
The three volumes of Flora of Tunisia were compiled in the 1950s and published over a period of two decades. The first volume (Cuénod et al., 1954) included Lycopodiophytes, Polypodiophytes, Gymnosperms and Monocots, while the next two volumes, published more than twenty years later (Pottier-Alapetite, 1979, 1981), addressed dicotyledons, i.e. basal Angiosperms, Magnoliideae and Eudicots. The compendium has become incomplete and largely obsolete due to numerous changes in nomenclature and taxonomy over the last several decades (e.g., Bowe et al., 2000; APG II, 2003; APG III, 2009; APG IV, 2016; PPG I, 2016). Le Floc’h and collaborators were moved to undertake a synonymic catalogue of Tunisian flora (Le Floc’h & Boulos, 2008; Le Floc’h et al., 2010), a subject that had already elicited renewed interest, such as El Mokni and collaborators’ publications on alien plant species in Tunisia (e.g., El Mokni & El Aouni, 2012; El Mokni & Iamonico, 2018; El Mokni & Hadj Khalifa, 2020; El Mokni, 2021).
Over the course of twenty years of field surveys conducted in Tunisia, we discovered a number of vascular plants that had not been included in the Tunisian Flora of Cuénod et al. (1954) and Pottier-Alapetite (1979, 1981). One of these (Pilularia minuta) was the subject of an in-depth study (Daoud-Bouattour et al., 2011, 2014); the others have only received mentions in works addressing ecology (Ferchichi-Ben Jamaa et al., 2014; Rouissi et al., 2014, 2016) or in botanical catalogues (Ghrabi-Gammar et al., 2009; Daoud-Bouattour et al., 2025), without providing details about localities, dates or diagnostic features. Here we report these discoveries and provide their localities, dates, the names of the collectors and herbarium specimens, and discuss their biogeography and taxonomy.
Since 2006, our Franco-Tunisian team has carried out annual botanical field surveys. The surveys initially focused on the wetland flora of northern Tunisia (e.g., Daoud-Bouattour et al., 2009, 2011; Ghrabi-Gammar et al., 2009; Muller et al., 2010, 2011), but we subsequently extended our research to the entire country (Figure 1) to help update the Le Floc’h et al. (2010) synonymic catalogue.
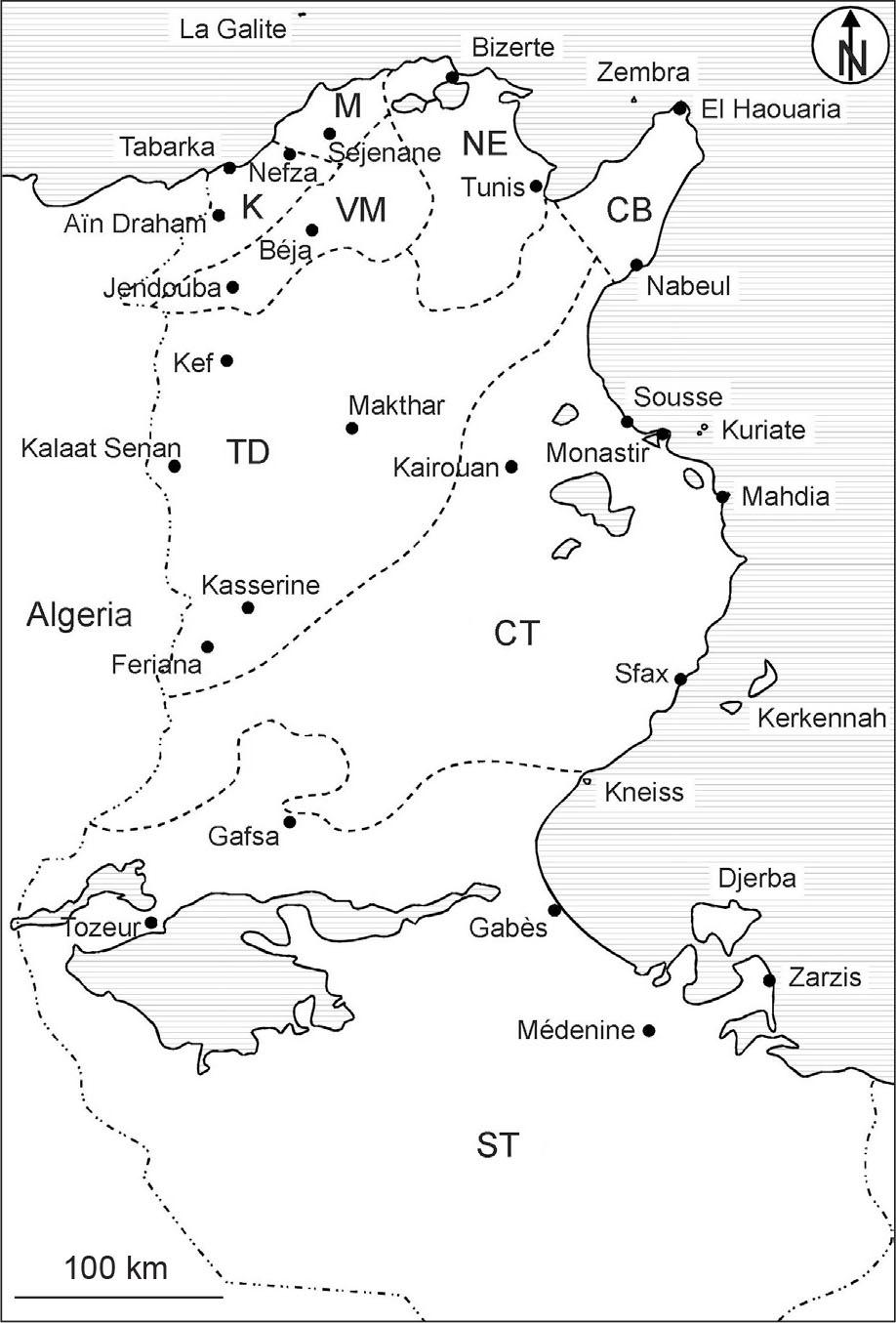
Regions and main cities of Tunisia. CB, Cap Bon; TD, Tunisian Dorsal; K, Kroumiria; M, Mogods; NE, Northeast; VM, Valley of Medjerda; CT, Central Tunisia; ST, Southern Tunisia.
Slika 1: Regije in glavna mesta Tunizije. CB, Cap Bon; TD, Tunijziski Dorsal; K, Kroumiria; M, Mogods; NE, severovzhod; VM, dolina Medjerda; CT, srednja Tunizija; ST, južna Tunizija.
Our collects have been identified according to Maghrebi floras (Cuénod et al., 1954; Quézel & Santa, 1962–1963; Pottier-Alapetite, 1979, 1981; Fennane et al., 1999, 2007, 2014) and related literature that was reviewed in the course of the catalogue update (Daoud-Bouattour et al., 2025). We also consulted ancient herbarium specimens preserved either in Tunisian herbariums (INAT, Faculté des Sciences de Tunis) or at Montpellier University (MPU). For some groups lacking modern, locally adapted keys, we created new ones using criteria given in recent floras of southern Europe: mostly Flora Gallica (Tison & De Foucault, 2014) and Flora Iberica (http://www.floraiberica.es/).
Alisma lanceolatum With., Bot. Arr. Brit. Pl., ed. 3, 2: 362 (1796) (Figure 2a)
≡ Alisma plantago-aquatica L. subsp. lanceolatum (With.) Arcang., Comp. Fl. Ital.: 709 (1882).
Specimens examined: Tunisia, Mogods, Garâa Sejenane (37°03′43″N, 09°13′12″E; 138 m a.s.l.), 21.IV.2008, leg. Amina Daoud-Bouattour; Tunisia, Kroumiria, Nefza (36°58′51″N, 09°04′02″E; 30 m a.s.l.), 23.IX.2023, leg. Imtinen Ben Haj Jilani.
Comments: Cuénod et al. (1954) and Le Floc’h et al. (2010) did not distinguish this taxon from Alisma plantago-aquatica L.. It has been observed since 2006 in Mogods and Kroumiria (northern Tunisia) by A. Daoud-Bouattour, I. Ben Haj Jilani and S. D. Muller (Daoud-Bouattour et al., 2011; Ferchichi-Ben Jamaa et al., 2014; Rouissi et al., 2014, 2016). It has been retained for Tunisia by Dobignard & Chatelain (2010–2013), without details about dates and localities, and then incorporated in the Tunisian flora by most of the online databases.
Alisma lanceolatum develops in shallow waterways, ditches and temporary ponds. It differs from A. plantago-aquatica mainly in its aerial leaf blades with L/l > 3 (vs. < 2.5) and a wedge-shaped base (vs. subcordate), and in petals that are generally pure pink (vs. white or pale pink), straight to bent stems (vs. straight) and occasionally a few achenes with 2 dorsal furrows (vs. always a single furrow). Our investigations only confirm the occurrence of both A. lanceolatum and A. plantagoaquatica in Tunisia. The recent mention of A. gramineum Lej. in the regions of Sejenane and Bizerte where it was erroneously considered an annual species by El Mokni & Domina (2020) could be the result of a confusion with A. lanceolatum.

Some newly recorded taxa: a, Alisma lanceolatum (Garâa Sejenane, 5 May 2007); b, Baldellia repens subsp. repens (Majen el Ma, 27 May 2009); c, Bolboschoenus glaucus (Garâa Sejenane, 4 May 2008); d, Carex echinata (Jbel Ghorra, 10 May 2023); e, Carex panormitana (Beni M’Tir, 5 May 2017); f, Crassula decumbens (Garâa Sejenane, 30 April 2013); g, Crassula helmsii (Garâa Sejenane, 8 May 2014); h, Crassula vaillantii (Garâa Sejenane, 20 April 2008); i, Thelypteris palustris (Ouchtata, 3 May 2013).
Slika 2: Nekateri novo zabeleženi taksoni: a, Alisma lanceolatum (Garâa Sejenane, 5.5.2007); b, Baldellia repens subsp. repens (Majen el Ma, 27.5.2009); c, Bolboschoenus glaucus (Garâa Sejenane, 4.5.2008); d, Carex echinata (Jbel Ghorra, 10.5.2023); e, Carex panormitana (Beni M’Tir, 5.5.2017); f, Crassula decumbens (Garâa Sejenane, 30.4.2013); g, Crassula helmsii (Garâa Sejenane, 8.5.2014); h, Crassula vaillantii (Garâa Sejenane, 20.4.2008); i, Thelypteris palustris (Ouchtata, 3.5.2013).
Baldellia repens (Lam.) van Ooststr. ex Lawalrée, Nouv. Fl. Belg., Luxemb. N. Fr.: 759 (1973) subsp. repens (Figure 2b)
≡ Alisma repens Lam., Encycl. Méth. Bot. 2: 515 (1788).
≡ Alisma ranunculoides L. var. repens (Lam.) Duby, Bot. Gall. 1: 437 (1828).
≡ Baldellia ranunculoides (L.) Parl. subsp. repens (Lam.) Á. Löve & D. Löve, Bot. Not. 114 (1): 49 (1961).
Specimens examined: Tunisia, Kroumiria, Majen el Ma (36°46′52″N, 08°47′24″E; 505 m a.s.l.), 27.V.2009, leg. Serge D. Muller; Tunisia, Mogods, Majen Chitane (37°09′08″N, 09°05′56″E; 150 m a.s.l.), 22.VI.2010, leg. Amina Daoud-Bouattour.
Comments: While Baldellia repens is sometimes considered to be a subsp. of Baldellia ranunculoides (L.) Parl., we retain it at the species level according to Plants of the World online (https://powo.science.kew.org/), Kozlowski & Matthies (2009) and Daoud-Bouattour et al. (2025), because of the clear morphological and ecological differences between both taxa.
Cuénod et al. (1954) did not differentiate this taxon from Baldellia ranunculoides but it was collected (under B. ranunculoides) in 1952 at Majen Azoug (Kroumiria) by G. Pottier-Alapetite (MPU000622). It has been observed in peaty habitats of Mogods and Kroumiria since 2006 by A. Daoud-Bouattour and S. D. Muller. It differs from B. ranunculoides by the presence of stolons (vs. absent), inflorescences with (1)2–6 flowers per whorl (vs. 6–20 flowers), that are often decumbent and rooting at nodes (vs. usually erect) and achenes with whitish papillae (vs. without papillae) that are generally grouped by <20 (vs. >20).
The Baldellia repens subsp. repens, endemic to the Iberian Peninsula, Canary Islands and the Maghreb, is opposed to subsp. cavanillesii, which is endemic to western Europe, and to the subsp. baetica, which is endemic to Andalusia. Kozlowski & Matthies (2009) note that B. repens grows on more acid substrates than B. ranunculoides, which develops in watery areas such as temporary ponds and lake shores. This is consistent with our own observations in Tunisia where B. repens is found in acid peaty habitats.
Red List status: Baldellia repens is listed as ‘Endangered’ in Tunisia (Daoud-Bouattour et al., 2022).
Crassula decumbens Thunb., Prod. Pl. Cap.: 54 (1794) (Figure 2f)
≡ Gomara decumbens (Thunb.) P. V. Heath, Calyx 5: 41 (1995).
Specimens examined: Tunisia, Kroumiria, Majen el Ma (36°46′52″N, 08°47′24″E; 505 m a.s.l.), 27.V.2009, leg. Amina Daoud-Bouattour; Tunisia, Mogods, Garâa Sejenane (37°05′09″N, 09°11′55″E; 100 m a.s.l.), 30.IV.2013, leg. Serge D. Muller.
Comments: Native to Australia and New Zealand, this taxon was collected at Garâa Sejenane, Cap Serrat (Mogods) and Majen el Ma (Kroumiria) by A. Daoud-Bouattour and S. D. Muller (Rouissi et al., 2016).
Crassula helmsii Cockayne, Trans. & Proc. New Zealand Inst. 39: 349 (1907) (Figure 2g)
= Crassula helmsii (Kirk) A. Berger, Nat. Pflanzenfam., ed. 2: 18a: 389 (1930).
Specimen examined: Tunisia, Mogods, Garâa Sejenane (37°05′29″N, 09°10′16″E; 100 m a.s.l.), 30.IV.2013, leg. Serge D. Muller.
Comments: Native to Australia and New Zealand and introduced first in England, this taxon was collected in 2009 in Garâa Sejenane (Mogods; Rouissi et al., 2016) and subsequently in the brackish marshes (sansouïres) of Utique Nouvelle and Ichkeul National Park (Northeast) by A. Daoud-Bouattour and S. D. Muller. Extremely invasive, it seems to be seriously competing with the local flora in Garâa Sejenane, where it constitutes dense carpets.
Crassula vaillantii (Willd.) Roth, Enum. Pl. Phaen. Germ. 1 (1): 992 (1827)
≡ Tillaea vaillantii Willd., Sp. Pl. 1: 720 (1798).
≡ Bulliardia vaillantii (Willd.) DC., Pl. Hist. Succ.: tab. 74 (1801).
Specimen examined: Tunisia, Mogods, Garâa Sejenane (37°05′09″N, 09°11′55″E; 100 m a.s.l.), 20.IV.2008, leg. Serge D. Muller.
Comments: Not mentioned by Pottier-Alapetite (1979), this taxon was discovered in 2006, along with Elatine campylosperma and Pilularia minuta (Ghrabi-Gammar et al., 2009), in Garâa Sejenane by A. Daoud-Bouattour and S. D. Muller. To date, this is its only known Tunisian locality.
The Flora of Tunisia (Pottier-Alapetite, 1979) only mentioned Crassula alata (Viv.) A. Berger, with two varieties today recognised at the species rank under C. alata and C. tillaea Lester-Garland. We thus propose a diagnostic key for the Tunisian Crassula that now comprise five species.
1a. Moss-like plant with very short internodes .............. 2
1b. Plant with long internodes, more or less ramified ... 3
2a. Sessile flowers, generally 3-merous ........................................................... C. tillaea [C. alata var. genuina]
2b. Pedunculate flowers, 4-merous, grouped in cymes of 3–7 flowers; distinctly mucronate sepals and upper leaves ..................... C. alata [C. alata var. trichopoda]
3a. Stem filiform, sparsely branched, with long inter-nodes (monopod); pedicellate flowers, solitary in leaf axils; aquatic form resembling a tiny callitriche and terrestrial form forming dense, tangled clusters ......................................................................... C. helmsii
3b. Stem branched at each node apparently dichotomously (sympod) .................................................... 4
4a. Obtuse leaves; pink flowers with short sepals much smaller than petals ................................. C. vaillantii
4b. Mucronate leaves; white flowers, sometimes tinged with pink, with long, fleshy sepals equal to petals ............................................................... C. decumbens
Bolboschoenus glaucus (Lam.) S. G. Sm., Novon 5: 101 (1995) (Figure 2c)
≡ Scirpus glaucus Lam., Tabl. Encycl. 1: 142 (1791).
= Scirpus tuberosus Desf., Fl. Atlant. 1: 50 (1798).
= Scirpus macrostachys Willd., Enum. Pl. Hort. Berol.: 78 (1809).
= Scirpus maritimus L. var. tuberosus (Desf.) Roem. & Schult., Syst. Veg. 2: 139 (1817).
= Bolboschoenus maritimus (L.) Palla subsp. macrostachys (Willd.) Soják, Čas. Nár. Mus., Odd. Přír. 152: 19 (1983).
Specimen examined: Tunisia, Mogods, Garâa Sejenane (37°05′17″N, 09°12′07″E; 110 m a.s.l.), 26.V.2009, leg. Amina Daoud-Bouattour.
Comments: Scirpus maritimus L., cited by Cuénod et al. (1954), is a complex of three taxa in Europe and the Mediterranean region, which are treated at the species rank in the genus Bolboschoenus by Hroudová et al. (2007). Two of these – Bolboschoenus glaucus and B. maritimus (L.) Palla – are cited in Morocco by Fennane et al. (2014). Hroudová et al. (2007) specify that B. glaucus is a freshwater taxon adapted to temporary habitats and the most thermophilous of the group. B. glaucus differs from B. maritimus in its freshwater habitat (vs. brackish), akenes with epicarp < mesocarp (vs. epicarp ≥ mesocarp) and generally ramified inflorescences (vs. none or few). It was identified in Garâa Sejenane in 2010 by S. D. Muller (Ferchichi-Ben Jamaa et al., 2010; Rouissi et al., 2016).
Carex echinata Murray, Prodr. Stirp. Gött.: 76 (1770) (Figure 2d)
= Carex stellulata Gooden., Trans. Linn. Soc. (London) 2: 144 (1794).
Specimen examined: Tunisia, Kroumiria, Kbar el Hallouf (36°35′14″N, 08°21′33″E; 1005 m a.s.l.), 10.V.2023, leg. Serge D. Muller.
Comments: To date, this northern peatland species was known in North Africa only from Morocco, in the Rif and the High Atlas (Dahlgren & Lassen, 1972; Dobignard & Chatelain, 2010; Fennane et al., 2014). We discovered it in a pastured peatland on the ridge of the Ghorra Mountain range on the Algerian-Tunisian boundary, more than 1,000 km from its nearest Moroccan locality. It is easily distinguishable from other members of the Carex muricata group by its peatland habitat, stellate spikes and null bracts. This unique and isolated population, which may be a relict, is seriously endangered by the overgrazing of the zone.
Carex panormitana Guss., Fl. Sicul. Syn. 2: 575 (1844) (Figure 2e)
? Carex gracilis Curtis var. mucronata Maire, Bull. Soc. Hist. Nat. Afr. N. 33: 90 (1942).
Specimens examined: Tunisia, Kroumiria, Aïn Jmel Falls (36°43′33″N, 08°42′09″E; 600 m a.s.l.), 05.V.2017, 09.IV.2019, leg. Serge D. Muller; Tunisia, Kroumiria, Jbel Khroufa (36°56′22″N, 08°56′56″E; 325 m a.s.l.), 03.V.2023, leg. Serge D. Muller; Tunisia, Kroumiria, Jbel Khroufa (36°56′13″N, 08°56′51″E; 380 m a.s.l.), 04.V.2023, leg. Serge D. Muller; Tunisia, Kroumiria, National Parc Oued Zen (36°46′31″N, 08°47′50″E; 505 m a.s.l.), 06.V.2023, leg. Serge D. Muller; Tunisia, Kroumiria, Statir, National Parc El Feija (36°31′36″N, 08°18′04″E; 930 m a.s.l.), 08.V.2023, leg. Serge D. Muller; Tunisia, Kroumiria, National Parc El Feija (36°29′52″N, 08°19′17″E; 670 m a.s.l.), 09.V.2023, leg. Serge D. Muller; Tunisia, Kroumiria, Oued Bata (36°29′12″N, 08°17′44″E; 730 m a.s.l.), 09.V.2023, leg. Serge D. Muller.
Comments: Cuénod et al. (1954) confused this taxon with Carex acuta L. (syn. C. gracilis Curtis). Carex panormitana was identified in Tunisia by Jiménez-Mejías et al. (2014) on the basis of an 1888 specimen collected by E. Cosson in the Oued Bata near El Feija (Kroumiria). It differs from C. acuta in that it has non-papillose utricles and lamina-less basal sheaths (Jiménez-Mejías et al., 2014). We verified G. Pottier-Alapetite’s herbarium specimen, which had been initially identified as C. acuta (Aïn Soltane, Kroumiria, 1949; MPU024815), and reassigned it to C. panormitana. We also sampled every population observed in Kroumiria (Aïn Soltane, El Feija, Oued Bata, El Merij, Aïn Jmel Falls, Oued Zen Park): all of these belong to C. panormitana, which is obviously the only species of the C. acuta group present in Tunisia. The species is thus endemic to Sardinia, Sicily, and Tunisia and may also occur in Algeria, in El Kala National Park and the mountains of Guelma, where Quézel & Santa (1962–1963) report C. gracilis var. mucronata Maire.
Red List status: Carex panormitana is listed as ‘Vulnerable’ in Tunisia under C. acuta (Daoud-Bouattour et al., 2022).
Isolepis pseudosetacea (Daveau) Carv.-Vasc., Bol. Soc. Brot., ser. 2, 4: 83 (1970)
≡ Scirpus pseudosetaceus Daveau, Bol. Soc. Brot. 9: 85 (1891).
Specimen examined: Tunisia, Cap Bon, Jbel Korbous (36°50′11″N, 10°34′51″E; 175 m a.s.l.), 24.IV.2008, leg. Serge D. Muller.
Comments: Not mentioned by Cuénod et al. (1954), this taxon was collected in 2008 on Jbel Korbous (Cap Bon) by A. Daoud-Bouattour and S. D. Muller. It has never been found again despite being actively sought throughout northern Tunisia. It differs from Isolepis cernua in seed shape and ornamentation (Pignotti 1998): concave (vs. convex) faces covered with papillae < 10 µ (vs. > 10 µ). Other features such as stem and bract length appear to be highly variable in I. cernua and cannot be used to distinguish the two species.
Red List status: Isolepis pseudosetacea is listed as ‘Vulnerable’ in Tunisia (Daoud-Bouattour et al., 2022).
Euphorbia maculata L., Sp. Pl., ed. 1: 455 (1753)
≡ Chamaesyce maculata (L.) Small, Fl. S. E. U. S.: 713, 1333 (1903).
Specimen examined: Tunisia, Mogods, Majen Choucha (37°00′38″N, 09°12′43″E; 445 m a.s.l.), 15.VII.2008, leg. Amina Daoud-Bouattour.
Comments: Pottier-Alapetite (1979) did not know this exotic taxon, which is native to North America. It has been observed twice in Tunisia: once in 2008 in Majen Choucha (Mogods) by A. Daoud-Bouattour and S. D. Muller, and again in 2012, by Véla et al. (in Dobignard & Chatelain, 2013). While its presence at Gammarth could originate from plantations around tourist hotels, its occurrence around the semi-permanent lake of Majen Choucha, located within the cork oak forest of Mogods hills, far away roads, is more surprising. It could have been brought there by the flocks of sheep that frequent the watering hole. It can be confused with E. chamaesyce L. and E. serpens, both of which have similar opposite and stipulated stem leaves. However, it has smooth or adorned seeds with 3–6 regular transverse undulations (vs. irregular granulations or bumps in E. chamaesyce) and hairy ovaries and fruits (vs. glabrous in E. serpens).
Ononis hispida Desf. subsp. arborescens (Desf.) Širj., Fl. Atlant. 2: 149, t. 193 (1798)
≡ Ononis arborescens Desf., Fl. Atlant. 2: 149 (1798).
Specimen examined: Tunisia, Tunisian Dorsal, Jbel Bargou (36°04′40″N, 09°37′06″E; 820 m a.s.l.), 6.V.2024, leg. Serge D. Muller.
Comments: Not mentioned by Pottier-Alapetite (1979–1981), this taxon was previously known from Morocco, Algeria and Libya (Fennane et al., 2007; Dobignard & Chatelain 2010–2013). It was collected in 2024 for the first time in Tunisia on the northern flank of Jbel Bargou (Tunisian Dorsal) by L. Dugerdil and S. D. Muller. The geographic range of this North African endemic thus encompasses Libya and the entire Maghreb. It differs from the typical subspecies by its dense inflorescences (vs. lax) and the fruit, which is as long as the calyx (vs. smaller).
Sphenopus divaricatus (Gouan) Rchb. subsp. permicranthus (Hausskn.) H.Scholz, Willdenowia 19: 410–411 (1990)
≡ Sphenopus gouanii Trin. var. permicranthus Hausskn., Mitt. Thüring. Bot. Ges. 13: 57 (1899).
Specimen examined: Tunisia, South Tunisia, Sebkha Zarkine (33°40′39″N, 10°14′31″E; 24 m a.s.l.), 17. III. 2022, leg. Serge D. Muller.
Comments: Overlooked by Cuénod et al. (1954), this taxon was first reported on the island of Djerba by Lambinon (2000). It was seen again by Médail et al. (2020) in 2014 on the Kerkennah archipelago, and in 2015 on the islands of Djerba and Kneiss. On mainland Tunisia, it was first observed in 2022 in the Sebkha Zarkine (southwestern Tunisia) by S. D. Muller. The subsp. permicranthus differs from the subsp. divaricatus by having smaller lemma (1.0 mm vs. 1.5–1.7 mm) and anthers (0.2 mm vs. 0.4 mm). Le Floc’h et al. (2010) mistakenly listed it as a Tunisian-Libyan endemic species, omitting Scholz (1990), who had reported it from Palestine to Spain.
Red List status: Sphenopus divaricatus subsp. permicranthus is listed as ‘Vulnerable’ in Tunisia (Ghrabi-Gammar et al., 2022a).
Zannichellia obtusifolia Talavera, García-Mur. & H. Smit, Lagascalia 14: 249 (1986) (Figure 3)
Specimen examined: Tunisia, Northeast, Kantaret Binzart (Pont de Bizerte) (36°59′35″N, 10°03′28″E; 08 m a.s.l.), 30.IV.2023, leg. Serge D. Muller.
Comments: Unknown to Cuénod et al. (1954), this taxon was considered with respect to North Africa to be only located in Morocco by Dobignard & Chatelain (2010). In 2023, S. D. Muller found it in an abandoned brackish continental and endorheic channel of the Oued Mejerda (Tunis region), which corresponds to the ecology described by Talavera et al. (1986). It is easily distinguished from all other Zannichellia species by its flat, obtuse leaves having no air canals (vs. thick, acute leaves with two air canals). A visit to the site by A. Daoud-Bouattour in spring 2025 showed its transformation into a dammed canal and the complete disappearance of aquatic vegetation. The species should be sought in similar habitats in northern Tunisia.

Details of Zannichellia obtusifolia (Kantaret Binzart, 30 April 2023). a, complete plant with a detached female flower; b, obtuse, mucronate apex of leaves; c, male flower with a complete stamen composed of an anther carried by a long filament (> 1 cm); d, carpels with membranous infundibuliform stigma. Scale bars represent 1 cm (a, c) and 1 mm (b, d).
Slika 3: Podrobnosti vrste Zannichellia obtusifolia (Kantaret Binzart, 30.4.2023). a, celotna rastlina z ločenim ženskim cvetom; b, topo zaobljen vrh listov; c, moški cvet s celotnim prašnikom, sestavljenim iz prašnic na dolgi prašnični niti (> 1 cm); d, plodni listi z membranasto brazdo v obliki lija. Merilo predstavlja 1 cm (a, c) in 1 mm (b, d).
Thelypteris palustris Schott, Gen. Fil. adnot, tab. 10 (1834) (Figure 2i)
= Acrostichum thelypteris L., Sp. Pl., ed. 2: 1071 (1753).
= Dryopteris thelypteris (L.) A. Gray, Man. Bot. North U. S.: 630 (1848).
Specimens examined: Tunisia, Kroumiria, Ouchtata (36°57′52″N, 09°00′35″E; 25 m a.s.l.), 03.V.2013, leg. Serge D. Muller; Tunisia, Kroumiria, Ouchtata (36°58′12″N, 09°00′42″E; 32 m a.s.l.), 02.V.2023, leg. Serge D. Muller.
Comments: This Northern-Temperate taxon is very rare in North Africa and was not retained for Tunisia by Cuénod et al. (1954), Dobignard & Chatelain (2010–2013) and Le Floc’h et al. (2010). The citation in Tunisia by Roux (2009) is dubious, giving no details about the source of the information.
To our knowledge, this taxon was known in North Africa only from the marshes of Low Loukkos in northwestern Morocco (Fennane et al., 1999) and the El Kala and Guerbès-Senhadja wetland complexes in northeastern Algeria (Quézel & Santa, 1962–1963; Belouahem-Abed et al., 2011). We discovered it in Tunisia in 2013 in a relictual alder swamp on the bank of the Nefza artificial lake, near Ouchtata (Kroumiria). This first population comprises several hundred leaves that developed at the edge of a disturbed alder swamp mixed with Taxodium distichum (L.) Rich. A second population, with fewer than 10 leaves, was found in 2023, approximately 1 km north of the first. This species differs from other Tunisian ferns by virtue of its 2-fold divided leaves, isolated along a subterranean rhizome, and in its peat-forming habitat (mainly alder swamps).
Red List status: Thelypteris palustris is listed as ‘Critically Endangered’ in Tunisia (Ghrabi-Gammar et al., 2022b).
Most recent additions to the vascular flora of Tunisia are alien species that were introduced after the redaction of the Tunisian Flora (Cuénod et al., 1954; Pottier-Alapetite, 1979, 1981) and native taxa described subsequently, such as Limonium spp. (Brullo & Erben, 1989; Suárez-Santiago et al., 2007; Sefi et al., 2020; Ben Haj Jilani et al., 2022), Muscari baeticum Blanca et al. (El Mokni, 2022), Orobanche dominae El Mokni (El Mokni, 2023) or Thymus numidicus Poir. subsp. vagaensis El Mokni & Domina (El Mokni et al., 2023). However, a number of taxa that obviously belong to the native flora of Tunisia but were unknown in Tunisia in the 1950s (Cuénod et al., 1954; Pottier-Alapetite, 1979, 1981) have been discovered in the last decades. In addition to the species mentioned here, we cite Asplenium balearicum Shivas (Médail et al., 2020), A. marinum L. (Muracciole et al., 2010), Bellis perennis L. (Médail et al., 2020), Carthamus creticus L. (Médail et al., 2020), Echinophora spinosa L. (El Mokni, 2020), Neottia nidus-avis (L.) Rich (El Mokni et al., 2010), Ononis minutissima L. (Pavon & Véla, 2011), Pilularia minuta Dur. (Daoud-Bouattour et al., 2009), Solenopsis laurentia (L.) C. Presl subsp. parvula Brullo (Domina & El Mokni, 2012), Sphenopus divaricatus subsp. permicranthus (Lambinon, 2000; Médail et al., 2020), Suaeda spicata (Willd.) Moq. (Sukhorukov et al., 2019), Suaeda splendens (Pourr.) Gren. & Godr. (El Mokni & Debruille, 2021), etc.
There are proportionately fewer additions than those concerning Charophytes (Muller et al., 2017; A. Daoud-Bouattour & S. D. Muller, unpublished data) and Bryophytes (e.g., Hugonnot et al., 2020; Ben Osman et al., 2021a, 2021b, 2022a, 2022b), but Daoud-Bouattour et al. (2025) estimate that they (including newly described taxa) represent almost 10% of the country’s vascular plants which are estimated to include 2543 species and 2842 infraspecific taxa. This clearly points to the fact that Tunisia remains under-researched, even for vascular flora. Nevertheless, the fact that several dozen taxa eluded botanists in the 20th century suggests their extreme rarity in the country, and therefore the great danger of their regional extinction.