
Chasmophytes are plant species that grow on rocky walls, often characterised by specific adaptations to thrive in extreme environments with limited soil and unique climatic conditions. Moreover, the rocks create conservative habitats that promote the evolution and survival of paleoendemics (Snogerup, 1971; Antonsson, 2012). Therefore, it is not surprising that a significant percentage of Mediterranean endemism is concentrated in chasmophytic vegetation, particularly those in proximity to the sea (Thompson, 2020). Iconic examples of this chasmophytic flora include Primula palinuri Petagna, Eokochia saxicola (Guss.) Freitag et G.Kadereit, and various species of the Brassica and Centaurea genera (Pignatti et al., 2017). This notable phytogeographic interest has sparked numerous phytosociological studies (Brullo & Marcenò, 1997; Biondi et al., 2000; Corbetta et al., 2000; Terzi & D'Amico, 2008; Wagensommer, 2017; Di Pietro & Wagensommer, 2008), recently synthesised for the central Mediterranean by Terzi et al. (2018). This review has highlighted the existence of several geographically localised alliances attributable to three orders: Asplenietalia glandulosi for the Tyrrhenian coasts, including Liguria, Sicily, and Sardinia; Centaureo-Campanuletalia (including Moltkietalia petreae) along the Adriatic coasts; and Onosmetalia frutescentis in the Eastern Mediterranean.
In Italy, one of the most significant centre of endemic chasmophytes is in the Sorrento Peninsula, western Campania, where especially in the short mountain ranges of Monti Lattari and Monti Picentini, numerous taxa with a very restricted range, some punctiform and generally rare, can be found. Examples include Lonicera stabiana, Globularia cordifolia subsp. neapolitana, Santolina neapolitana, Galium lucidum subsp. venustum, Centaurea tenorei, and Seseli polyphyllum. Although these species were extensively studied by different botanists (Tenore, 1823; Pasquale, 1875; Lacaita, 1922; Schwarz, 1938; Moraldo et al., 1985; Natali, 1998; Del Guacchio et al., 2020), they are often subjects of taxonomic controversies, partially arising from a limited knowledge of the local populations of these species. Overall, the Sorrento Peninsula remains relatively understudied from a vegetation and ecological standpoint, with some notable exceptions (Guadagno, 1916; Salerno et al., 2007; Fanelli et al., 2020).
In the Sorrento Peninsula, two zones of chasmophytic vegetation can be distinguished: one closer to the sea, where intense anthropogenic impacts have made their presence sporadic, and the other in the mountainous zone, housing the majority of endemics. The focus of this study is the latter vegetation zone.
This study expands on a previous investigation that identified an association dominated by Lonicera stabiana, an exceedingly rare stenoendemic species (Salerno et al., 2007). Although the previous work was focused on the description of the associations (Fanelli et al., 2020), it allowed for some initial speculations on the mountainous chasmophytic vegetation of the Sorrento region. This follow-up study aims to 1) review the chasmophytic associations of the Sorrento Peninsula's mountainous area, particularly the rare association Globulario neapolitanae-Loniceretum stabianae, and 2) to contextualise these associations at the level of order and alliance within the chasmophytic vegetation of the central Mediterranean.
The investigated area encompasses two mountainous ranges, one in the Gulf of Naples and the second one in the inland area between Salerno and Avellino provinces (Figure 1). The Gulf of Naples extends over 195 kilometres, spanning from Mount Procida to “Punta Campanella”. Towards the south, there is the Sorrento Peninsula, a modest mountain range jutting out into the sea, which reaches its highest point at Mount Sant'Angelo a Tre Pizzi (1444 m) and only one more notable peak exceeding 1000 meters, i.e. Mount Faito (1131 m).
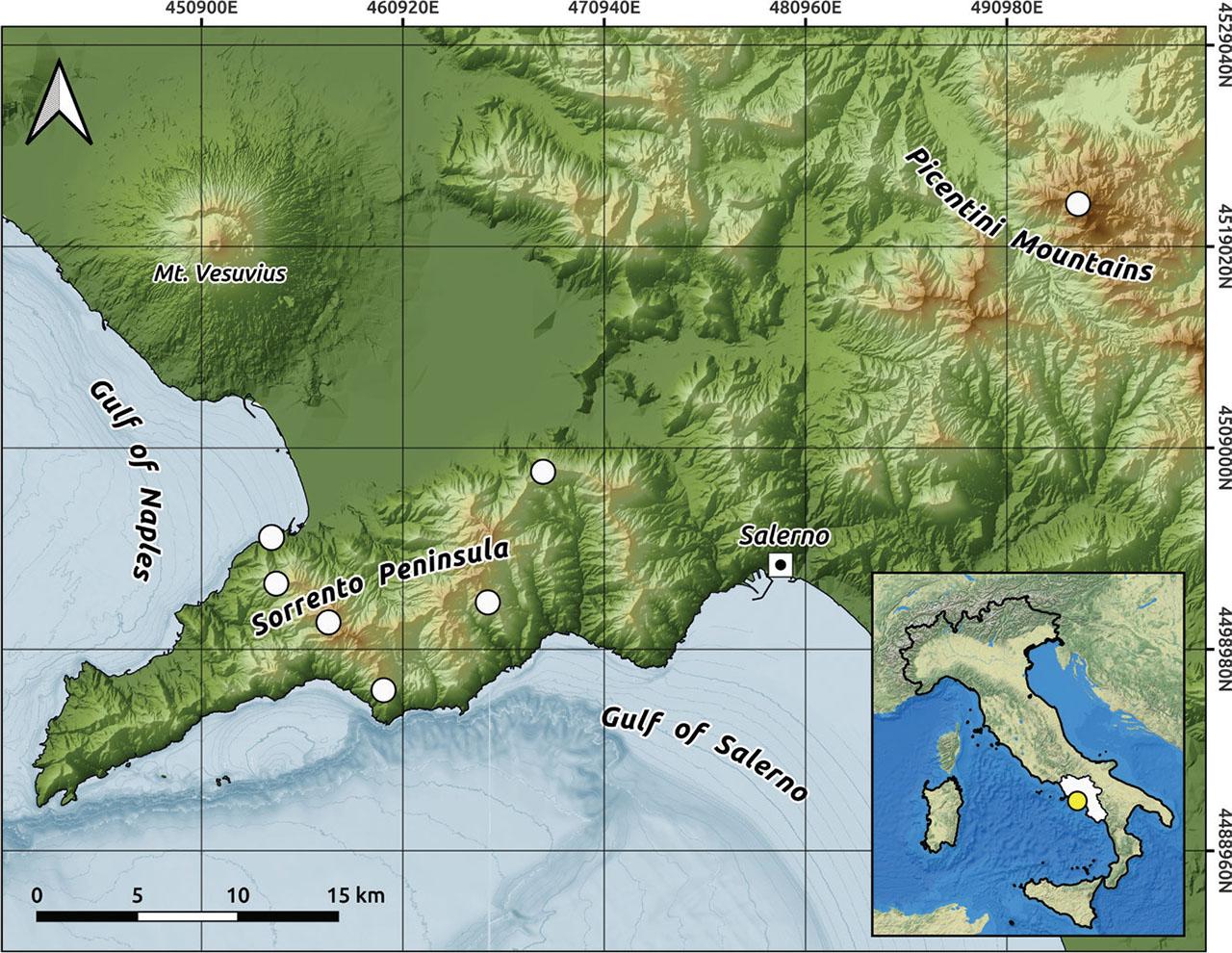
Map of the study area. White points indicate the places where the relevés have been conducted.
Slika 1: Zemljevid preučevanega območja. Bele točke predstavljajo lokacije vegetacijskih popisov.
The geology of this mountainous region is dominated by dolomitic steep slopes and cliffs. The geological complexity of the area results from the Apennine orogenesis, followed by an extensional phase that led to the formation of volcanoes along the Tyrrhenian coast. These geological phases are evident in the Sorrento Peninsula, marked by an anti-Apennine thrust, and the volcanic complex of Vesuvius and Campi Flegrei (Casciello et al., 2006).
The Picentini Mountains, located eastward from the Sorrento Peninsula, exhibit a diverse geological and stratigraphic composition. Primarily consisting of folded and faulted sedimentary rocks like limestones and marls, these mountains reflect their origin in a marine environment. The Terminio-Accellica unit is of particular interest as presenting dolomitic limestones and one of the highest peaks of the area (Terminio Mountain, 1806 m a.s.l.; Pappone & Ferranti, 1995). Volcanic activities from the Campanian Volcanic Arc has also left traces of tuffs and lavas, adding one more layer to the region's geological structure (Pescatore et al., 2022).
The whole area can be referred to the Mediterranean macrobioclimate, except for the areas at higher altitudes, such as Mt. Faito and Mt. San Michele in the Sorrento Peninsula, and the whole complex of the Picentini mountains, featured by a temperate thermotype and an upper humid ombrotype climate. The continentality index shows these altitudes as weak semicontinental (Pesaresi et al., 2017).
The Picentini Mountains and Sorrento Peninsula have experienced significant anthropogenic impacts stemming from centuries of deforestation, agricultural practices and other human activities that have remarkably shaped the landscape and increasingly exposed the natural and semi-natural ecosystems to soil erosion and degradation in local ecosystems. In more recent times, urbanisation, unregulated tourism and infrastructure development have exacerbated habitat fragmentation and loss of biodiversity (Cancellieri & Caneva, 2007; Assennato et al., 2022).
Over two field campaigns in the spring and autumn of 2022, twenty phytosociological relevés were carried out in the Lattari Mountains area, surveying both the Sorrento and Amalfi coasts, as well as the peak of Mount Terminio in the Picentini Mountains. Additionally, we added eight relevés from the previous work by Fanelli et al. (2020). The relevés were carried out following the Braun-Blanquet method (Westhoff & van der Maarel, 1980) using the modified Braun-Blanquet scale of abundance (Barkman et al. 1964). The taxonomical nomenclature follows Bartolucci et al. (2018).
The area of relevés is on average from 1 to 5 m2, occasionally with much larger relevés to 50 m2. Since this could represent potentially a bias, we have carried out a supplementary analysis excluding relevés with area larger than 10 m2.
Different types of multivariate analysis methods were employed to process the vegetation data. An ordination analysis through cluster analysis was performed to categorise the relevés into distinct groups. Euclidean distance was used as a dissimilarity measure, and the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) method was employed for clustering. Before selecting the Euclidean distance, we conducted several preliminary trials, finding this measure to be the most effective for distinguishing clusters. The silhouette test and cophenetic distance were used respectively to estimate the optimal number of clusters and as measures of the goodness of the cluster analysis (Kassambara, 2017).
After obtaining the cluster division of the relevés, we proceeded with an Indicator Species Analysis (ISA) (De Cáceres et al., 2010), which revealed the association between species patterns and the selected group combinations. The number of permutations used was 4999.
A Non-Metric Multidimensional Scaling (NMDS) was carried out to visualise the identified groups in a two-dimensional space. Two field-collected environmental variables (Elevation and Slope) and five bioclimatic variables calculated for the Mediterranean region based on Rivas-Martínez et al. (2011) (unpublished data) were overlaid to potentially explain the ordination of the relevés (Table 1). The same distance measure used for cluster analysis was applied to NMDS, with 1000 random starts and a seed set to “123” for analysis repeatability. The “envfit” function with 999 permutations was employed for this purpose. To distinguish the classes identified through multivariate analysis and to provide a statistical rather than merely descriptive value, we conducted Kruskal-Wallis tests between the groups and for each variable, represented using boxplots. All the analyses in this paper were computed with R software (R Core Team, 2021).
Environmental and bioclimatic variables used for the ordination analyses and to differentiate the different groups.
Tabela 1: Okoljske in bioklimatske spremenljivke, uporabljene v ordinacijski analizi in za ločevanje različnih skupin.
| Variable | Description |
|---|---|
| Elevation | Vertical distance from the sea level |
| Slope | Vertical change in the elevation determined over a given horizontal distance, expressed in degrees |
| Tp | Annual Positive Temperature – sum of monthly average temperatures in months where higher than 0 °C |
| Ic | Continentality index |
| Itc | Compensated thermal index |
| Io | Umbrothermic index |
| TpIc | Annual Positive Temperature and Continentality Index ratio |
Finally, we compared our findings with the chasmophytic vegetation of the Mediterranean through a synoptic table, incorporating our relevés with others from the literature and highlighting the different syntaxa. We included vegetation from the Italian Peninsula and Sicily, as well as some associations from Croatia (Trinajstić, 1987; Brullo & Marcenò, 1997; Biondi et al., 2000; Corbetta et al., 2000; Di Pietro & Wagensommer, 2008; Terzi & D'Amico, 2008). The vegetation from the Eastern Mediterranean, Provence, and the Sardo-Corsican region, floristically distinct from the Sorrento Peninsula, was not considered. The orders Asplenietalia glandulosi, Centaureo-Campanuletalia, and Onosmetalia frutescentis were included. We also included the order Potentilletalia caulescentis due to the relatively high altitudes of our relevés. In structuring the table we followed Terzi et al. (2018) and Biondi et al. (2000), the latter only for Potentilletalia caulescentis.
To further characterise the syntaxa retrieved, we overlapped our relevés on the maps of bioclimatic indexes according to Rivas-Martínez et al. (2011) calculated by Pesaresi et al. (2017) for the entire Italian territory to compare with our ordination results. We considered ombrotype, thermotype, continentality and bioclimate variants.
The silhouette test (Suppl. material 1) identified three well-differentiated clusters in the dendrogram (Figure 2 and Table 2). Furthermore, a correlation test between the original distance matrix and the cophenetic distance of the dendrogram, with a value of 0.803, indicates the validity of the ordering (Suppl. material 1). To exclude potential biases to the uneven area of the relevés, a parallel analysis on relevés not larger than 10 m2 returned exactly the same three clusters (Suppl. Material 1).
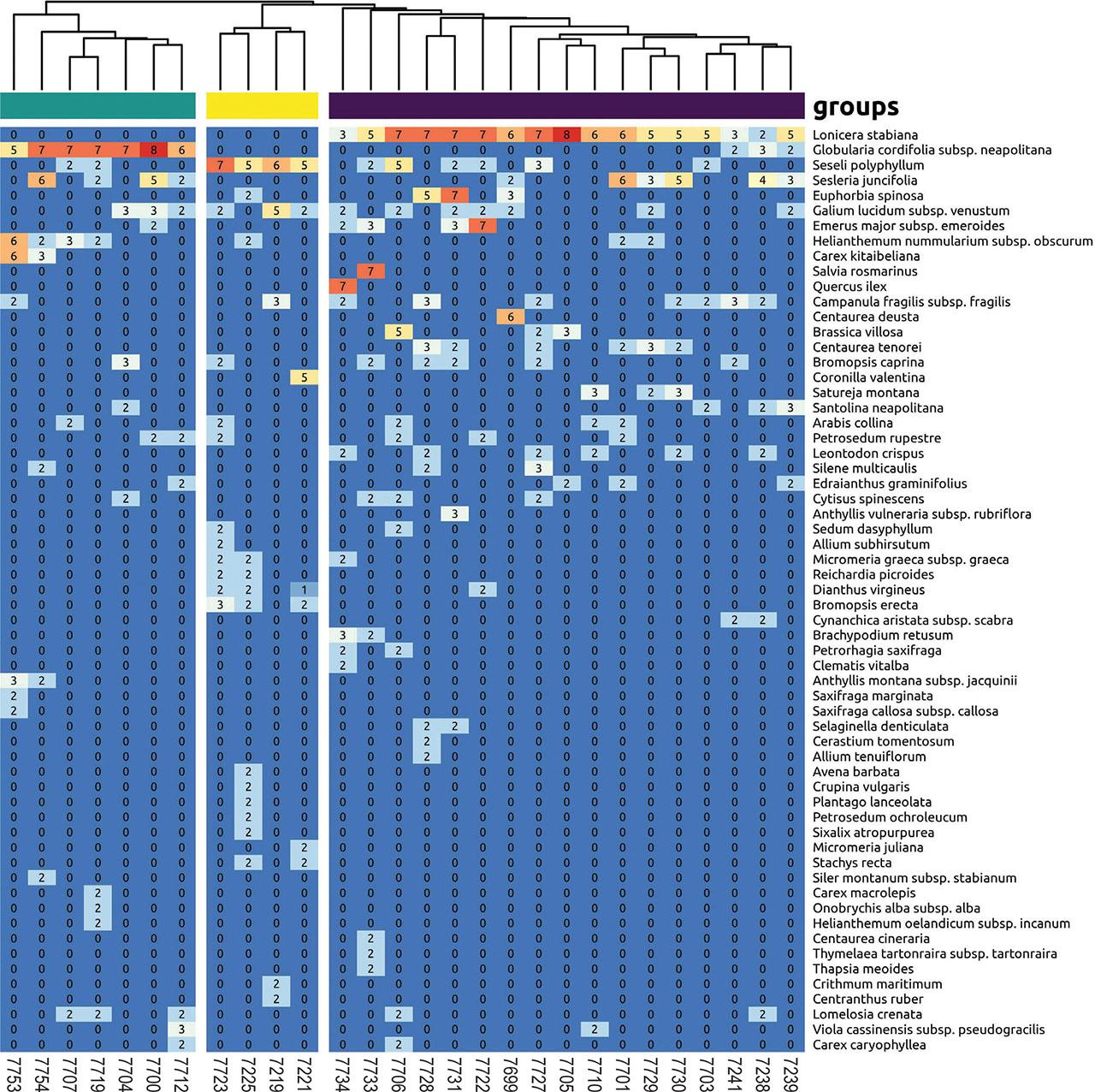
Dendrogram retrieved from the cluster analysis together with species abundances. The three groups are clearly separated by different indicator species.
Slika 2: Dendrogram klastrske analize z abundancami vrst. Tri skupine so jasno ločene z različnimi indikatrorskimi vrstami.
The relevé clusters (1, 2, 3), subjected to the analysis of indicator species, correspond with the dominance of Lonicera stabiana, Globularia cordifolia subsp. neapolitana and Seseli polyphyllum, for cluster 1, 2 and 3, respectively. For the ISA, Lonicera stabiana and Globularia cordifolia subsp. neapolitana represent the unique indicator species for clusters 1 and 2, while cluster 3 identifies several differential species (Suppl. material 1). NMDS clearly distinguishes the 3 clusters (Figure 3), with a stress value of 0.15.
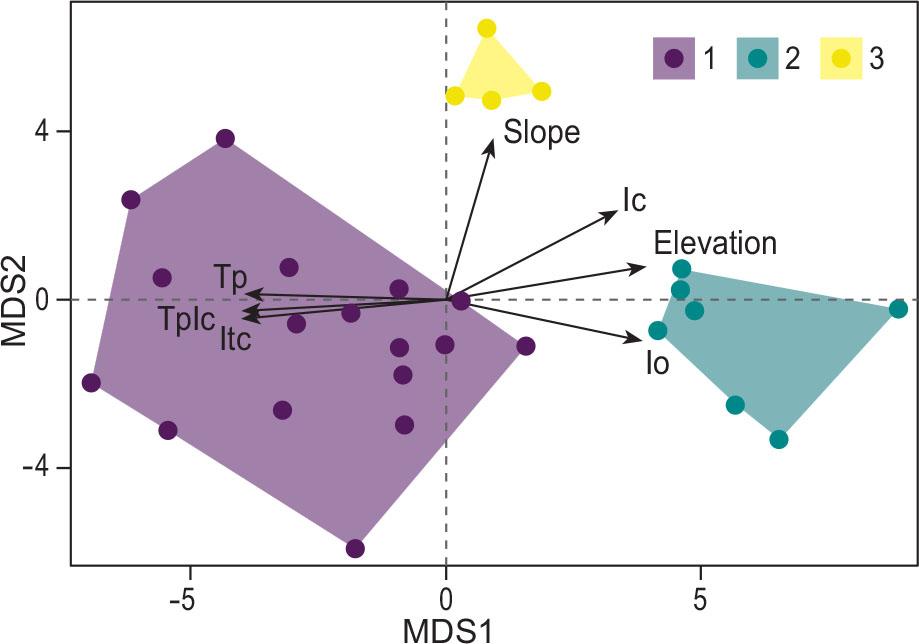
NMDS ordination analysis depicting separation of the three different groups and clearly separated with the environmental and bioclimatic variables. Ic=Continentality index; Io=Ombrothermic index; Itc=Compensated thermal index; Tp=Annual Positive Temperature; TpIc=Tp & Ic ratio.
Slika 3: NMDS ordinacija prikazuje ločitev treh različnih skupin, ki so jasno ločene z okoljskimi in bioklimatskimi spremenljivkami. Ic=indeks kontinentalnosti; Io=ombrotermični indeks; Itc=kompenzirani termični indeks; Tp=letne pozitivne temperature; TpIc=razmerje Tp in Ic.
The biplot of the environmental and bioclimatic variables clearly distinguishes the three clusters. On the first axis, cluster 1 (Lonicera stabiana dominant) and cluster 2 (Globularia cordifolia subsp. neapolitana dominant) are separated according to Io, Itc, Tp and TpIc. Cluster 2 and 3 are separated from cluster 1 on the positive side of Ic index. Among all these variables, Tp, Itc, Io, and TpIc are significant according to the envfit output. The range of the variables in the three different clusters, with the corresponding Kruskal-Wallis test result, is presented in the boxplots of Figure 4. This figure shows in detail the main differences between identified clusters. In particular cluster 1 (Globulario-Loniceretum typicum) and cluster 2 (Globulario-Loniceretum globularietosum neapolitanae) are clearly and significantly separated from the cluster 3 (Diantho-Seselietum), with the latter found mainly in areas at low elevation, with higher temperatures and lower precipitations than the other two clusters.
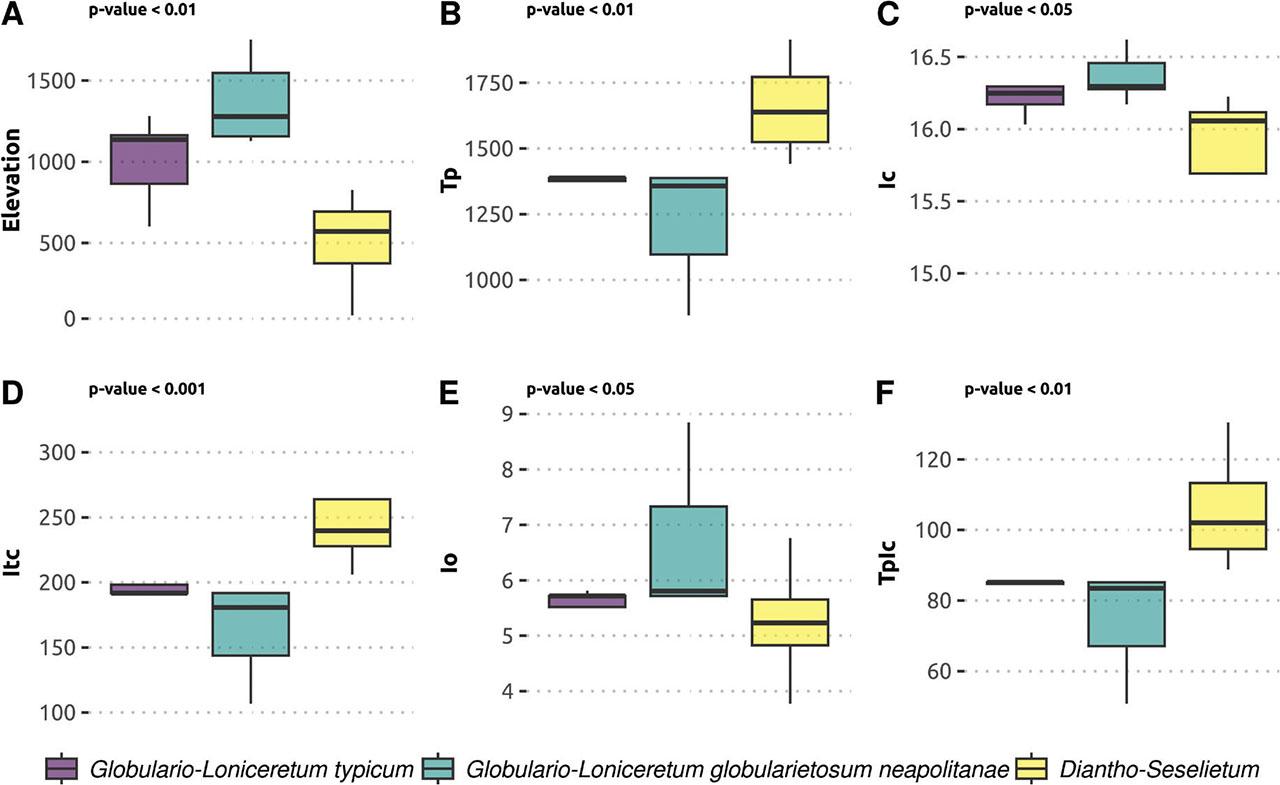
Boxplots depicting the significative differences between the three syntaxa based on the environmental and bioclimatic variables. On top left there are the p-values. A Elevation; B Annual Positive Temperature; C Continentality index; D Compensated thermal index; E Ombrothermic index; F Tp and Ic ratio.
Slika 4: Škatle z okvirji prikazujejo statistično značilne razlike med tremi sintaksoni na osnovi okoljskih in bioklimatskih spremenljivk. Levo zgoraj so prikazane p-vrednosti. A Nadmorska višina; B letne pozitivne temperature; C indeks kontinentalnosti; D kompenzirani termični indeks; E ombrotermični indeks; F TpIc=razmerje Tp in Ic.
According to the synoptic table (Suppl. material 2), a group of species distinguishes the three clusters of vegetation in the Sorrento Peninsula from all others, including Bromopsis caprina, Galium lucidum subsp. venustum, Centaurea tenorei and Santolina neapolitana, while the species of the Tyrrhenian alliances Dianthion rupicolae and Campanulenion fragilis are completely absent. Apart from these species, there is a number of differential species belonging for instance, to Sedo-Scleranthetea, etc. but we did not analyse such species because in the table derived from literature are considered only true character species and not differential. At the order level, the situation is not entirely clear, but it is evident that the species of Asplenietalia glandulosi are completely missing, and there is instead a small number of species of Centaureo-Campanuletalia (Cytisus spinescens, Sesleria juncifolia).
Phytosociological table of the relevés.
Tabela 2: Fitocenološka tabela vegetacijskih popisov.
| Relevé n. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | *17 | 18 | 19 | 20 | 21 | 22 | 23 | *24 | 25 | 26 | 27 | *28 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Relevé area (m2) | 5,0 | 1,0 | 8,0 | 3,0 | 3,0 | 1,0 | 4,0 | 2,5 | 1,5 | 2,0 | 2,5 | 1,0 | 1,0 | 1,0 | 6,0 | 20,0 | 15,0 | 2,0 | 3,0 | 1,0 | 2,0 | 1,0 | 0,5 | 0,5 | 4,0 | 5,0 | 50,0 | 10,0 | |
| Altitude (m a.s.l.) | 1163 | 1132 | 1153 | 1166 | 1170 | 1281 | 651 | 861 | 861 | 861 | 889 | 877 | 602 | 610 | 642 | 1160 | 1157 | 1163 | 1153 | 1170 | 1278 | 1342 | 1752 | 1750 | 649 | 50 | 668 | 826 | |
| Exposure | W-SW | SW | SE | E-SE | SW | W | W-SW | E | E | E | E | NE | SE | N | W | SW | SW | W-SW | SE | SW | W | W | SW | W-SW | W-SW | W | N | SE | |
| Slope (°) | 90 | 80 | 90 | 90 | 90 | 90 | 80 | 85 | 90 | 90 | 90 | 90 | 80 | 85 | 90 | 90 | 90 | 80 | 90 | 90 | 80 | 60 | 90 | 70 | 70 | 90 | 90 | 90 | |
| Cover herb layer (%) | 50 | 70 | 40 | 90 | 80 | 40 | 80 | 60 | 50 | 35 | 70 | 70 | 60 | 30 | 15 | 35 | 10 | 80 | 60 | 30 | 20 | 80 | 50 | 80 | 50 | 30 | 20 | 35 | |
| Char. Globulario neapolitanae-Loniceretum stabianae typicum Fanelli et al. 2020 | Freq. | ||||||||||||||||||||||||||||
| Lonicera stabiana | 2B | 2B | 2A | 4 | 3 | 2B | 3 | 3 | 3 | 2A | 2A | 3 | 2A | 1 | + | 2A | 1 | · | · | · | · | · | · | · | · | · | · | · | 61 |
| Char. Globulario neapolitanae-Loniceretum stabianae globularietosum neapolitanae subass. nova | |||||||||||||||||||||||||||||
| Globularia cordifolia subsp. neapolitana | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 1 | + | + | 4 | 3 | 3 | 2B | 3 | 2A | 3 | · | · | · | · | 36 |
| Char. Diantho virginei-Seselietum polyphylli ass. nova | |||||||||||||||||||||||||||||
| Seseli polyphyllum | · | · | + | · | 2A | · | + | 1 | · | · | · | + | + | · | · | · | · | · | · | + | · | + | · | · | 3 | 2B | 2A | 2A | 43 |
| Dianthus virgineus | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | r | + | 14 |
| Bromopsis erecta | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 1 | · | + | + | 11 |
| Stachys recta | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | + | 7 |
| Micromeria graeca subsp. graeca | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | + | · | · | + | 11 |
| Char. Lonicerion stabianae all. nova | |||||||||||||||||||||||||||||
| Bromopsis caprina | · | · | · | · | · | · | · | + | + | · | · | + | + | · | · | · | + | · | 1 | · | · | · | · | · | + | · | · | · | 25 |
| Galium lucidum subsp. venustum | + | · | · | · | + | · | + | · | · | + | · | + | · | + | · | + | · | 1 | 1 | · | + | · | · | · | + | 2A | + | · | 46 |
| Centaurea tenorei | · | + | · | · | · | · | · | + | 1 | 1 | + | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 21 |
| Santolina neapolitana | · | · | + | · | · | · | · | · | · | · | · | · | · | · | + | 1 | · | · | + | · | · | · | · | · | · | · | · | · | 14 |
| Centaureo-Campanuletalia Trinajstiæ 1980 | |||||||||||||||||||||||||||||
| Cytisus spinescens | · | · | · | · | + | · | · | + | · | · | · | · | + | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | 14 |
| Sesleria juncifolia | + | 2B | · | · | · | · | · | · | · | 1 | 2A | · | · | · | 2M | 1 | · | 2A | · | · | + | + | · | 2B | · | · | · | · | 36 |
| Asplenietea trichomanis Br.-Bl. in Meier et Br.-Bl. 1934) Oberd. 1977 | |||||||||||||||||||||||||||||
| Sedum dasyphyllum | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | 7 |
| Arabis collina | · | + | · | · | + | + | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | + | · | · | · | 18 |
| Euphorbia spinosa | 1 | · | · | · | · | · | · | · | 2A | · | · | 3 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 14 |
| Petrosedum ochroleucum | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 4 |
| Centranthus ruber | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | 4 |
| Companions | |||||||||||||||||||||||||||||
| Campanula fragilis subsp. fragilis | · | · | + | · | · | · | · | + | 1 | · | + | · | · | + | + | · | 1 | · | · | · | · | · | + | · | · | 1 | · | · | 32 |
| Helianthemum nummularium subsp. obscurum | · | + | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | 1 | · | + | 2B | + | · | · | · | + | 25 |
| Leontodon crispus | · | · | · | · | · | + | · | + | + | · | + | · | · | + | + | · | · | · | · | · | · | · | · | · | · | · | · | · | 21 |
| Petrosedum rupestre | · | + | · | · | + | · | + | · | · | · | · | · | · | · | · | · | · | + | · | · | + | · | · | · | + | · | · | · | 21 |
| Emerus major subsp. emeroides | · | · | · | · | · | · | 3 | · | · | · | · | 1 | 1 | + | · | · | · | + | · | · | · | · | · | · | · | · | · | · | 18 |
| Lomelosia crenata | · | · | · | · | + | · | · | · | · | · | · | · | · | · | + | · | · | · | · | + | + | + | · | · | · | · | · | · | 18 |
| Edraianthus graminifolius | · | + | · | + | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | + | · | · | · | · | · | · | · | 14 |
| Satureja montana | · | · | · | · | · | 1 | · | · | · | + | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 11 |
| Brassica villosa | · | · | · | 1 | 2A | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 11 |
| Silene multicaulis | · | · | · | · | · | · | · | 1 | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | 11 |
| Cynanchica aristata subsp. scabra | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | + | · | · | · | · | · | · | · | · | · | · | · | 7 |
| Selaginella denticulata | · | · | · | · | · | · | · | · | + | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 7 |
| Petrorhagia saxifraga | · | · | · | · | + | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 7 |
| Viola cassinensis subsp. pseudogracilis | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 1 | · | · | · | · | · | · | · | 7 |
| Brachypodium retusum | · | · | · | · | · | · | · | · | · | · | · | · | + | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 7 |
| Carex caryophyllea | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | 7 |
| Anthyllis montana subsp. jacquinii | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 1 | + | · | · | · | · | 7 |
| Carex kitaibeliana | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 2B | 1 | · | · | · | · | 7 |
| Reichardia picroides | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | + | 7 |
| Centaurea deusta | 2B | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Anthyllis vulneraria subsp. rubriflora | · | · | · | · | · | · | · | · | · | · | · | 1 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Cerastium tomentosum | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Allium tenuiflorum | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Thymelaea tartonraira | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Salvia rosmarinus | · | · | · | · | · | · | · | · | · | · | · | · | 3 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Thapsia meoides | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Centaurea cineraria | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Quercus ilex | · | · | · | · | · | · | · | · | · | · | · | · | · | 3 | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Clematis vitalba | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 4 |
| Micromeria juliana | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | 4 |
| Coronilla valentina | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | 2A | · | 4 |
| Crithmum maritimum | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | 4 |
| Sixalix atropurpurea | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 4 |
| Plantago lanceolata | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 4 |
| Crupina vulgaris | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 4 |
| Avena barbata | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | 4 |
| Allium subhirsutum | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | 4 |
| Onobrychis alba subsp. alba | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | 4 |
| Helianthemum oelandicum subsp. incanum | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | 4 |
| Carex macrolepis | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | · | 4 |
| Siler montanum subsp. garganicum | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | 4 |
| Saxifraga marginata | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | 4 |
| Saxifraga callosa subsp. callosa | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | · | + | · | · | · | · | · | 4 |
The cluster containing Seseli polyphyllum occupies a lower humid to upper subhumid ombrotypes, predominantly Mediterranean, occasionally temperate thermotypes, and exhibits a weakly euoceanic continentality type (Figure 4). The other two clusters share mainly temperate, occasionally Mediterranean thermotypes, and display a weakly semicontinental, sometimes weak euoceanic conditions (Figure 5 and Figure 6). The clusters differ in that the one dominated by Globularia cordifolia subsp. neapolitana ranges from lower hyperhumid to lower humid, while the cluster dominated by Lonicera stabiana is from upper humid to lower humid.
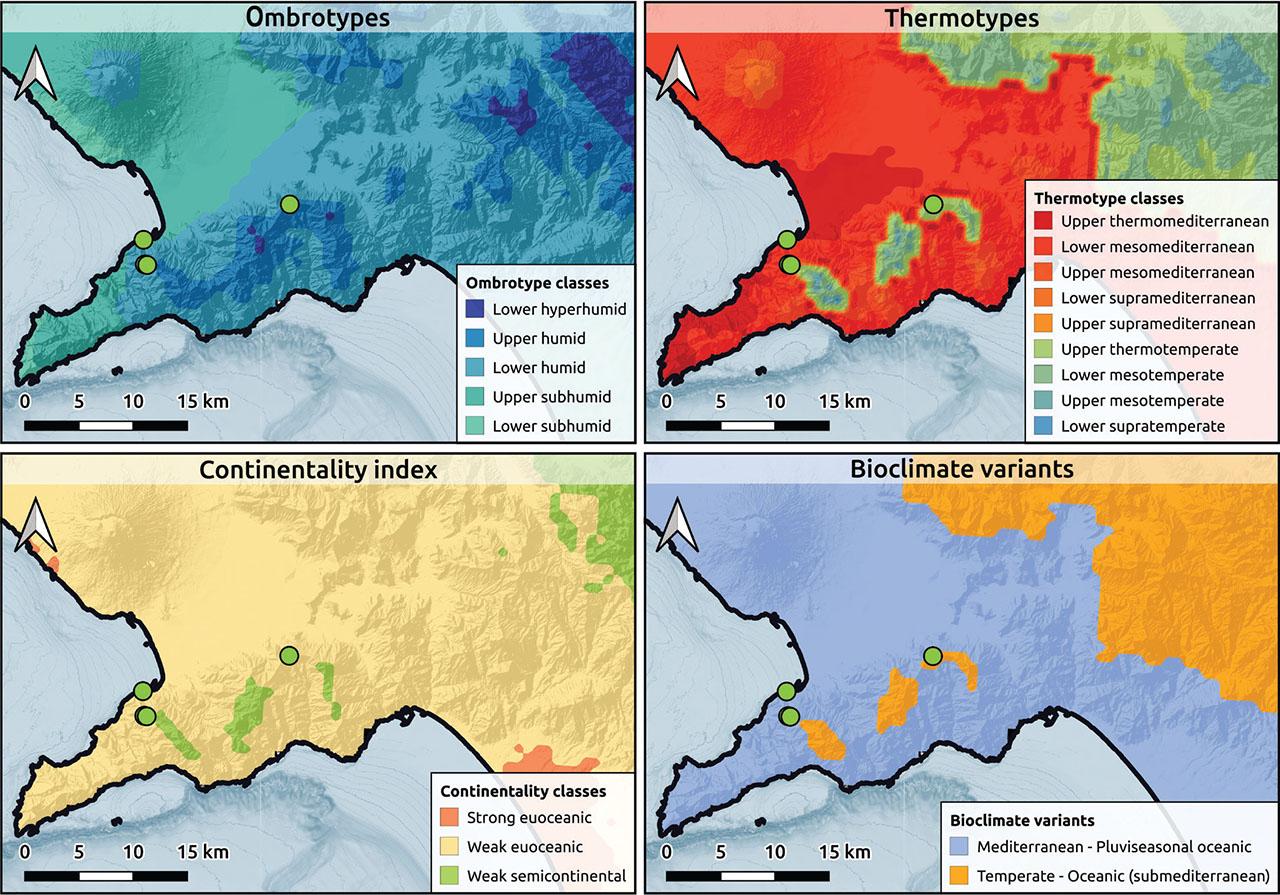
Bioclimatic maps from Pesaresi et al. (2017) for Diantho virginei-Seselietum polyphylli ass. nova.
Slika 5: Bioklimatski zemljevidi iz Pesaresi et al. (2017) za Diantho virginei-Seselietum polyphylli ass. nova.
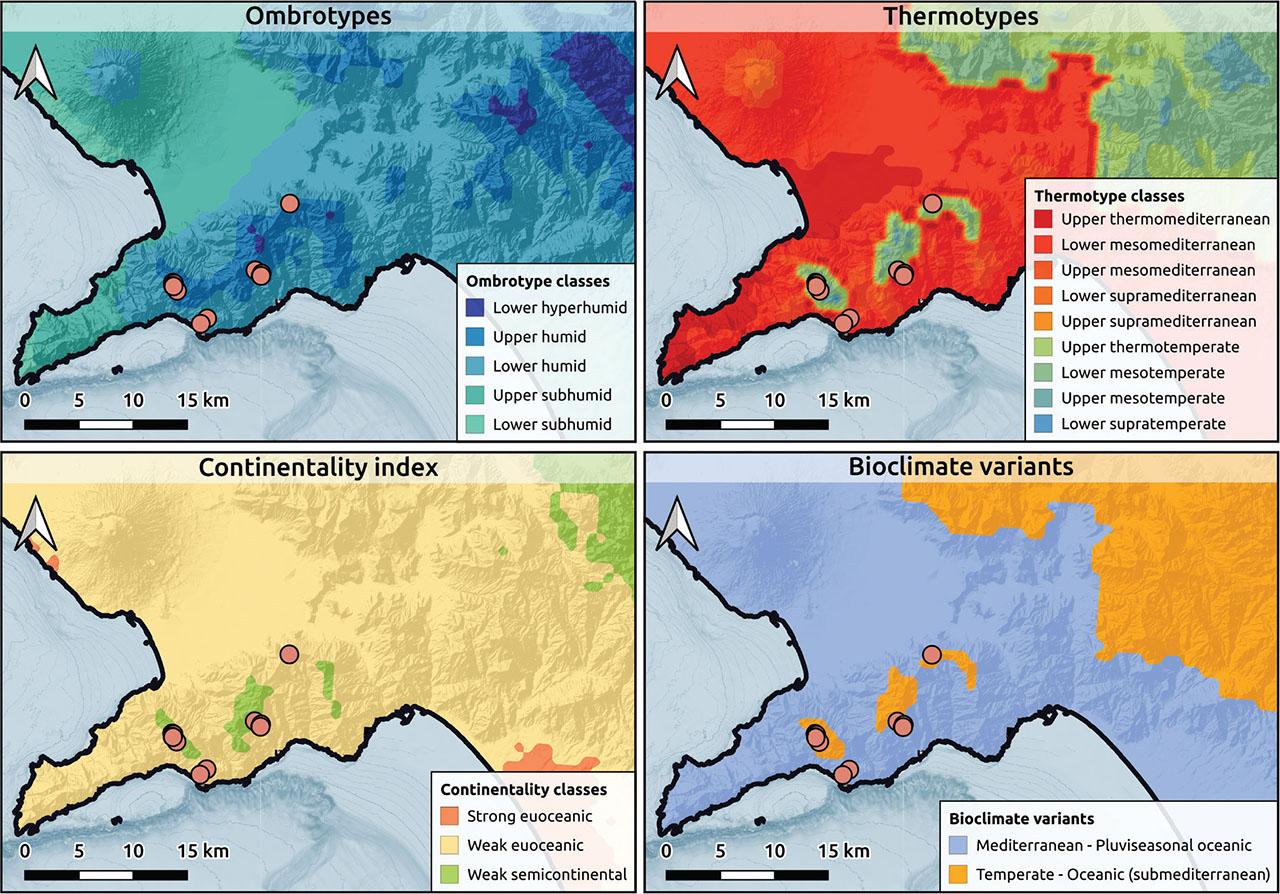
Bioclimatic maps from Pesaresi et al. (2017) for Globulario neapolitanae-Loniceretum stabianae typicum.
Slika 6: Bioklimatski zemljevidi iz Pesaresi et al. (2017) za Globulario neapolitanae-Loniceretum stabianae typicum.
The analysis identifies three clearly differentiated clusters of chasmophytic vegetation in the Sorrento Peninsula, dominated by Lonicera stabiana, Globularia cordifolia subsp. neapolitana, and Seseli polyphyllum, respectively. It seems reasonable to distinguish the investigated plant assemblages into two associations, one with Globularia cordifolia subsp. neapolitana and Lonicera stabiana, and one with Seseli polyphyllum. The latter association develops at slightly lower altitudes. Other researchers might view these three clusters as variants of the same association, but they are very distinct floristically and ecologically, to a degree that necessitates considering them as different syntaxa (see Figure 4).
The relevés dominated by Globularia cordifolia subsp. neapolitana and Lonicera stabiana represent two subassociations of the previously described association of Globulario neapolitanae-Loniceretum stabianae. The subassociation with dominance of Globularia cordifolia subsp. neapolitana represents a new subassociation, globularietosum neapolitanae, whereas the subassociation with dominance of Lonicera stabiana represents the subassociation typicum.
The subassociation globularietosum is typically found in the lower part of the slopes, suggesting a need for more soil accumulation at the base of the cliff, while the subassociation dominated by Lonicera stabiana (typicum) occupies the higher parts of the slopes and often upper parts of the cliffs, though there are significant overlaps.
The association dominated by Seseli polyphyllum is characterised by the prevalence of this species. Moreover, the floristic composition is different from that of Globulario-Loniceretum, as stressed by the cluster analysis and NMDS. Finally, the association grows at lower altitudes in a different bioclimatic context, as shown by the map (Figure 5). Dianthus virgineus is not a characteristic species but is frequently present in the association.
A group of stenoendemic species characterises the chasmophytic vegetation of Sorrento Peninsula (Lonicera stabiana, Globularia cordifolia subsp. neapolitana, Seseli polyphyllum, Centaurea tenorei, Galium lucidum subsp. venustum, Bromopsis caprina). The widespread species Sesleria juncifolia and Edraianthus graminifolius might be present with specific forms different from those in the Central Apennines (Fanelli et al., 2020). A study by Surina et al. (2014) has shown that populations of Edraianthus graminifolius from the Central Apennines are genetically distinct from those in Southern Italy. All these species allow for delineating a distinct alliance, Lonicerion stabianae, endemic to the Sorrento Peninsula and with a small exclave in the Picentini Mountains, where Globularia cordifolia subsp. neapolitana is present. These species represent true character species and are as such indicated in the synoptic table. We refrained from indicating differential species because a rigorous assessment of such species requires revision of the chasmophytic vegetation of the central Mediterranean, which is outside the aim of this paper, although a number of differential species are probably present, distinguishing the chasmophytic vegetation of the Sorrento Peninsula and Picentini from other chasmophytic vegetation of the Central Mediterranean but which are otherwise shared with dry grasslands.
A possible alternative hypothesis to the description of a new alliance is that the vegetation described in this paper falls within the Dianthion rupicolae, but this remains implausible for ecological, floristic, and phytogeographic reasons. As evident from the synoptic table, species from Dianthion rupicolae are virtually non-existent in the relevés of Lonicerion stabianae. If the alliances were synonymous, a substantial presence of Dianthion rupicolae species would be expected, given the geographical proximity to areas associated with this alliance. Indeed, ecologically, Lonicerion stabianae is distinguished through two features: it develops at relatively high altitudes (about 1000 m) with a temperate and weakly continental climate and growth on dolomitic substrates (Fanelli et al., 2020). Dianthion rupicolae develops in a thermo- to meso-mediterranean climate, with much more maritime conditions, and generally on non-dolomitic substrates (Terzi et al., 2018). The bioclimatic analysis shows that the alliance Lonicerion stabianae is characterised by a weakly continental climate and a predominantly temperate thermotype, which ecologically isolates this alliance within xerophytic cliff vegetation (Terzi et al., 2018). In this context, Diantho virginei-Seselietum polyphylli occupies a somewhat intermediate position with Dianthion rupicolae. In fact, Seseli polyphyllum occurs sometimes with low frequencies in a few associations of Dianthion rupicolae.
The synoptic table does not reveal any affinity with Asplenietalia glandulosi at the order level, while a moderate affinity with the Adriatic Centaureo-Campanuletalia appears. Species from Asplenietalia glandulosi are absent, but two species of Centaureo-Campanuletalia, in particular Cytisus spinescens and Sesleria juncifolia, are present. This is somewhat surprising, since Asplenietalia glandulosi is the order of central Mediterranean chasmophytic vegetation, whereas Centaureo-Campanuletalia is fundamentally distributed in the Adriatic.
Only a few species of Centaureo-Campanuletalia were encountered in our surveys, but this could be explained by the very marginal position of the Sorrento Peninsula for the main area of Centaureo-Campanuletalia. This is not surprising since there is a strong disjunction between the Sorrento Peninsula and the main area of Centaureo-Campanuletalia in the Adria. In summary, the affinities of the mountainous zone of the Sorrento Peninsula seem to be eastern, while the lower zones have a distinctly more western character.
Our results are based on the analysis of the synoptic table and bioclimatic characterisation. A numerical analysis of cliff vegetation in the central Mediterranean could further support these findings, as such analyses are now relatively easy thanks to the integration of large databases (Chytrý et al., 2016). However, this type of analysis has already been conducted by Terzi et al. (2018), and adding relevés from the Sorrento Peninsula would likely be redundant. Moreover, the scope of our study is to describe the vegetation of the Sorrento Peninsula and align it within the framework of known vegetation as presented in the literature. This approach follows the standard method in phytosociology, which begins with a detailed description of the vegetation at the local scale and, only after a substantial amount of data has been collected, proceeds to numerical analysis and revision (e.g., Bonari et al., 2020).
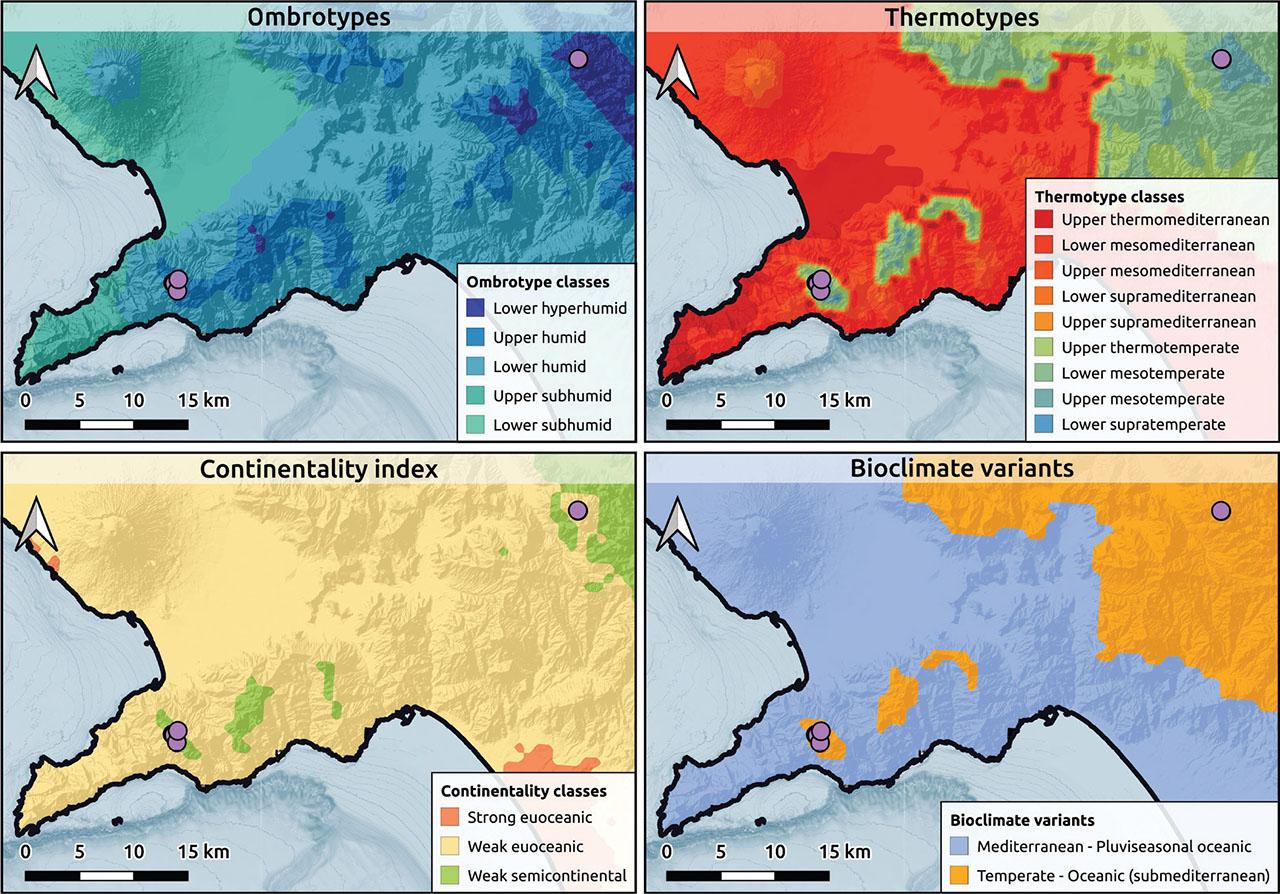
Bioclimatic maps from Pesaresi et al. (2017) for Globulario neapolitanae-Loniceretum stabianae globularietosum neapolitanae subass. nova.
Slika 7: Bioklimatski zemljevidi iz Pesaresi et al. (2017) za Globulario neapolitanae-Loniceretum stabianae globularietosum neapolitanae subass. nova.
In conclusion, we propose the following syntaxonomical scheme:
Globulario neapolitanae-Loniceretum stabianae globularietosum neapolitanae subass. nova hoc loco (Holotypus n. 24)
Globulario neapolitanae-Loniceretum stabianae typicum Fanelli et al. 2020
Diantho virginei-Seselietum polyphylli ass. nova hoc loco (Holotypus n. 28) Notes:
Lonicerion stabianae all. nova hoc loco (Holotypus Globulario neapolitanae-Loniceretum stabianae).
Centaureo-Campanuletalia Trinajstić 1980
Asplenietea trichomanis Br.-Bl. in Meier et Br. Bl. 1934) Oberd. 1977
The Sorrento Peninsula represents a significant centre of endemism, which Italian botanists have somewhat overlooked from a biogeographic perspective, with a few exceptions (Guadagno, 1916; Salerno et al., 2007). Our proposal to describe an alliance for this restricted biogeographic sector, aimed at integrating renewed ecological and phytogeographic foundations, successfully increasing the number of relevés for Globulario neapolitanae-Loniceretum stabianae and identifying the subassociation globularietosum also in the Picentini Mountains.
In the standard classification of the ecoregions of Italy (Blasi et al., 2018), the Sorrento Peninsula is not distinguished from the Cilento subsection, which has a very different floristic setting, with iconic species such as Eokochia saxicola, Limonium remotispiculum and Primula palinuri. The main distinction between the Sorrento Peninsula and Cilento is climatic, with the latter showing a mainly a thermo-mediterranean climate.
The study's findings contributed to drawing attention to the uniqueness of this peculiar territory. The Sorrento Peninsula is an island of endemism comparable to other Italian plant diversity hotspots (e.g. the Apuan Alps, Belluno Dolomites, Aspromonte, Etna, etc.), which, despite a very limited size, hosts a considerable number of rare species.
According to our analysis, the Sorrento Peninsula should have more affinities with the Adriatic ranges than with the western Mediterranean or the Tyrrhenian domains. This is relatively not surprising when considering the climate of the Sorrento Peninsula. Unlike neighbouring regions, the highest tops of Sorrento Peninsula is more exposed to the winter currents of cold air coming from the Balkan spots, as only partially shielded by the Picentini Mountains (Capozzi et al., 2023). This is reflected in the weakly continental bioclimate classification of the higher altitudes of the Lattari and Picentini Mountains (Pesaresi et al., 2017). Conversely, in summer, the very close maritime currents, especially in the lower zones, ensure a milder climate enabling the plants to flourish. A more detailed study of the lower cliff zone would be interesting, although made difficult by the extensive urbanisation of the area.