
Invasions of aquatic macrophytes are negative and dangerous phenomena that can cover the water areas of natural and artificial reservoirs. Dense mats of plants cause shallowing, increase the rates of water siltation, block sunlight, reducing primary production, disturbance in the gas exchange of reservoirs, decrease levels of oxygen and increased levels of nitrate, ammonium and phosphorus, that leads to eutrophication, increase mortality rates of fish and macro-invertebrates (Attionu, 1976; EPPO, 2017; May, 2006). In addition, such water invasions lead to economic losses and significantly complicate economic and environmental protection activities in the affected water bodies (Coetzee et al., 2017). At the same time, eliminating the consequences of large-scale invasions of aquatic macrophytes is a rather difficult task and requires large financial and resource costs (Global Invasive Species Programme (GISP), 2001; May, 2006).
An example of such an invasion was the appearance and further spread of Pistia stratiotes L. in 2013 in the reservoirs of the Siverskyi Donets River valley (Kharkiv region, Ukraine). This made it necessary to solve the problem of the negative impact of large-scale phytoinvasion on natural hydroecosystems (Kazarinova, 2016). After the first detection in 2013, this species appeared every year in various reservoirs and rivers of the Siversky Donets basin, in 2014 it was already recorded within the Kharkiv city in the course of the Udy River (49°57’24.8»N 36°09’59.9»E).
Pistia stratiotes L. (Araceae), or water lettuce, is an alien species of pantropical origin, noted in Ukraine since the 80s of the last century (Dubyna et al., 2017). It is a pleistophyte, floats freely on the water surface, has rapid vegetative reproduction by stolons and the ability to form a large number of seeds with high germination rates (Kazarinova, 2016). P. stratiotes is spread by water currents and wind, as well as by birds, which allows vegetative and generative diaspores of plants to overcome significant distances between isolated water bodies (Adebayo et al, 2011). Plants are capable of rapid vegetative reproduction with the help of stolons that develop in the axils of the lower leaves. New individuals are formed at the ends of stolons, which form stolons of the second order. Adult plants are able to form up to 8 inflorescences, which are contained in the axils of the leaves in the center of the rosette (Kazarinova, 2016). According to our research, this species forms 4–8 fruits in September–October with an average seed productivity 7.1 seeds per fruit, which are characterized by high germination. The optimal conditions for seed germination are high water temperature, which is observed most often in anthropogenically modified ecosystems (Kazarinova, 2016). Our field and laboratory studies confirm the viability of seeds that have been frozen and the overwintering of P. stratiotes in natural water bodies (Kazarinova, 2016).
Pontederia crassipes Mart. (Pontederiaceae) or water hyacinth is an alien species of subtropical-tropical South American origin (Amazon River Basin). According to Prokopuk et al. (2021), this species was first registered in Ukraine in 2020 in the reservoirs of Kyiv city. However, Yu.V. Benhus already found Pontederia crassipes in Kharkiv city in 2009 (Benhus & Neko, 2023). Due to occurrence in different types of water bodies, rapid development and wide distribution, P. crassipes and P. stratiotes have been included by various wildlife agencies around the world among the 100 most invasive and harmful exotic species (Sierra-Carmona et al., 2022).
Both species are widely and, unfortunately, uncontrollably used in urban aquaculture, aquarism, which makes cities a source of their penetration into hydroecosystems with subsequent spread and formation of stable local populations in various types of water bodies. This is facilitated by the competitiveness and euritopicity of these plant species (Prokopuk & Zub, 2022).
In this regard, we aimed to assess the distribution of local populations of Pistia stratiotes and Pontederia crassipes in hydroecosystems, to establish the phytocenotic and ecological features of plant communities with the participation of these alien species using the example of Kharkiv city.
Kharkiv city is located in the North-East of Ukraine between the Central Russian Highlands and the Donetsk Lowlands. The city territory is a hilly plain cut by river valleys, streams and ravines. This territory is a relatively deep erosion basin formed by the activity of the Kharkiv, Lopan, Udy and Nemyshlya Rivers, which belong to the Siversky Donets basin. The Kharkiv, Lopan and Nemyshlya Rivers with their tributaries are small, and the Udy River is medium. The hydrological characteristics of these rivers are given in a number of publications (Shvebs & Ihoshyn, 2003; Karpets, 2008). In addition, there are small reservoirs on the territory of the city – Zhuravlivske, Oleksiivske, Lozovenkivske, Novobavarske and a number of ponds (Gamulya & Zvyagintseva, 2010). The rivers divide the city into three parts, which differ little in area, but have a characteristic relief. Most of the city's territory (about 55%) consists of elevated areas (105–192 m above sea level). Low areas (90–105 m) make up 45.5% of the city's territory, mid areas (105–166 m) a similar 48.2%, and higher altitudes (166–192 m) only 4.3% (Zviahintseva et al., 2023)
Field research was conducted during 2014–2023 in various areas of the city, so that the research covered the main water bodies of Kharkiv. First, these are the Kharkiv, Lopan, Udy and Nemyshlya Rivers. In addition, small reservoirs within the city were surveyed – Zhuravlivske, Oleksiivske, Zhovtneve, Lozovenkivske, Novobavarske and a number of ponds.
Standard floristic and phytosociological methods were used during field research. Determination of the phytocoenotic diversity of aquatic vegetation was based on the analysis of 10 vegetation relevés on plots of 15–25 m2 along three rivers within the city area, where populations of alien species were found (Figure 1).
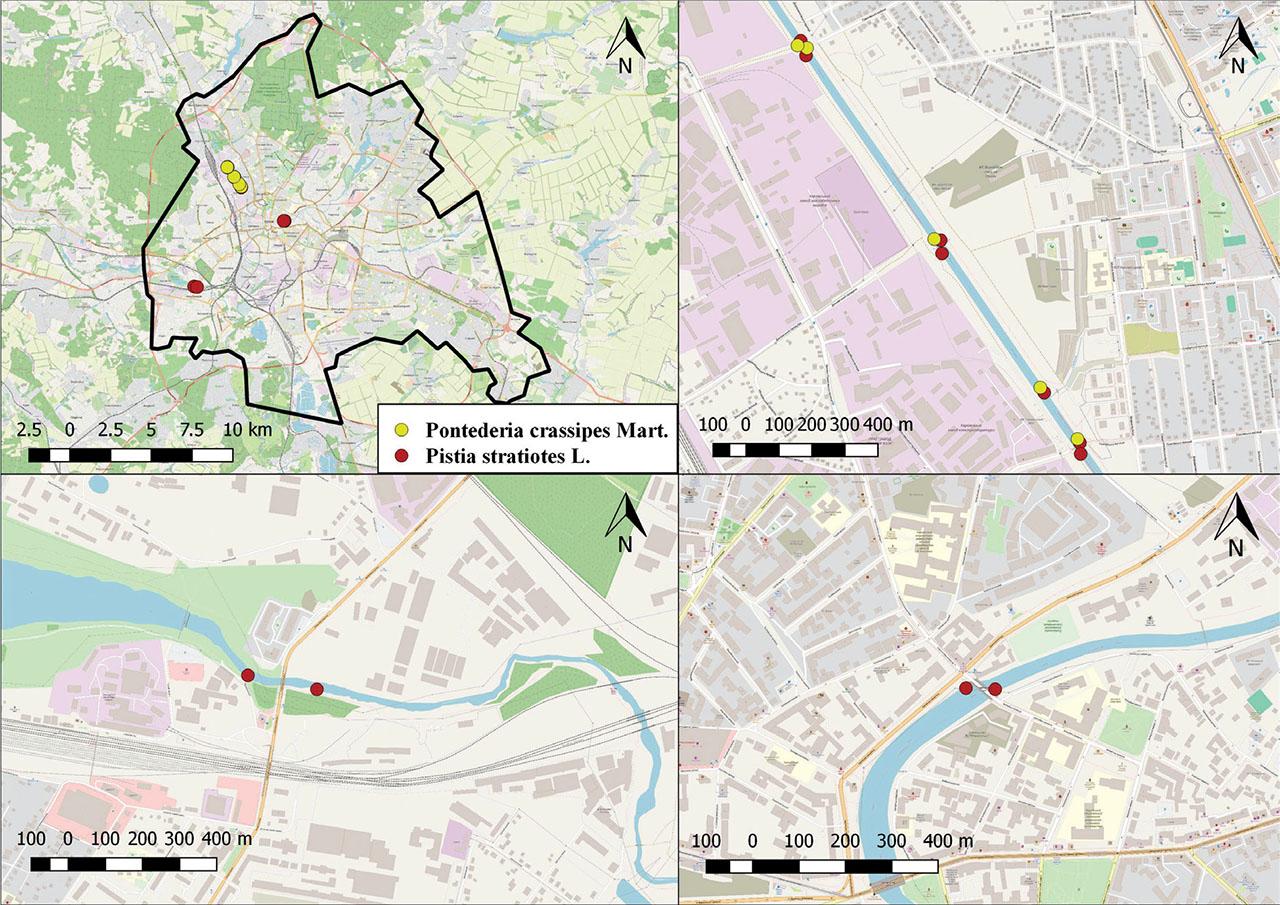
Locations of the investigated plots in Kharkiv city: 1 – Kharkiv city area; 2 – The Lopan River; 3 – The Udy River; 4 – The Kharkiv River.
Slika 1: Lokacije raziskanih ploskev v mestu Harkov: 1 – območje mesta Harkov; 2 – reka Lopan; 3 – reka Udy; 4 – reka Harkov.
We studied the local populations of Pistia stratiotes and Pontederia crassipes in the following habitats (coordinates according to WGS84):
The course of the Udy River below the Zhovtneve Reservoir (three plots on the same bank 49°57′26.9″N 36°09′57.1″E; 49°57′25.1″N 36°09′59.8″E; 49°57′23.2″N 36°10′07.4″E);
The course of the Kharkiv River near the Horbatyi Bridge (two plots on different banks 49°59′32.9″N 36°14′36.6″E; 49°59′33.4″N 36°14′40.0″E);
The course of the Lopan River near Pavlivskyi Bridge (one plot 50°00′40.6″N 36°12′26.6″E);
The course of the Lopan River below the confluence of the right tributary Sarzhynka River (one plot 50°00′47.0″N 36°12′19.9″E);
The course of the Lopan River near the industrial zone (two plots on different banks of the river 50°01′03.5″N 36°12′02.0″E; 50°01′01.8″N 36°12′02.9″E);
The course of the Lopan River near Rudakivskyi Bridge (one plot 50°01′19.1″N 36°11′45.2″E).
Studied locations of water bodies in Kharkiv.
Tabela 2: Preuèevane lokacije vodnih teles v Harkovu.
| Number of location | Type of water body | Date | Water temperature, °C | The presence of alien species | Geographical coordinates | |
|---|---|---|---|---|---|---|
| Longitude | Latitude | |||||
| 1 | Udy river | 19.09.2014 | 24 | Pistia stratiotes (projective cover 25%) | 36°09′59.8″E | 49°57′25.1″N |
| 2 | Udy river | 19.09.2014 | 24 | Pistia stratiotes (projective cover 15%) | 36°10′07.4″E | 49°57′23.2″N |
| 3 | Zhovtneve Reservoir | 19.09.2014 | 23 | Pistia stratiotes (projective cover 25 %) | 36°09′57.1″E | 49°57′26.9″N |
| 4 | Kharkiv river | 20.08.2016 | 22 | Pistia stratiotes (projective cover 40%) | 36°14′36.6″E | 49°59′32.9″N |
| 5 | Kharkiv river | 30.09.2020 | 14 | Pistia stratiotes (projective cover 35%) | 36°14′40.0″E | 49°59′33.4″N |
| 6 | Lopan river | 6.09.2021 | 18 | Pistia stratiotes (projective cover 30%), Pontederia crassipes (projective cover 5%) | 36°12′26.6″E | 50°00′40.6″N |
| 7 | Lopan river | 6.09.2021 | 18 | Pistia stratiotes (projective cover 5%), Pontederia crassipes (projective cover 5%) | 36°12′19.9″E | 50°00′47.0″N |
| 8 | Lopan river | 6.09.2021 | 18 | Pistia stratiotes (projective cover 5%), Pontederia crassipes (projective cover 10%) | 36°12′02.0″E | 50°01′03.5″N |
| 9 | Lopan river | 6.09.2021 | 19 | Pistia stratiotes (projective cover 5%), Pontederia crassipes (projective cover 15%) | 36°12′02.9″E | 50°01′01.8″N |
| 10 | Lopan river | 6.09.2021 | 18 | Pistia stratiotes (projective cover 45%), Pontederia crassipes (projective cover 15%) | 36°11′45.2″E | 50°01′19.1″N |
The studies were carried out from the end of August to the second half of September during 2014–2021. The water temperature in the studied areas ranged from +14–+19 °C in Kharkiv river (2020–2021) to +22–+24 °C in Udy river (2014–2016). Field research in 2022 was not conducted due to the shelling of the city and mine danger. On September 9–10, 2023, areas of the Lopan River and the Kharkiv River were surveyed, both alien species were not recorded.
All stands were recorded according to the method of Braun-Blanquet (Braun-Blanquet, 1964) on sites with a more or less uniform cover of aquatic vegetation. We chose 10 plants at different points of the plot in all sampling locations and measured the diameter of adults and the length of leaves. The water temperature was also measured at all locations. Quantitative participation of species was calculated using the Braun-Blanquet abundance scale, where “r”<less than 1% plot cover, 3–5 individuals; “+” <5%, few individuals; “1”<5%; numerous individuals; “2” 5–25%; “3” 25–50%; “4” 50–75%; “5” 75–100%. Vegetation relevés were stored in Turboveg 2.91 database (Hennekens & Schaminée, 2001). Classification and ecological indicator values analysis was carried out using program Juice 7.0.127 (Tichý, 2002). Further determination of plant communities was carried out according to the Prodromus of the Vegetation of Ukraine (Dubyna et al., 2019). The method of synphytoindication was used to determine the ecological optima of plant communities in relation to the leading environmental factors (Didukh, 2011). For this, 12 quantitative indicators were used, with values of ecological characteristics ascribed to more than three thousand species. Quantitative indicators for plant communities were calculated based on the average indicators of species’ tolerance amplitudes of all informative species recorded in a plot, accounting for their projective cover.
ϒ = k1x1 + k2x2 +…knxn / k1 + k2 +…kn, where x1…n – the mean of species’ tolerance amplitudes in accordance with scale, k1…n – the coefficients of cover, n – number of informative species (Didukh, 2011).
The taxonomy of species is given in accordance with Plants of the World Online (POWO, 2023).
As a result of field research, local populations of alien species were recorded in three out of four rivers surveyed. We did not detect any of the species in the surveyed reservoirs or ponds of the Kharkiv city (Figure 2).

Aquatic plant communities with present alien species Pistia stratiotes L. and Pontederia crassipes Mart.: A) the Udy River (19. 9. 2014); B) the Kharkiv River (20. 8. 2016); C) the Lopan River (6. 9. 2021); D) the Lopan River (6. 9. 2021).
Slika 2: Vodne rastlinske združbe s prisotnima tujerodnima vrstama Pistia stratiotes L. in Pontederia crassipes Mart.: A) reka Udy (19. 9. 2014); B) reka Harkov (20. 8. 2016); C) reka Lopan (6. 9. 2021); D) reka Lopan (9. 6. 2021).
Pistia stratiotes was first registered in Kharkiv in 2014 in the Udy River. Probably, the species got to the river from places of trade in aquarium plants, which were located nearby. In 2016, P. stratiotes appeared in the Kharkiv River, where we continuously monitored the population from 2016 to 2020. The size of adult plants reached 12–15 cm in diameter. Up to 5–6 plants were combined into a single system of shoots through stolons. In the autumn of 2016, large individuals had flowers and seeds at water temperatures from +13°C (October) to +3 °C (November). No seed formation was observed in 2020.
The local population of Pontederia crassipes Mart. was first discovered in August 2021 on the Lopan River in the city center. The plants formed aquatic thickets with a projective cover of 5 to 15% together with a population of P. stratiotes whose projective cover ranged from 5 to 40%. The size of adult individuals of P. stratiotes reached 37–38 cm in diameter, up to 5–6 plants connected by stolons into a system of shoots. Individuals of Pontederia crassipes had diameter of up to 30 cm, the size of individual systems of shoots was up to 50 cm.
Based on the results of classification, a phytocoenotic table of plant communities with the participation of both alien species was constructed and published in the conference materials (Kazarinova, 2023). Both species occurred with high abundance in the communities of Lemnetea O. de Bolòs et Masclans 1955 and with low abundance in the fragments of communities Phragmito-Magnocaricetea Klika in Klika et Novák 1941, which are constantly mown along the rivers of the city. The plant communities were characterized by a small floristic richness (from 6 to 11 species), with a total projective cover of 40 to 100% and occupied shallow areas of three rivers with a depth of 10–50 cm.
The vegetation of the surveyed areas is represented by Hydrocharitetum morsus-ranae van Langendonck 1935 with a projective cover of the diagnostic species Hydrocharis morsus-ranae L. up to 80%, Ceratophylletum demersi Corillion 1957 with a projective cover of Ceratophyllum demersum L. up to 60%, Myriophyllo-Nupharetum Koch 1926 with a projective cover of Nuphar lutea (L.) Sm. up to 50%. During 2022–2023, Phragmitetum australis Savič 1926, Typhetum angustifoliae Pignatti 1953, Sparganietum erecti Roll 1938, Glycerietum maximae Nowiński 1930 corr. Šumberová, Chytrý et Danihelka in Chytrý 2011 spread along the banks of the Kharkiv River and almost half of the Lopan River bed.
According to the results of ecological analyses, plant communities with Pistia stratiotes and Pontederia crassipes show sensitivity to low temperatures, light, total salt regime and nitrogen content. A comparison of ecological indicator values of plant communities indicates an increase in the amplitude to variability of humidity for coenoses with Pontederia crassipes compared to communities with Pistia stratiotes (Figure 3).
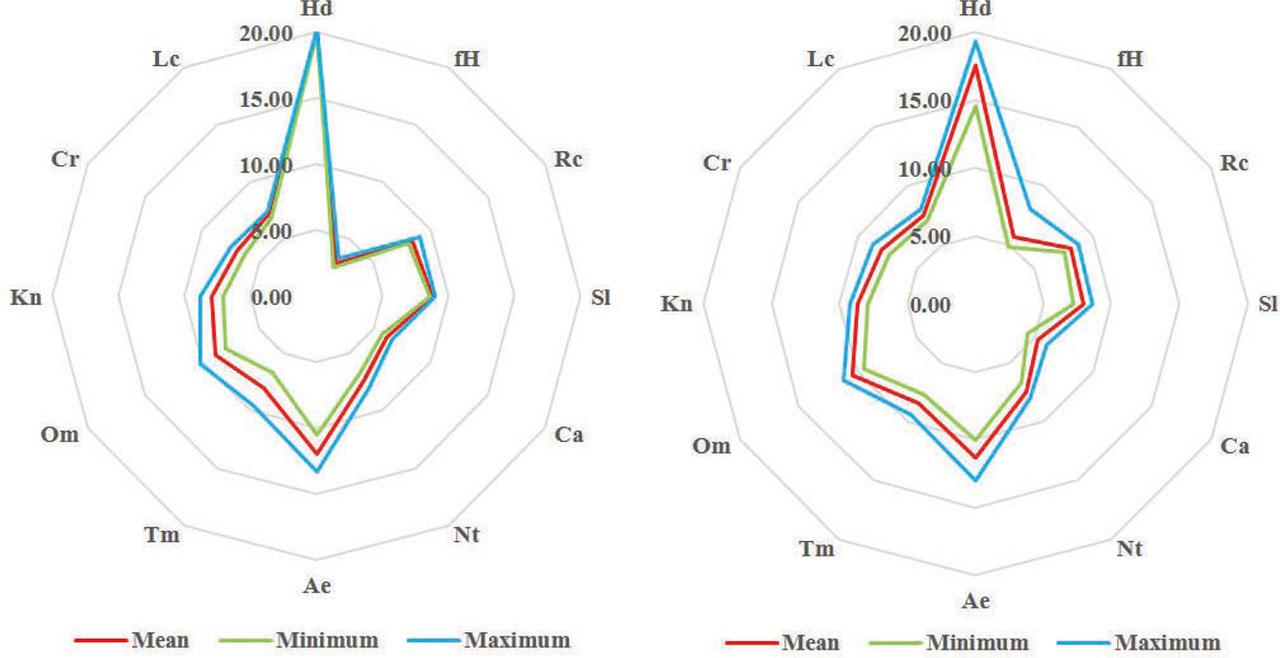
Zone of tolerance of plant communities in the hyperspace of Didukh's ecological indicator values: A) with Pistia stratiotes; B) with Pistia stratiotes and Pontederia crassipes
Legend: Hd – soil humidity, fH – variability of soil humidity, Rc – acidity, Sl – total salt regime, Ca – carbonate content, Nt – nitrogen content, Ae – soil aeration, Tm – the radiation balance (thermal climate), Om – the aridity or humidity of climate (ombroregime), Kn – continentality of climate, Cr – the rigours of winter (cryoclimate), Lc – light.
Slika 3: Cona tolerance rastlinskih združb v hiperprostoru vrednosti Didukhovih ekoloških indikatorskih vrednosti: A) z vrsto Pistia stratiotes; B) z vrstama Pistia stratiotes in Pontederia crassipes
Legenda: Hd – vlažnost tal, fH – variabilnost vlažnosti tal, Rc – kislost, Sl – skupni režim soli, Ca – vsebnost karbonatov, Nt – vsebnost dušika, Ae – zračnost tal, Tm – sevalna bilanca (toplota podnebja), Om – suhost ali vlažnost podnebja (ombrorežim), Kn – celinskost podnebja, Cr – ostrina zime (krioklima), Lc – svetloba.
Phytoindicative analysis based on ecological indicator values for 10 vegetation relevés is illustrated in Table 2. Studied plant communities are formed in water conditions with fluctuation from stable water level (hydrocontrastophobic) to irregular water level (hemihydrocontrastophilic conditions).
Ecological indicator values of studied plant communities according to twelve indicators.
Tabela 2: Vrednosti dvanajstih ekoloških indikatorskih vrednosti raziskovanih rastlinskih združb.
| Environmental factors | Plant communities with participation | Plant communities with participation | ||||||
|---|---|---|---|---|---|---|---|---|
| Pistia stratiotes | Pistia stratiotes and Pontederia crassipes | |||||||
| Mean | Minimum | Maximum | Std. Dev. | Mean | Minimum | Maximum | Std. Dev. | |
| Hd | 20,13 | 20,00 | 20,30 | 0,15 | 17,48 | 14,50 | 19,30 | 1,86 |
| fH | 2,90 | 2,60 | 3,30 | 0,36 | 5,68 | 4,80 | 8,00 | 1,33 |
| Rc | 8,43 | 8,00 | 9,10 | 0,59 | 8,10 | 7,60 | 8,70 | 0,51 |
| Sl | 8,83 | 8,60 | 9,00 | 0,21 | 7,94 | 7,20 | 8,60 | 0,60 |
| Ca | 6,17 | 5,70 | 6,60 | 0,45 | 5,34 | 4,40 | 6,10 | 0,67 |
| Nt | 7,30 | 6,70 | 8,00 | 0,66 | 7,44 | 6,70 | 8,00 | 0,57 |
| Ae | 11,90 | 10,50 | 13,30 | 1,40 | 11,34 | 10,00 | 13,00 | 1,09 |
| Tm | 7,97 | 6,70 | 9,60 | 1,48 | 8,46 | 7,70 | 9,40 | 0,66 |
| Om | 8,83 | 7,90 | 10,20 | 1,21 | 10,48 | 9,50 | 11,20 | 0,90 |
| Kn | 7,90 | 7,10 | 8,80 | 0,85 | 8,64 | 7,90 | 9,20 | 0,47 |
| Cr | 6,97 | 6,30 | 7,50 | 0,61 | 7,92 | 7,30 | 8,70 | 0,61 |
| Lc | 7,20 | 6,90 | 7,40 | 0,26 | 7,54 | 7,10 | 8,00 | 0,38 |
Regarding soil reaction, plant communities are formed in neutrophilic conditions and have a small amplitude. According to the total salt regime, plant communities grow in eutrophic conditions. According to the carbonate content, coenoses exhibit hemicarbonatophobia. According to the nitrogen content, communities show nitrophily. The ecological indicator values of the detected plant communities according to the soil aeration indicates their formation in weakly aerated environment (aerophobic and megaaerophobic conditions).
Plant communities have fairly wide ranges of tolerance to the influence of climatic factors, as well as a shift in the climate humidity towards drier conditions according to conducted phytoindication analysis by this indicator.
According to the radiation balance and humidity of climate, the coenoses are differentiated in the range of values that correspond to submicro-submesothermal (35–50 kcal/cm2) and meso-subaridophyte conditions (humidity index according to Didukh (2011) from −1000 mm to −200 mm). According to the continentality of climate, the amplitude of plant communities covers indicators from hemi-oceanic to hemi-continental climate, which corresponds to 120–140% according to Ivanov formula for the estimation of continental climate (Didukh, 2011) and explained by the location of Ukraine. According to characteristic of winter extreme conditions (the effect of low temperatures), the ecological amplitude of coenoses corresponds to sub-cryophytichemicryophytic conditions with temperature variations of the coldest months from −18°C to −6°C. According to the light regime, plant communities are able to grow in subheliophytic conditions with slight shading.
Results of phytoindicative analysis show forming of studied plant communities in conditions of variability of humidity, minimal soil aeration, its neutral reaction, eutrophication, supply with nitrogen content and insignificant carbonate content. Phytoindication by climatic indicators show wide ranges of tolerance of plant communities under submicro-submesothermic, meso-subaridophytic, subcryophytic-hemicryophytic conditions.
Ecological analysis of studied aquatic and riparian vegetation according to the method of synphytoindication (Didukh, 2011) show the spread of plant communities with alien species in areas characterized by fluctuations in humidity, soil aeration and the temperature regime of water bodies. According to our previous studies (Kazarinova, 2016), such ecological indicators as the effect of low temperatures, light, total salt regime and nitrogen content are limiting for plant communities with the participation of Pistia stratiotes. This is also confirmed by research of coenopopulations of P. stratiotes in water bodies of Kyiv city (Prokopuk & Zub, 2022). The same results were obtained in the study of distribution of Pontederia crassipes in South Africa (Coetzee & Hill, 2012). The leading role of temperature regime of water bodies in the growth and spread of invasive species in natural ecosystems is confirmed by different studies (Kazarinova, 2016; Jaklič et al., 2020). The intensity of growth and distribution of alien species depends both on the viability of the seeds that got into water bodies, and on the presence of favorable conditions for overwintering plants and seed germination during the cold period of the year (Kazarinova, 2016; Šajna et al., 2023). We established that anthropogenically altered ecosystems with shifted microclimatic indicators can serve as a kind of “buffers” during the penetration of invasive species into natural ecosystems.
Modeling of the distribution of alien species in hydro-ecosystems indicates the possibility of their colonization of new territories in the future (Mosyakin & Kazarinova, 2014) with P. crassipes potentially expanding beyond its northern range, with the greatest potential for future expansion in Europe (Kriticos & Brunel, 2016). Since the temperature regime in urban conditions is milder, it promotes the introduction and spread of alien tropical species. The large cities are sources of invasions of alien macrophytes due to the escape of the latter from aqua-culture or the deliberate use of water bodies for the reproduction of decorative macrophytes. Another factor is the presence of industry-related facilities with an open water cycle (Dubyna et al., 2017).
The war had a significant impact on the development of Ukrainian cities, including Kharkiv. Many industrial facilities and objects of critical infrastructure have been destroyed or stopped activities, which will undoubtedly have an impact on the further development and spread of alien species in hydroecosystems. Due to the shutdown of some thermal power station, there is no influence of heated waters on the thermal conditions of water bodies and the spread of tropical macrophytes. The shelling of the oil depot (March 2024) caused about 3,000 tons of oil products to enter the water bodies of the city and region, which led to a significant increase in the level of water pollution. Further research will help establish the consequences of such processes for the biodiversity of water bodies in urban environment.
Research on the introduction of alien macrophytes within the Kyiv city (Prokopuk et al., 2021) show that the aquatic communities in urban hydroecosystems are succeptible to the penetration of alien species due to the weakness of coenotic connections and the small species diversity in aquatic plant communities. This process is facilitated by the eutrophication of water bodies, the reduction of species diversity and changes in the structure of hydrobiocenoses. This is confirmed by research of alien macrophytes conducted in Europe (EPPO, 2017)
According to our own research and based on the data of other researchers (Dubyna et al., 2017; Prokopuk et al., 2021), the spread of tropical alien species in water bodies is ephemeral: the mass development of such communities gradually turns into a lull, which is accompanied by a change in the phytocenotic composition of aquatic coenoses with the gradual formation of alien species as their permanent components.
Alien species are distributed with high abundance in the Lemnetea communities and with a lesser abundance in fragments of the Phragmito-Magnocaricetea communities. The leading indicators that determine ecological difference of the investigated plant communities from typical aquatic phytocenoses are thermal regime of water bodies, light, effect of low temperatures (the rigours of the winter), total salt regime. The communities are formed in conditions of variability of humidity, minimal soil aeration, its neutral reaction, eutrophication, supply with nitrogen and insignificant carbonate content. Phytoindication according to climatic indicators showed wide ranges of tolerance of plant communities under the value of radiation balance 30–50 kcal/cm2, the humidity index value from −800 to −200 mm, temperature variation of the coldest months from −14°C to −2°C. The ecological values of plant communities show their adaptation to the climatic and hydrological conditions of the urban habitats, namely increased indicators of the thermal regime, significant content of organic and mineral substances in water bodies. We think that the ability of alien tropical macrophytes to reproduce and overwinter outside of thermally abnormal waters will increase as a result of rising average annual temperatures and lengthening of the growing season in temperate continental climates. A high level of nutrients in the water will favor the development of these species. We can not exclude the possibility of the appearance of other alien species through human introduction (movement of population, goods, transport, the release of aquarium species into natural water bodies).