
The Danube Delta is a unique territory, Europe’s most prominent and best-preserved delta (Claudino-Sales, 2019). Due to its high biodiversity and the 13 ecosystem services (Gómez-Baggethun et al., 2019) it provides, this unique ecosystem is of global importance (Heath & Evans, 2000). Located in the Pontic biogeographic region, the Danube Delta supports the migration and dispersal of species across the European continent (Tockner & Stanford, 2002; Covaliov et al., 2023). With a biodiversity of over 11.000 taxa, this delta is recognized as one of Europe’s most important biodiversity hotspots. Although it is protected by various national and international agreements, as biosphere reserve, UNESCO World Heritage sites, Ramsar sites, Natura 2000 sites, and national legislation (Claudino-Sales, 2019), it faces impacts of hydrological changes and intensive agriculture, which may threaten the integrity of its natural habitats (Covaliov et al., 2023). These factors also affect soil conditions, which are important for ecosystem stability (Rusu et al., 2025). Thus, soil quality, fertility and health influence ecosystem dynamics and efficiency (Rusu et al., 2024a, b).
From an ecological perspective, the Danube Delta is a unique complex of ecosystems that hydrological factors, such as water level, flow measurements, riverbed profiles, and average water velocity, significantly influence. These factors can have an important role in shaping vegetation. Therefore, conducting a case study to ascertain the degree of influence of these factors on vegetation cover in some natural islands in the Danube Delta is a valuable strategy. Hanganu et al. (2017) studied the effects of multiple pressures, including hydrological factors, on the lower Danube basin, where they showed insights into the relationships between hydrological conditions and the overall ecological status of water bodies.
The construction of dams, modification of natural watercourses, and other anthropogenic interventions intensify the loss of riparian habitats in the Danube Delta. Due to these changes, there has been a severe alteration in the connectivity of aquatic and terrestrial ecosystems. This has affected the functioning of ecosystems and biodiversity (Schneider, 2011). Also, the construction of dams can limit the natural flow of sediments and water, which are essential for maintaining the habitats and vegetation typical of the delta (Gâştescu & Ştiucă, 2006). Due to these practices, many wetlands have been transformed into agricultural land. In this manner, the natural areas for the flora and fauna specific to the delta have been significantly reduced.
In addition, research in other deltas, such as the Mississippi Delta (Day et al., 2007), the Mekong Delta (Le et al., 2007), and the Nile Delta (Frihy, 2003), shows the vital role of vegetation in supporting ecosystem services in terms of climate change impacts and biodiversity. Because of their vulnerability to anthropogenic impacts and climate change, these deltaic ecosystems have been the subject of extensive ecological studies. Vegetation in deltaic ecosystems is vulnerable to changes in hydrology, nutrient availability, and sediment dynamics (Temmerman et al., 2013).
This study aims to analyze the habitats and hydrological and morphological factors of seven islands in the Danube Delta. The objectives of the study were (1) to identify habitats and plant associations, (2) to record hydrological measurements, and (3) to analyze the dynamics of the Lower Danube islands. This study can contribute to understanding the dynamics of islands in the Danube Delta, an area with remarkable biodiversity vulnerable to climatic variations and anthropogenic impact. The study’s results can also help implement conservation measures in the context of restoring affected habitats.
The study was conducted on seven islands of the Danube Delta (Figure 1) in 2023 and 2024. These islands are distributed as follows: one island located (A1) on the Chilia branch with an area of 10.91 ha; three islands located (A2, A3, and A4) in the vicinity of Isaccea port, between Cotul Pisicii and Ceatal Izmail which have an area of 69 ha; and three islands (A5a, A5b, A6) located on the Sf. Gheorghe branch with an area of 32.54 ha.

Study areas.
Slika 1: Preučevana območja.
The size of the plots was 100 m2 (Cristea et al., 2004). The vegetation was classified using the Agglomerative Hierarchical Clustering method (ß-flexible method, β = −0.25 and Bray-Curtis dissimilarity) based on square root transformed data. The initial data were represented by average percentage values corresponding to the Braun-Blanquet (1964) cover-abundance scale (Cristea et al., 2004): r (0.05%), + (0.5%), 1 (5%), 2 (17.5%), 3 (37.5%), 4 (62.5%), and 5 (87.5%).
The dendrogram was made in the GINKGO software (Bouxin, 2005), and the optimal number of clusters was identified based on the average Silhouette index (Rousseeuw, 1987). The synoptic table was obtained from the Juice program version 7.1 (Tichý, 2002).
The coenotaxonomic classification of plant communities is defined according to Chifu et al. (2014a, 2014b), Chifu & Irimia (2014). For each cluster, plant species were presented according to the phi-coefficient value (Chytrý et al. 2002; Tichý & Chytrý 2006). A fidelity threshold of phi = 0.4 and Fisher’s exact test (p < 0.05) were used.
The nomenclature of plant species followed Euro+Med (2024), and the nomenclature of associations followed Chifu et al. (2014a, 2014b), and Chifu & Irimia (2014). The nomenclature of higher syntaxa followed Mucina et al. (2016). Habitat types were classified using the EUNIS habitat classification expert system (Chytrý et al., 2020).
In the study, bathymetric measurements were made using the SonTek RiverSurveyor M9 equipment, mounted on a HydroBoard II. The equipment uses acoustic pulses to determine water velocity, depth, flow, and riverbed elevations based on advanced technology. Multiple acoustic frequencies ensure accurate measurements at shallow and deep depths, even while moving. Data were collected on longitudinal and transverse sections at distances of 100–150 m and in critical areas at 50 m to obtain a detailed representation of the riverbed profile. The equipment features GPS RTK for precise real-time positioning (±3 cm), transmitting and correcting data via Bluetooth. The equipment’s compass was calibrated to compensate for magnetic interference, and the sensors were positioned upstream of the boat’s engine to avoid the influences of vibrations and propeller-generated currents (Sočuvka, 2024). For accuracy, additional topographic measurements were also performed using the Spectra Precision SP80 GPS and the LEICA NA 524 optical level to validate the water level. The collected data provides detailed information on the bathymetric profile, current velocity, and discharge of the riverbed’s cross-sections, ensuring a general picture of the hydrological parameters of the studied area. Violin plots of hydrological factors were produced in R Statistical Software (v4.1.4; R Core Team 2024) via ‘ggplot2’ v.3.5.1 (Wickham, 2016).
The analysis of lower Danube River Island dynamics within this study is conducted through morphometric methods based on high-resolution remote sensing techniques. This approach allows for the precise quantification of changes, whether growth, erosion, or migration, through the detailed comparison of data from various periods. The Danube River islands within this study constitute a complex natural laboratory for the investigation of fluvial morphodynamic processes, reflecting the dynamic interaction between hydrological forces and sediment transport (Leopold & Wolman, 1957; Bondar et al., 1992; Church & Ferguson, 2015). The comparative analysis, utilizing morphometric methods with data derived from ANCPI orthophotoplans (2011 & 2021) and Airbus imagery (2006–2020), reveals a significant variability in their spatio-temporal evolution.
In the plots made in the seven natural islands of the Danube Delta (20 relevés), 98 taxa were identified.
Four EUNIS habitat types are represented in: T11 Temperate Salix and Populus riparian forest; V38 Dry perennial anthropogenic herbaceous vegetation; Q51 Tall-helophyte bed; and Q54 Inland saline or brackish helophyte.
The analyzed vegetation is classified into four phytocoenological classes: Artemisietea vulgaris, Lemnetea, Phragmito-Magnocaricetea, and Salicetea purpureae. In the seven natural islands analyzed, six associations and one sub-associations were identified, including dominant and characteristic species for each habitat.
Artemisietea vulgaris Lohmeyer et al. in Tx. ex von Rochow 1951
Agropyretalia intermedio-repentis T. Müller et Görs 1969
Convolvulo arvensis-Agropyrion repentis Görs 1967
Convolvulo-Agropyretum repentis Felföldy 1943
Lemnetea O. de Bolòs et Masclans 1955
Lemnetalia minoris O. de Bolòs et Masclans 1955
Lemnion minoris O. de Bolòs et Masclans 1955
Lemno-Azolletum filiculoides Br.-Bl. 1952
Phragmito-Magnocaricetea Klika in Klika et Novák 1941
Oenanthetalia aquaticae Hejný ex Balátová-Tuláčková et al. 1993
Eleocharito palustris-Sagittarion sagittifoliae Passarge 1964
Bolboschoenetum maritimi Eggler 1933
Phragmitetalia Koch 1926
Phragmition communis Koch 1926
Glycerietum maximae Hueck 1931
Salicetea purpureae Moor 1958
Salicetalia purpureae Moor 1958
Salicion albae Soó 1951
Salicetum albae Issler 1926
amorphoetosum fruticosae Morariu et Danciu 1970
Rubo caesii-Salicetum albae Doniță et Dihoru 1961 em. Doniță et al. 1966
The data obtained from the cluster analysis were presented as a dendrogram (Figure 2) and a synoptic table (Table 1). The vegetation was grouped into seven clusters and assigned to different syntax groups described in the literature.
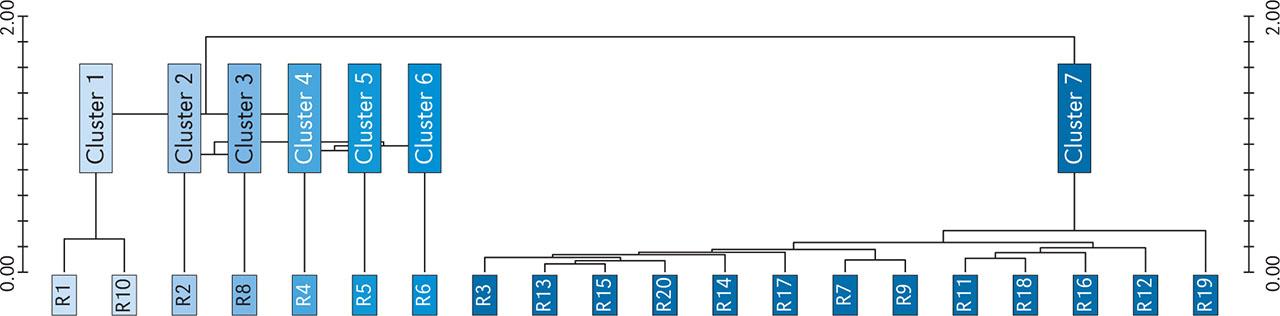
Dendrogram of vegetation data obtained from cluster analysis.
Slika 2: Dendrogram vegetacijskih podatkov, narejen s klastrsko analizo.
Synoptic table of the percentage frequencies of plant species in the analyzed communities.
Tabela 1: Sinoptična tabela preučevanih združb in pogostosti rastlinskih vrst v odstotkih.
| No. cluster | Phi | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| No. relevés | 2 | 1 | 1 | 1 | 1 | 1 | 13 | |
| Cluster 1: Salicetum albae: amorphoetosum fruticosae | ||||||||
| Brachypodium sylvaticum (Huds.) P. Beauv. | 0.688 | 50 | . | . | . | . | . | . |
| Digitaria sanguinalis (L.) Scop. | 0.688 | 50 | . | . | . | . | . | . |
| Fumaria officinalis L. | 0.688 | 50 | . | . | . | . | . | . |
| Helianthemum nummularium (L.) Mill. | 0.688 | 50 | . | . | . | . | . | . |
| Aristolochia clematitis L. | 0.577 | 100 | 100 | . | . | . | . | 15 |
| Vitis vinifera L. | 0.454 | 100 | 100 | 100 | . | . | . | 23 |
| Fraxinus pennsylvanica Marshall | 0.444 | 50 | 100 | . | . | . | . | . |
| Glechoma hederacea L. | 0.444 | 50 | . | . | . | . | . | 8 |
| Potentilla reptans L. | 0.444 | 50 | . | . | . | . | . | 8 |
| Cluster 2: Convolvulo-Agropyretum repentis | ||||||||
| Urtica dioica L. | 0.793 | . | 100 | 100 | . | . | . | 8 |
| Dactylis glomerata L. | 0.688 | . | 100 | . | . | . | . | . |
| Drymochloa sylvatica (Pollich) Holub | 0.688 | . | 100 | . | . | . | . | . |
| Elytrigia repens (L.) Nevski | 0.688 | . | 100 | . | . | . | . | . |
| Hordeum murinum L. | 0.688 | . | 100 | . | . | . | . | . |
| Lolium perenne L. | 0.688 | . | 100 | . | . | . | . | . |
| Potentilla recta L. | 0.688 | . | 100 | . | . | . | . | . |
| Fraxinus angustifolia Vahl | 0.577 | 50 | 100 | 100 | . | . | . | 15 |
| Acer negundo L. | 0.509 | 50 | 100 | 100 | . | . | . | 23 |
| Cirsium arvense (L.) Scop. | 0.454 | 50 | 100 | 100 | 100 | . | . | 23 |
| Sonchus asper (L.) Hill | 0.454 | 50 | 100 | 100 | . | . | . | 31 |
| Vitis vinifera L. | 0.454 | 100 | 100 | 100 | . | . | . | 23 |
| Agrostis capillaris L. | 0.444 | . | 100 | . | . | . | . | 8 |
| Bromus hordeaceus L. | 0.444 | . | 100 | . | . | . | . | 8 |
| Fraxinus pennsylvanica Marshall | 0.444 | 50 | 100 | . | . | . | . | . |
| Cluster 3: Rubo caesii-Salicetum albae | ||||||||
| Chenopodium album L. | 0.577 | . | . | 100 | . | . | . | 31 |
| Cluster 4: Bolboschoenetum maritimi | ||||||||
| Bolboschoenus maritimus (L.) Palla | 1.000 | . | . | . | 100 | . | . | . |
| Carduus acanthoides L. | 1.000 | . | . | . | 100 | . | . | . |
| Carex elata All. | 1.000 | . | . | . | 100 | . | . | . |
| Phalaroides arundinacea (L.) Rauschert | 1.000 | . | . | . | 100 | . | . | . |
| Polygonum aviculare L. | 1.000 | . | . | . | 100 | . | . | . |
| Rorippa austriaca (Crantz) Besser | 1.000 | . | . | . | 100 | . | . | . |
| Althaea officinalis L. | 0.688 | . | . | . | 100 | . | . | 8 |
| Convolvulus arvensis L. | 0.688 | . | . | . | 100 | . | . | 8 |
| Myosotis scorpioides L. | 0.546 | . | . | . | 100 | . | . | 15 |
| Plantago major L. | 0.546 | . | . | 100 | 100 | . | . | 8 |
| Cluster 5: Glycerietum maximae | ||||||||
| Azolla filiculoides Lam. | 0.459 | . | . | . | 100 | 100 | 100 | 8 |
| Glyceria maxima (Hartm.) Holmb. | 0.459 | . | . | . | 100 | 100 | . | 15 |
| Cluster 6: Lemno-Azolletum filiculoides | ||||||||
| Potamogeton natans L. | 1.000 | . | . | . | . | . | 100 | . |
| Azolla filiculoides Lam. | 0.459 | . | . | . | 100 | 100 | 100 | 8 |
| Cluster 7: Salicetum albae | ||||||||
| Phragmites australis (Cav.) Steud. | 0.798 | . | . | . | 100 | . | . | 62 |
| Stachys palustris L. | 0.698 | 50 | . | . | . | . | . | 54 |
| Calamagrostis arundinacea (L.) Roth | 0.638 | . | . | . | . | . | . | 38 |
| Salix cinerea L. | 0.638 | . | . | . | . | . | . | 38 |
| Solanum dulcamara L. | 0.596 | 50 | . | 100 | . | . | . | 85 |
| Jacobaea paludosa (L.) G. Gaertn. & al. | 0.553 | . | . | . | . | . | . | 31 |
| Populus alba L. | 0.553 | . | . | . | . | . | . | 31 |
| Sium latifolium L. | 0.553 | . | . | . | . | . | . | 31 |
| Typha angustifolia L. | 0.553 | . | . | . | . | . | . | 31 |
| Symphytum officinale L. | 0.504 | 50 | . | . | . | . | . | 38 |
| Alisma plantago-aquatica L. | 0.464 | . | . | . | . | . | . | 23 |
| Lemna minor L. | 0.464 | . | . | . | . | . | . | 23 |
| Lemna minuta Kunth | 0.464 | . | . | . | . | . | . | 23 |
| Lycopus europaeus L. | 0.414 | 50 | . | . | 100 | . | . | 69 |
Structure and composition of plant community: Vegetation cover ranged from 42% to 67%, and the number of species /100 m2 ranged from 10 to 25. Amorpha fruticosa had a cover of 37.5%, and Salix alba had a cover ranged from 0.5% to 17.5%. The floristic composition of this cluster was included in the class Salicetea purpureae, the order Salicetalia purpureae, characteristic of communities in floodplain areas, and the alliance Salicion albae. These relevés are part of the T11 habitat, according to the EUNIS classification.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, average temperatures ranged from −0.7 °C in January to 22 °C in July. These habitats thrive in conditions of high humidity and periodic flooding. The annual mean precipitation ranged from 443 to 493 mm, and the annual mean temperature was above 10.8 °C.
Distribution: The two relevés included in this cluster, were distributed on two islands (A2 and A3) from Isaccea.
Structure and composition of plant community: The dominant species was Elytrigia repens, which had a cover of 62.5%. The vegetation cover was 73%, and the number of species /100 m2 was 21. The vegetation belongs to the class Artemisietea vulgaris, the order Agropyretalia intermediorepentis, and the alliance Convolvulo arvensis-Agropyrion repentis, at the phytocoenological level. This relevé is part of the V38 habitat, according to the EUNIS classification.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, and average temperatures from −0.7 °C in January to 22 °C in July. Water level ranged from 2.8 m in spring, 2.7 m in summer, 3.3 m in autumn to 4.1 m in winter. Regarding flow measurements, it ranged from 6059 m3/s in spring, 5883 m3/s in summer, 7179 m3/s in autumn, to 7846 m3/s in winter. Regarding average water velocity, it ranged from 0.80 m/s in spring, 0.78 m/s in summer, 0.95 m/s in autumn, to 1.03 m/s in winter. Riverbed profiles ranged from −6.52 m in spring, −6.6 m in summer, −6.68 m in autumn to −6.5 m in winter.
Distribution: A relevé was included in this cluster, which was distributed on one island (A4) from Isaccea.
Structure and composition of plant community: The dominant species was Rubus caesius, with a cover of 37.5%. The vegetation cover was 48%, and the number of species /100 m2 was 21. The vegetation was classified within the class Salicetea purpureae, the order Salicetalia purpureae, and the alliance Salicion albae. This relevé is part of the T11 habitat, according to the EUNIS classification.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, and average temperatures from −0.7 °C in January to 22 °C in July. Water level ranged from 2.8 m in spring, 2.7 m in summer, 3.3 m in autumn to 4.1 m in winter. Regarding flow measurements, it ranged from 6059 m3/s in spring, 5883 m3/s in summer, 7179 m3/s in autumn, to 7846 m3/s in winter. Regarding average water velocity, it ranged from 0.80 m/s in spring, 0.78 m/s in summer, 0.95 m/s in autumn, to 1.03 m/s in winter. Riverbed profiles ranged from −6.52 m in spring, −6.6 m in summer, −6.68 m in autumn to −6.5 m in winter.
Distribution: A relevé was included in this cluster, which was distributed on one island (A4) from Isaccea.
Structure and composition of plant community: The dominant species is Bolboschoenus maritimus, which had a cover of 37.5%. Vegetation cover was 49%, and the number of species /100 m2 was 23. The EUNIS habitat is Q54 Inland saline or brackish helophyte bed. The vegetation is part of the class Phragmito-Magnocaricetea, the order Bolboschoenetalia maritimi, and the alliance Scirpion maritimi.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, and average temperatures from −0.7 °C in January to 22 °C in July. Water level ranged from 2.8 m in spring, 2.7 m in summer, 3.3 m in autumn to 4.1 m in winter. Regarding flow measurements, it ranged from 6059 m3/s in spring, 5883 m3/s in summer, 7179 m3/s in autumn, to 7846 m3/s in winter. Regarding average water velocity, it ranged from 0.80 m/s in spring, 0.78 m/s in summer, 0.95 m/s in autumn, to 1.03 m/s in winter. Riverbed profiles ranged from −6.52 m in spring, −6.6 m in summer, −6.68 m in autumn to −6.5 in winter.
Distribution: A relevé from one of the islands (A4) from Isaccea was included in this cluster.
Structure and composition of plant community: Glyceria maxima was the dominant species and had a cover of 62.5%. Vegetation cover was 65%, and the number of species /100 m2 was 5. The EUNIS habitat is Q51 Tall-helophyte bed. The vegetation belongs to the class Phragmito-Magnocaricetea, the order Phragmitetalia, and the alliance Phragmition communis.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, and average temperatures from −0.7 °C in January to 22 °C in July. Water level ranged from 2.8 m in spring, 2.7 m in summer, 3.3 m in autumn to 4.1 m in winter. Regarding flow measurements, it ranged from 6059 m3/s in spring, 5883 m3/s in summer, 7179 m3/s in autumn, to 7846 m3/s in winter. Regarding average water velocity, it ranged from 0.80 m/s in spring, 0.78 m/s in summer, 0.95 m/s in autumn, to 1.03 m/s in winter. Riverbed profiles ranged from −6.52 m in spring, −6.6 m in summer, −6.68 m in autumn to −6.5 m in winter.
Distribution: A relevé was included in this cluster found on one island (A4) from Isaccea.
Structure and composition of plant community: Azolla filiculoides was the dominant species and had a cover of 37.5%. The vegetation cover was 39% and the number of species /100 m2 was 3. The EUNIS habitat is C1.3 Permanent eutrophic lakes, ponds and pools. This relevé belongs to the class Lemnetea, the order Lemnetalia minoris, and the alliance Lemnion minoris.
Ecology: Average precipitation ranged from 26 mm in January to 57 mm in June, and average temperatures from −0.7 °C in January to 22 °C in July. Water level ranged from 2.8 m in spring, 2.7 m in summer, 3.3 m in autumn to 4.1 m in winter. Regarding flow measurements, it ranged from 6059 m3/s in spring, 5883 m3/s in summer, 7179 m3/s in autumn, to 7846 m3/s in winter. Regarding average water velocity, it ranged from 0.80 m/s in spring, 0.78 m/s in summer, 0.95 m/s in autumn, to 1.03 m/s in winter. Riverbed profiles ranged from −6.52 m in spring, −6.6 m in summer, −6.68 m in autumn to −6.5 m in winter.
Distribution: A relevé was included in this cluster, which was distributed on one island (A4) from Isaccea.
Structure and composition of plant community: Vegetation cover ranged from 41% to 60%, and the number of species /100 m2 varied from 8 to 31. Salix alba had a cover of 37.5%. The vegetation corresponds to the class Salicetea purpureae, the order Salicetalia purpureae, and the alliance Salicion albae. These relevés are part of the T11 habitat, according to the EUNIS classification.
Ecology: Average precipitation ranged from 23 mm in January to 49 mm in June, and average temperatures from 0.2 °C in January to 22 °C in July. Water level ranged from 2.1 m in spring, 2.07 m in summer, 2.7 m in autumn to 3.3 m in winter. Regarding flow measurements, it ranged from 4116 m3/s in spring, 3978 m3/s in summer, 4996 m3/s in autumn, to 5568 m3/s in winter. Regarding average water velocity, it ranged from 0.68 m/s in spring, 0.65 m/s in summer, 0.87 m/s in autumn, to 1.00 m/s in winter. Riverbed profiles ranged from −5.27 m in spring, −5.33 m in summer, −5.28 m in autumn to −6.5 m in winter.
Distribution: In this cluster, 13 relevés were included, distributed in Isaccea (A4), Chilia (A1) and Sf. Gheorghe (A5a, A5b, and A6).
Water level (Figure 3; Table 2). The water level measured at different points showed significant seasonal fluctuations. At Isaccea, levels remained relatively stable in spring and summer (approximately 2.86 m in spring and 2.73 m in summer). However, it increased considerably in autumn and winter, reaching 4.12 m in winter. The same trend was observed at locations such as Dunavăț and Dranov, where winter values were higher than summer values (for example, at Dranov, the water level increased from 0.87 m in summer to 2.00 m in winter).
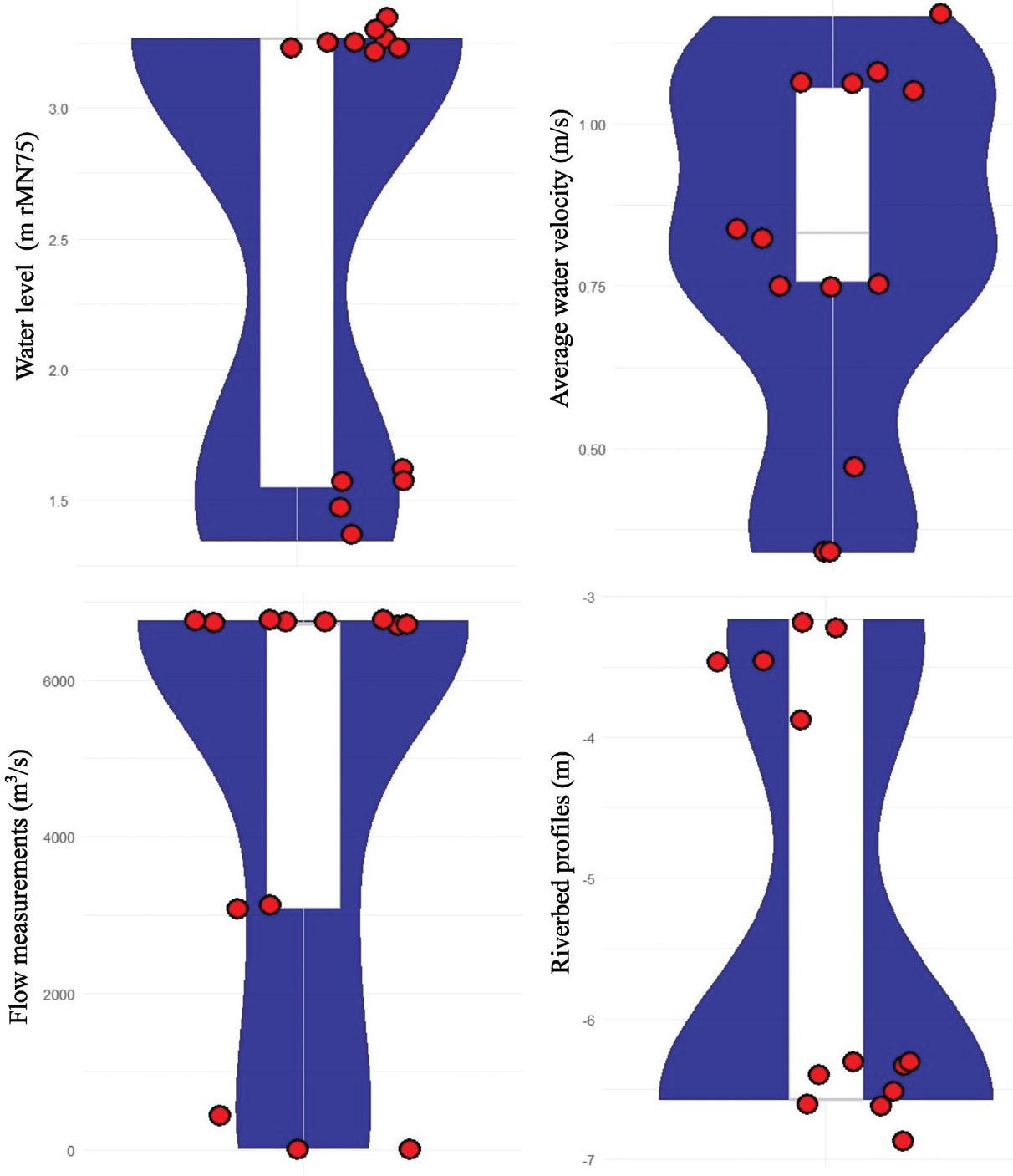
Variation of hydrological measurements in the analyzed areas.
Slika 3: Variacija hidroloških meritev na preučevanih območjih.
Results of hydrological measurements.
Tabela 2: Rezultati hidroloških meritev.
| Location | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|
| Water level measured in the field (m rMN75) | Isaccea | 2.86 | 2.73 | 3.36 | 4.12 |
| 2.86 | 2.73 | 3.36 | 4.12 | ||
| 2.86 | 2.73 | 3.36 | 4.12 | ||
| Dunavăț | 1.17 | 1.07 | 1.74 | 2.23 | |
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| 1.17 | 1.07 | 1.74 | 2.23 | ||
| Dranov | 0.97 | 0.87 | 1.54 | 2.00 | |
| 0.97 | 0.87 | 1.54 | 2.00 | ||
| 0.97 | 0.87 | 1.54 | 2.00 | ||
| 0.97 | 0.87 | 1.54 | 2.00 | ||
| 0.97 | 0.87 | 1.54 | 2.00 | ||
| Flow measurements (m3/s) | Isaccea | 6052 | 5876 | 7171 | 7859 |
| 6034 | 5858 | 7149 | 7793 | ||
| 6083 | 5906 | 7207 | 7867 | ||
| Dunavăț | 17 | 16 | 26 | 32 | |
| 10 | 9 | 15 | 19 | ||
| 15 | 14 | 23 | 29 | ||
| 18 | 17 | 27 | 35 | ||
| 16 | 15 | 24 | 31 | ||
| 2314 | 2143 | 3463 | 4438 | ||
| 2355 | 2181 | 3524 | 4517 | ||
| Dranov | 574 | 508 | 825 | 1089 | |
| 356 | 315 | 517 | 637 | ||
| 344 | 304 | 498 | 612 | ||
| 368 | 326 | 537 | 662 | ||
| 23 | 20 | 33 | 37 | ||
| Average water velocity (m/s) | Isaccea | 0.68 | 0.66 | 0.81 | 0.88 |
| 0.75 | 0.73 | 0.89 | 0.96 | ||
| 0.95 | 0.92 | 1.13 | 1.22 | ||
| Dunavăț | 0.36 | 0.34 | 0.55 | 0.67 | |
| 0.32 | 0.3 | 0.49 | 0.62 | ||
| 0.19 | 0.18 | 0.29 | 0.37 | ||
| 0.29 | 0.27 | 0.43 | 0.55 | ||
| 0.11 | 0.1 | 0.16 | 0.21 | ||
| 0.81 | 0.75 | 1.21 | 1.55 | ||
| 0.87 | 0.81 | 1.31 | 1.67 | ||
| Dranov | 0.14 | 0.13 | 0.2 | 0.27 | |
| 0.47 | 0.43 | 0.69 | 0.85 | ||
| 0.51 | 0.46 | 0.75 | 0.91 | ||
| 0.49 | 0.45 | 0.73 | 0.89 | ||
| 0.22 | 0.20 | 0.32 | 0.36 | ||
| Riverbed profiles (m) | Isaccea | −6.52 | −6.60 | −6.68 | −6.50 |
| Dunavăț | −3.12 | −3.15 | −3.19 | −3.19 | |
| Dranov | −3.87 | −3.99 | −3.95 | −3.97 | |
Water velocity (Figure 3; Table 2). The water velocity had prevailing values between 0.3 and 0.9 m/s throughout the year, suggesting a consistency in moderate flow areas. The highest velocities were recorded in autumn and winter, reflecting the water accumulations in these periods. At Isaccea, the water velocity increased constantly throughout the year, from spring to winter, which indicates an intensification of the water flow, especially during the rainy season and snowmelt. Thus, at Isaccea, the water velocity was 0.68 m/s in spring, decreased slightly in summer to 0.66 m/s, but showed a significant increase in autumn (0.81 m/s) and reached 0.88 m/s in winter. In Dunavăț and Dranov, the water velocity was lower but followed the same trend. At Dranov, the water velocity increased from 0.13 m/s in summer to 0.27 m/s in winter.
Flow measurements (Figure 3; Table 2). At Isaccea, the water flow was 6052 m3/s in spring and decreased slightly in summer to 5876 m3/s. It increased considerably in autumn (7171 m3/s) and reached a maximum of 7859 m3/s in winter. At Dranov, the flow was significantly lower but followed a similar trend, increasing from 508 m3/s in summer to 1089 m3/s in winter.
Riverbed profiles (Figure 3; Table 2). Bathymetric measurements indicate the elevation of the riverbed, helping to assess changes in its structure. At Isaccea, Dunavăț, and Dranov, the values were relatively constant throughout the year, with slight variations, suggesting the stability of the riverbed in these areas. At Isaccea, the depth of the riverbed varied very little from −6.52 m in spring to −6.50 m in winter, indicating sediment accumulation. At Dranov, the values showed a linear trend, ranging from −3.87 m in spring to −3.99 m in winter. The intermediate range remained relatively constant in all seasons, varying very little between −3.87 and −3.99 m.
The Danube Delta, a dynamic fluvial-marine environment, showcases a complex interplay of erosion and accumulation, influencing the morphology of its numerous islands. An analysis of seven islands, reveals distinct patterns of sediment redistribution. Particularly, all islands exhibit a net positive accumulation, a testament to the delta’s robust depositional regime. However, the magnitude of this accumulation varies significantly, highlighting the influence of local hydrodynamics and island positioning.
The recent emergence of A5a and A5b islands, around 2010, along with the substantial accumulation observed in A1, A2, and A3, suggests a modification of this regime, most likely associated with a significant change in the amplitude of the water level and river discharge (Figure 4). Principally, decreases in flow promote rapid sediment deposition. These hydrological events favored rapid sediment accumulation, leading to the formation and expansion of the islands. In this respect, island A5a demonstrated a spectacular area increase, from 3.64 to 18 ha in 10 years, surpassing the expansion rate of island A5b (Figure 4), while A4 demonstrates a dual pattern of erosion and sedimentation. This pronounced accumulation may be attributed to its location within the rectified Perivolovca meander, where altered flow dynamics promote enhanced sediment deposition. In contrast, A2 exhibits the lowest accumulation, only 0.28 ha, concentrated on its right bank, shielded from the direct river discharge. This island, however, experiences 0.88 ha of erosion on its left bank, facing the primary river flow, illustrating a localized disparity in morphometric processes. A1, initially showing exclusive accumulation, has since migrated and become a definitive part of the Danube’s right bank. A2 and A3 display both erosion and accumulation, with accumulation exceeding erosion. This variability highlights the local influences of flow conditions and sediment availability.
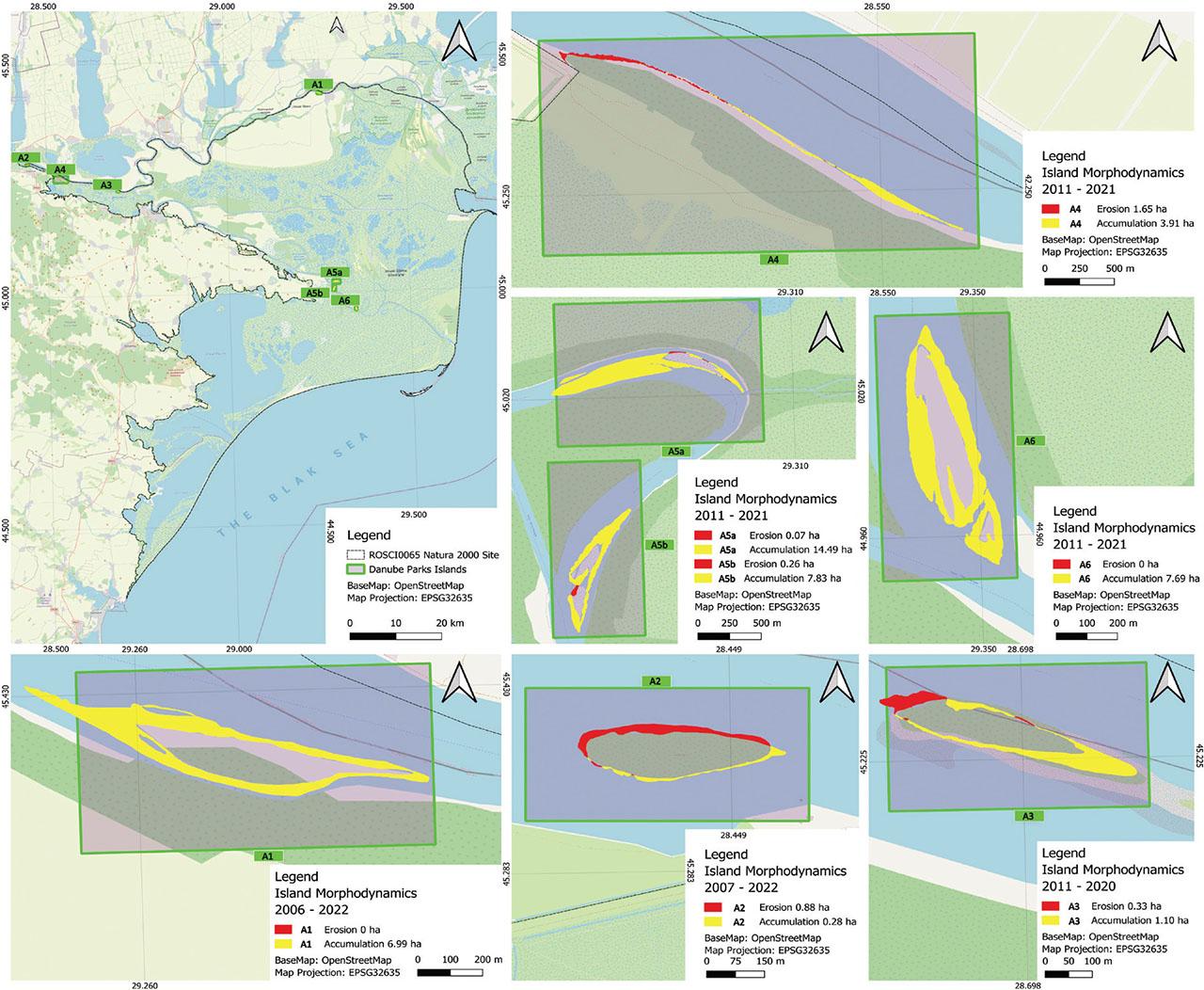
The location of the study’s islands along the lower Danube River – islands morphodynamics.
Slika 4: Lokacija preučevanih otokov ob spodnjem teku reke Donave – morfodinamika otokov.
The tendencies of sedimentation and stabilization are observable in the advancement of islands A5a and A5b toward the river banks, and in the land-tied evolution of islands A4 and A1 to the Danube’s right bank. A5b, also located within the rectified Perivolovca meander, demonstrates a significant accumulation of 7.83 ha, further supporting the fact that meander rectification can amplify sediment deposition. A6, situated in the rectified Dranov 1 meander, displays a similar trend, with 7.69 ha of accumulation and no recorded erosion, reinforcing the influence of altered channel geometry on sediment dynamics.
Although less widespread than accumulation, erosion remains a noteworthy factor. The dual dynamics, characterized by upstream erosion, where river erosion is strongest, and downstream sedimentation, as observed in island A4. Thus, the first half of island A4 was eroded of 1.65 ha in its upstream over 10 years, while the downstream half exhibited emergent zones totaling 3.9 ha within the same period (Figure 4). Conversely, A5a registers minimal erosion, a mere 0.07 ha. A3 and A2 show intermediate erosion values, 0.33 and 0.88 ha respectively, indicating localized erosion patterns influenced by their position relative to the river’s flow. A5b displays 0.26 ha of erosion. A1 and A6 register zero erosion. The varied sedimentation rates, again, evidenced by the spectacular area growth of island A5a, suggest an intense sediment transport dynamic by the river, exceeding the expansion rate of island A5b.
Furthermore, the land-tied evolution observed in A1 and A4 underscores the dynamic nature of island development. A1, exhibiting 6.99 ha of accumulation, is undergoing land-tied evolution towards the right bank, indicating a process of gradual connection to the mainland. Similarly, A4, with 3.91 ha of accumulation in its downstream segment, displays land-tied evolution towards the right bank and underwent significant changes along its left bank at fluctuating rates being exposed to the river’s discharge. These processes are indicative of the delta’s continuous shaping by sedimentary processes. The predominant upstream extension of island A5a indicates progressive sedimentation of the Perivolovca meander, while lateral extension and bank land-tied evolution, observed in island A5b, reflect lateral deposition processes (Figure 4).
The Danube Delta is a dynamic territory and complex of ecosystems with an area of approximately 5,800 km2 (Hanganu et al., 2017). It is located at the confluence of the Danube River and the Black Sea, which gives it a special ecological importance (Catianis et al., 2016).
In the analysed islands, the communities of Salix alba and Amorpha fruticosa predominate. According to Sitzia et al. (2018), S. alba has an important role in the ecology of the Danube Delta. Thus, through its root structure, S. alba can increase soil resistance and provide additional support through bioengineered structures (Zhang et al., 2013). At the same time, it has been shown that at high phosphorus concentrations (Hrynkiewicz et al., 2015), a decrease in ectomycorrhizal colonization of Salix viminalis and Betula pendula seedlings was recorded. In contrast, Amorpha fruticosa can fix nitrogen and enrich the soil with nitrogen compounds, which causes changes in natural habitat conditions (Gudžinskas & Žalneravičius, 2015). This species also has significant regeneration ability (Avramidou et al., 2023) and is a threat to native ecosystems (Nagy et al., 2018).
Riparian vegetation is strongly influenced by hydrological factors (Dijk et al., 2013; James et al., 2016; Zhu et al., 2017) along river systems. Variation in flow patterns and water depth influences seed distribution, which can affect vegetation stability (Dijk et al., 2013; Zhu et al., 2017). Also, topographic factors and landscape structure control the hydrological response, which will subsequently shape the spatial distribution and diversity of riparian vegetation (Jencso & McGlynn, 2011; Hwang et al., 2012).
In the study, the data obtained showed a significant seasonal fluctuation of the water level, with minimum values in summer and spring, and maximum values in autumn and winter. These values are comparable with the data in the literature (Romanescu, 2014; Trifanov et al., 2019), which show that precipitation and anthropogenic processes influence the water levels in the Danube Delta. In this context, the variations recorded in this study confirm the characteristic hydrological dynamics of the Delta, being highlighted by seasonal water accumulations. Regarding the water velocity, the data from this study showed an increase in the water velocity in autumn and winter. These results are consistent with the observations in the literature (Pekárová et al., 2019; Crețescu et al., 2021) which show that snowmelt and intense precipitation upstream contribute to the variation of natural factors. Moreover, constant and moderate velocities in the other seasons indicate hydrological stability, in which the redistribution of the flow between the Chilia, Sulina, and Sf. Gheorghe arms influences the water flow regime (Pekárová et al., 2019). In the case of water flow, a significant increase was observed in winter.
According to the literature (Pekárová et al., 2019; Crețescu et al., 2021), the Danube acts as a major water collector, in which the discharges are influenced by climatic conditions and flows from upstream countries. At the same time, our study indicates a significant correlation between flow and water velocity during periods of maximum water accumulation. The data obtained for riverbed profiles showed the stability of the riverbed, with minimal variations in depth. According to the literature (Romanescu, 2014; Bănescu et al., 2020), the stability of the riverbed profile is the result of geomorphological processes. In this context, our results confirm that the riverbed is stable and characterized by reduced erosion processes.
The observed morphogenetic processes are closely linked to the Danube’s hydrological regime (Leopold & Maddock, 1953; Bondar, 1992). The intensified dynamics of these sediments are evidenced by considerable changes in shape and area, reflecting their sensitivity to hydrological fluctuations (Knighton, 1998; Gordon et al., 2004). With this sediment accumulation, the rapid colonization of emergent vegetation plays a vital role in soil stabilization, contributing to sediment fixation and the consolidation of island morphology (Bondar, 1992). The dual dynamics, characterized by upstream erosion, reflect a dynamic and unstable morphological equilibrium, influenced by local variations in current and sediment distribution patterns (Panin, 1996). The direction of island A4 expansion is controlled by the complex interaction between water level, velocity, and current direction (Knighton, 1998).
The morphodynamic analysis of the Danube Delta’s islands show the important role of the hydrological regime in shaping this complex ecosystem, highlighting the river’s capacity to induce rapid transformations through sedimentation and erosion processes. The comparative study, utilizing geomorphological methods and remote sensing, has indicated the complex interactions among hydrological, sedimentary, and vegetative factors, offering a detailed perspective on the riverbed including island’s morphological evolution. The data obtained through morphometric analysis signals the necessity for sustainable management, which takes into account the river’s natural dynamics and the potential impact of climate change. Finally, the remarkable intensity of morphodynamic processes, manifested by accelerated rates of sedimentation, erosion, and island shape modification, underscores the highly dynamic nature of the Danube River, where even minor hydrological fluctuations can generate significant morphological transformations within a short timeframe (Bondar et al., 1992; Church & Ferguson, 2015).
Seasonal fluctuations of hydrological factors highlight the influence of natural factors and anthropogenic interventions on the hydrological regime. Also, the stability of the riverbed profile indicates a local geomorphological balance.
From a phytocoenological perspective, six associations and one subassociations were identified, included in four classes: Artemisietea vulgaris, Lemnetea, Phragmito-Magnocaricetea, and Salicetea purpureae.
The morphodynamic analysis of the islands in this study highlights the complexity and dynamics of fluvial processes. The comparative study, utilizing geomorphological methods and remote sensing, provides valuable information for understanding the Danube riverbed morphological evolution and for the sustainable management of its complex ecosystem. The morphometric analysis of these seven Danube Delta islands reveals a complex interplay of erosion and accumulation, influenced by factors such as island location, river flow dynamics, and human interventions like meander rectification.