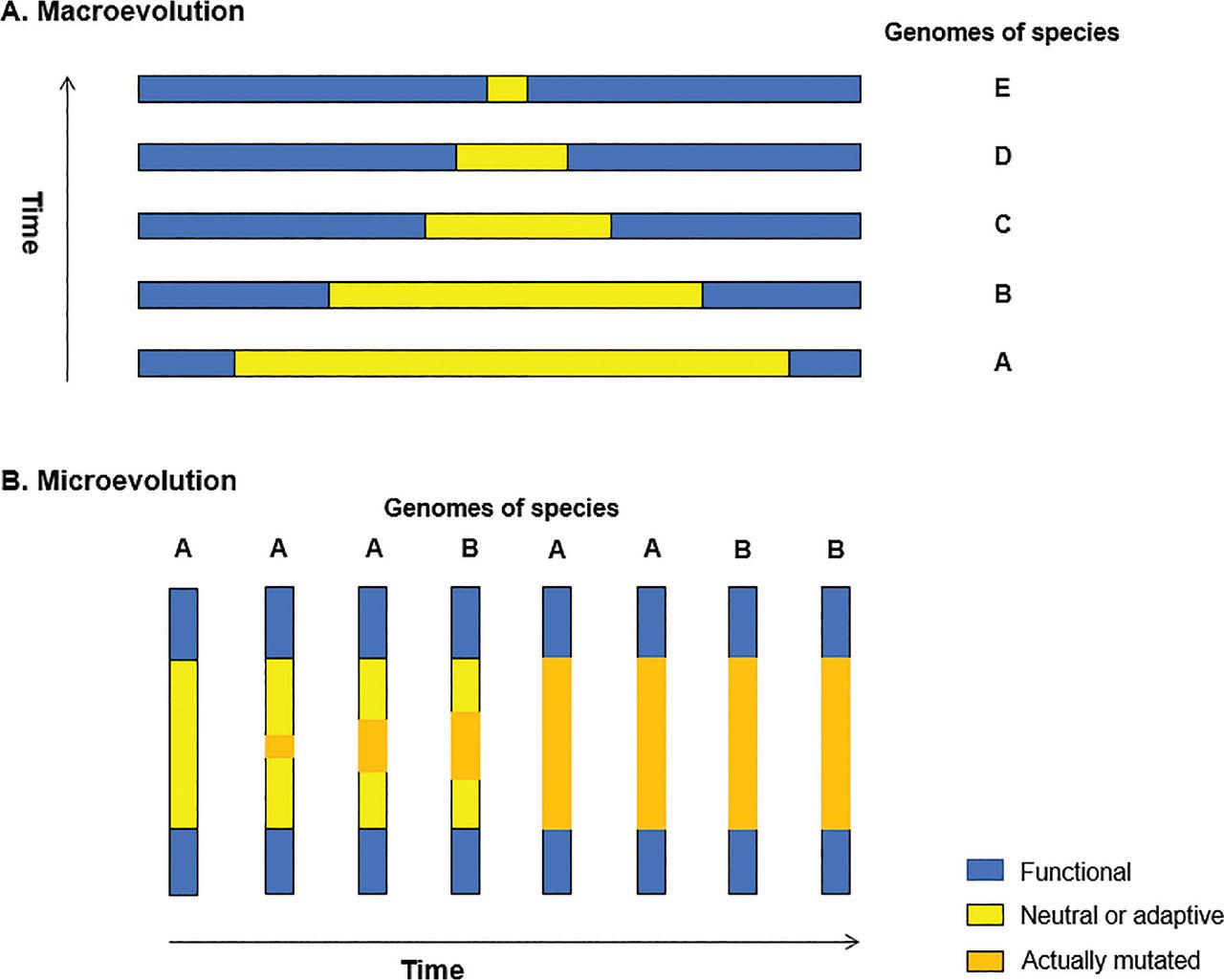
Figure 3.

Figure 4.

Percentage non-identity among species in Dot1_
| Species | Percentage non-identity | ||||||
|---|---|---|---|---|---|---|---|
| YEASA | CAEEL | STRPU | DANTE | XENTR | TAEGU | MYOLU | |
| CAEEL | 71 | ||||||
| STRPU | 73 | 69 | |||||
| DANTE | 70 | 66 | 29 | ||||
| XENTR | 71 | 68 | 31 | 9 | |||
| TAEGU | 72 | 67 | 32 | 8 | 5 | ||
| MYOLU | 72 | 67 | 31 | 6 | 3 | 2 | |
| HUMAN | 72 | 67 | 31 | 7 | 3 | 2 | 1 |
The MGD theory versus the NT_
| Features | MGD theory | NT |
|---|---|---|
| Micro-evo linear stage | NT and natural selection | NT and natural selection |
| Micro- and Macro-evo | Different mechanisms | Same |
| Role of epigenetics | Yes | No |
| Noises suppressed | Yes | No |
| Neutral regions | Complexity dependent | Complexity independent |
| Time frames | Any | Relevant to short times |
| Mutation rates | Any | True for slow rates |
| Genetic equidistance | Maximum and linear | Linear only |
| MGD | Yes | No |
| Genetic variants | Mostly non-neutral | Mostly neutral |
| Recurrent mutations | Common | Rare |
| Quasi-neutrality | Common | Rare |
| Turnover of alleles | Fast and common | Slow and rare |
| Non-conservation | Adaptive function | No function |
| Physiological selection | Yes | No |
| Independent tests | Confirmed | Rejected |
| Axiom based | Yes | No |