
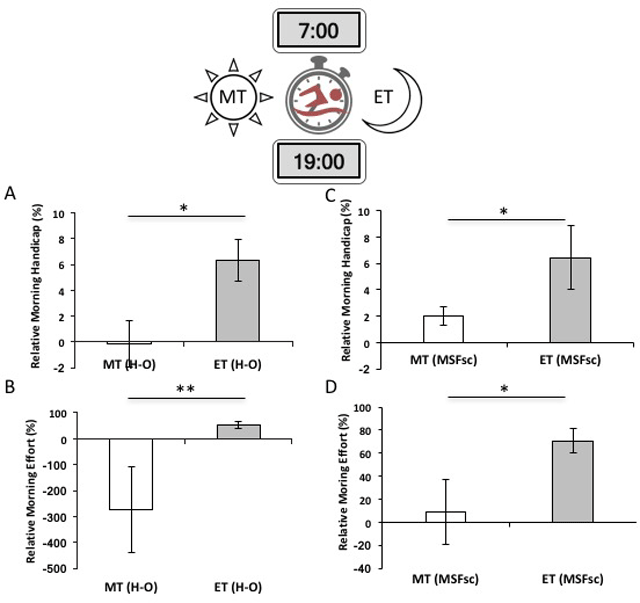
Figure 1
Relative morning handicap representing the average per-individual time-of-day effects on athletic performance and effort in self-reported circadian chronotypes (±SE). A & B) Morning handicap on performance is measured as the average per-individual time trial score ((AM–PM)/PM). Individual trial time (in seconds) differences (AM–PM) are normalized to evening times; values shown represent relative percent differences in performance. A) Evening-types, as measured by diurnal preference (HO-MEQ), have a significantly greater morning handicap than morning-types, swimming 6% slower in morning (07h00) than evening (19h00) 200-meter timed-trials (t = 2.70, df = 12, p = 0.017). B) Morning handicap on performance in chronotypes (MSFsc). Differences between ET and MT are significant (t = 2.45, df = 11, p = 0.034). C & D) Time-of-day effects on physiological effort measured by salivary α-amylase levels. Data are reported as average difference in α-amylase concentration between the morning and evening trials (relative to individual- and time-specific baselines; ±SE). C) For circadian phenotypes measured by diurnal preference (HO-MEQ), both MT and ET participants exert more effort at off-peak times and there is a significant relative percent difference in diurnal effort between MT and ET (t = 2.64, df = 10, p = 0.014). D) For chronotypes (MSFsc), only evening-types exert more effort at their non-peak morning trials and differences between MT and ET are marginally significant (t = 2.64, df = 10, p = 0.05).
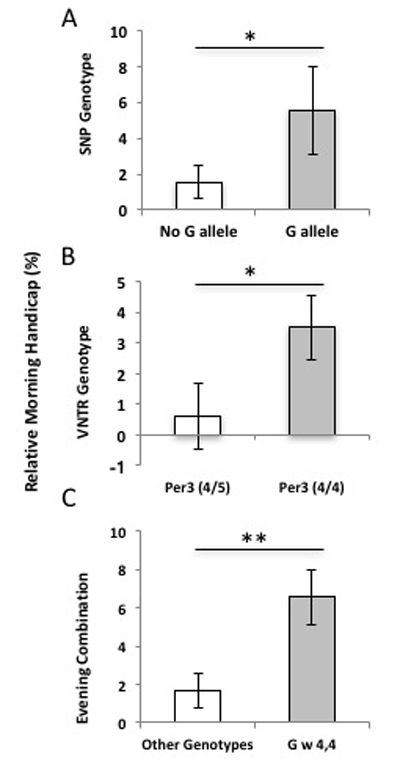
Figure 2
Influence of the PER3 SNP and VNTR polymorphisms on athletic performance. Relative morning handicap represents the average per-individual time-of-day effects (M-E/E) on athletic performance in PER3 genotypes (±SE). A) Participants with a G allele are significantly slower in the morning swim trial (Mw/G = 8.64 ± 0.024) than those homozygous for the C allele (MCC = 1.847s ± 1.140) representing an 6% decrease in morning performance in individuals with G-allele (t = 2.313, df = 23, p = 0.015). B) PER34/4 genotypes show a 3–4% slower trend in 200m swim performance, but this trend is not significant (t = 1.685, df = 23, p = 0.053). Participants with PER34/4 genotypes tend to swim nearly 5 seconds slower in the morning trials (M4/4 = 4.830 ± 1.52) compared to participants with PER34/5 genotypes (M4/5 = 0.667 ± 2.116). C) Participants with the PER3 haplotype swam 10 seconds (Meve = 10.325 ± 2.087) slower in the morning, on average, while all other genotype combinations swim only 2 seconds slower (Mother = 2.000 ± 1.129; t = 2.905, df = 24, p = 0.008), this represents an 6.5 % slower individual morning performance in athletes with both evening-type polymorphisms (t = 7.26, df = 24, p = 0.013).

Figure 3
Allele and chronotype frequency and chronotype association of the PER3 SNP and VNTR in elite swimmers. A) The PER3 G allele is more frequent in ET (n = 7) than in MT and NT (n = 19) (OR = 6.800, phi = 1.45, p = 0.148). B) Genotypes with a G present have a significantly lower average MEQ score (M = 41.4 ± 4.55) than CC genotypes (M = 47.9 ± 1.11) (t = 13.7, df = 24, p = 0.046). C) The PER3 5 repeat allele is most prevalent in intermediates but does not have a significant association with a specific chronotype (OR = 0.208, phi = 0.28, p = 0.208). D) Participants with a heterozygous VNTR genotype (PER34/5) do not have higher average MEQ scores (M = 47.6 ± 3.47) than participants with the homozygous (PER34/4) genotype (M = 46 ± 8.10; t = 0.545, df = 23, p = 0.591). E) The PER3 ‘evening’ haplotype (a G-allele and PER34/4) is more frequent in ET than in MT and NT (OR = 8.000, phi = 1.63, p = 0.103). F) The average MEQ score is significantly lower in participants with the PER3 eveningness haplotype—individuals homozygous for the 4-repeat VNTR (PER34/4 genotype) and possessing a G allele—than the average of all the other genotypes (Meve = 40 ± 5.583; Mother = 47.86 ± 1.082; t = 5.64, df = 23, p = 0.026).