
Osmoprotectants (compatible solutes, osmolytes) are molecules that balance the osmotic difference between the cell environment and the cytosol without interfering with the enzymatic function of proteins or other biomolecules (Omara et al., 2020; Slama et al., 2015). They can be classified into the following chemical classes: polyamines (e.g. putrescine, spermidine), sugars (e.g. sucrose, trehalose), sugar alcohols (e.g. inositol), betaines (e.g. glycine betaine) and amino acids (e.g. proline, ectoine) (Kido et al., 2019). Betaine is a naturally occurring sugar with osmotic properties. It is synthesised in the cells of microorganisms (bacteria, archaea, fungi) to protect the cells from drought, osmotic and temperature stress (Zou et al., 2016). Betaine is also produced in some plants, including sugar beet, where it usually accounts for around 3–8% of the dry weight of beet molasses (Abdollahzadeh et al., 2022). Ectoine, an amino acid described as the “anti-stress molecule of the future” (Stępniewska et al., 2014) is synthesised by a small number of bacterial species, mainly halophilic bacteria, which are resistant to salt and temperature stress thanks to it (Stöveken et al., 2011). Inositol, on the other hand, is a sugar alcohol produced by plants and some microorganisms in response to physiological stress conditions (Kido et al., 2013; Mukherjee et al., 2019). The bacteria best known for osmoprotectant production are halophilic and halotolerance, which produce ectoine, glycine betaine, among others (Tanimura et al., 2013). In plants exposed to abiotic stresses (salinity/drought), osmoprotectants help maintain cell turgor and drive the water uptake gradient (Kido et al., 2019). Studies have shown that bacterial osmoprotectants can enhance plant resistance to drought (Morcillo et al., 2021). Their synthesis by microorganisms also occurs in flooded environments (Patel et al., 2021). The use of bacteria capable of producing osmoprotectants as bioinoculants can also help plants sensitive to abiotic stress (Fouda et al., 2021).
Wheat is one of the staple cereals grown around the world. Spring wheat, due to its underdeveloped root system, is a crop susceptible to drought stress and, in addition, studies have shown that for every unit increase in soil salinity, the number of seeds, ears, 1000 grain weight and economic yield of wheat are significantly reduced (Eynard et al., 2005; Zhang et al., 2024). Studies have shown that soil salinity causes a decrease in wheat yield of about 20%–43% in different regions of the world (Mustafa et al., 2019). At the same time, soil salinity is an important factor limiting the area of cultivated soils, productivity and crop quality, while the extent of saline soils is increasing (Corwin, 2021).
Ongoing climate change is forcing the search for new adaptation solutions in agriculture (Zhao et al., 2022). Drought and salinity are among the major problems for agriculture (Rontein et al., 2002). Some plants produce osmoprotectants, but in smaller quantities than microorganisms. Genetic engineering has allowed the introduction of osmoprotectant pathways into plants, improving their stress tolerance, but transgenic plants showed variable levels of tolerance due to many metabolic limitations (Rontein et al., 2002; Zulfiqar et al., 2020). Therefore, attempts have been made to add exogenous osmoprotectants either directly to the soil or to the leaves at the time of stress (Ashraf, Foolad, 2007). Among others, studies (Sabagh Ayman et al., 2016) showed that adding exogenous osmoprotectants (proline, glycine betaine) to the soil improved proline and nitrogen content in soybean leaves and increased seed yield under drought stress conditions. Research (Repke et al., 2022) has shown that the use of the glycine betaine additive in soybeans increases plant drought tolerance. In maize exposed to cadmium stress, the addition of ectoine had a positive effect on plant growth, water potential and antioxidant capacity (Ozfidan-Konakci et al., 2022). The addition of myo-inositol was shown to alleviate salt stress in quinoa by engaging antioxidants, improving plant growth characteristics and reducing oxidative damage (Al-Mushhin et al., 2021). Similarly, the addition of exogenous myo-inositol to salt-treated early seedlings of Mesembryanthemum crystallinum L. (ice plant) increased leaf biomass, relative water content and chlorophyll content (Li C.H. et al., 2023).
Following the positive reports, we wanted to test whether osmoprotectants (betaine, ectoine, and inositol) added to the culture medium would enable the spring wheat (Triticum aestivum L.) to grow better under osmotic stress conditions induced by the addition of sodium chloride.
A model experiment was carried out using glass jars (V = 900 mL) with universal plant culture Hoagland medium in 2023. The medium was prepared using reagents purchased from Merck according to the recipe described by Parven et al. (2022). The medium was modified by adding agar at a rate of 17 g per 1000 mL of medium to obtain a solid medium. 150 mL each of sterile medium was poured into sterile jars and allowed to set under sterile conditions.
Two experimental variants were used: (1) standard medium; (2) stress medium. The standard medium refers to the Hogland solid medium, while the stress medium was prepared by adding sterile, free-flowing NaCl (Sigma Aldrich) to the medium in an amount allowing a final concentration of 1% (m/V) (Nazarov et al., 2022). This study aimed to test the effect of osmoprotectants on plants under high stress conditions – hence a concentration of 1% (m/V), corresponding to about 0,17 M L−1 (Kłosowska, 2010). A scheme of the experiment is shown in Figure 1.
Scheme of experiment (author's elaboration).
Triticum aestivum L. var. Telimena kernels (provided by Danko) were sown in vessels prepared as described previously. Wheat seeds were surface sterilised with 95% ethanol to exclude possible substrate contamination according to the method described by Błaszczyk et al. (2021). Ten seeds were sown in each glass. Five biological replicates were prepared for each variant of the experiment (n = 40).
The osmoprotectants were dissolved in the medium before sowing the plants. Three different commercially available osmoprotectants of microbial origin were selected: betaine, ectoine, inositol (Sigma Aldrich). The concentration of the betaine and inositol was estimated to be 25 mM (Ghorbani et al., 2023; Li C.H. et al., 2023); when of the ectoine it was 0.25 mM based on literature data (Ma et al., 2023).
All jars with plants were placed in a growth chamber (Heraeus, Vötsch, Hanau, Germany) maintained at 20 °C in the daytime (16 h) and at 19 °C during the night (8 h). The humidity was maintained at 60%. After 17 days the number of germinated seeds, fresh and dry plant weight, average root length and average shoot length were determined. Germination capacity was calculated according to Kaur and Gill (2020). Plant weight and root and shoot length were determined on all plants germinated in a given jar (different n, from 4 to 10) and averaged per jar, and then biological replicates were averaged for the entire combination (n = 5). For the determination of the dry weight of the plants, collected wheats were dried at 55 °C for 48 h and then weighed. The dry matter was measured with an accuracy of 0.0001 g and then averaged over each combination.
Statistical analyses were performed using Statistica ver. 10.0 (StatSoft. Inc., Tulsa, OK, USA). Diagrams were performed using MS Excel software (Microsoft Corporation, 2016).
Analysis of the germination capacity of wheat seeds in the presence of added osmoprotectants showed that the addition of ectoine resulted in the highest number of germinated plants and inositol in the lowest, although still higher than the control (Table 1). It is noteworthy that even under salt stress conditions, the addition of ectoine and betaine resulted in 25% more sprouts being obtained (Figure 2A). However, these differences were not statistically significant (at p < 0.05).
Germination capacity [%] of Triticum aestivum L. seeds in different variants of the experiment.
| Medium | Control | Betaine | Ectoine | Inositol |
|---|---|---|---|---|
| Hoagland medium | 40.0 | 50.0 | 55.0 | 46.7 |
| Hoagland medium with NaCl (salt stress) | 40.0 | 50.0 | 50.0 | 43.3 |
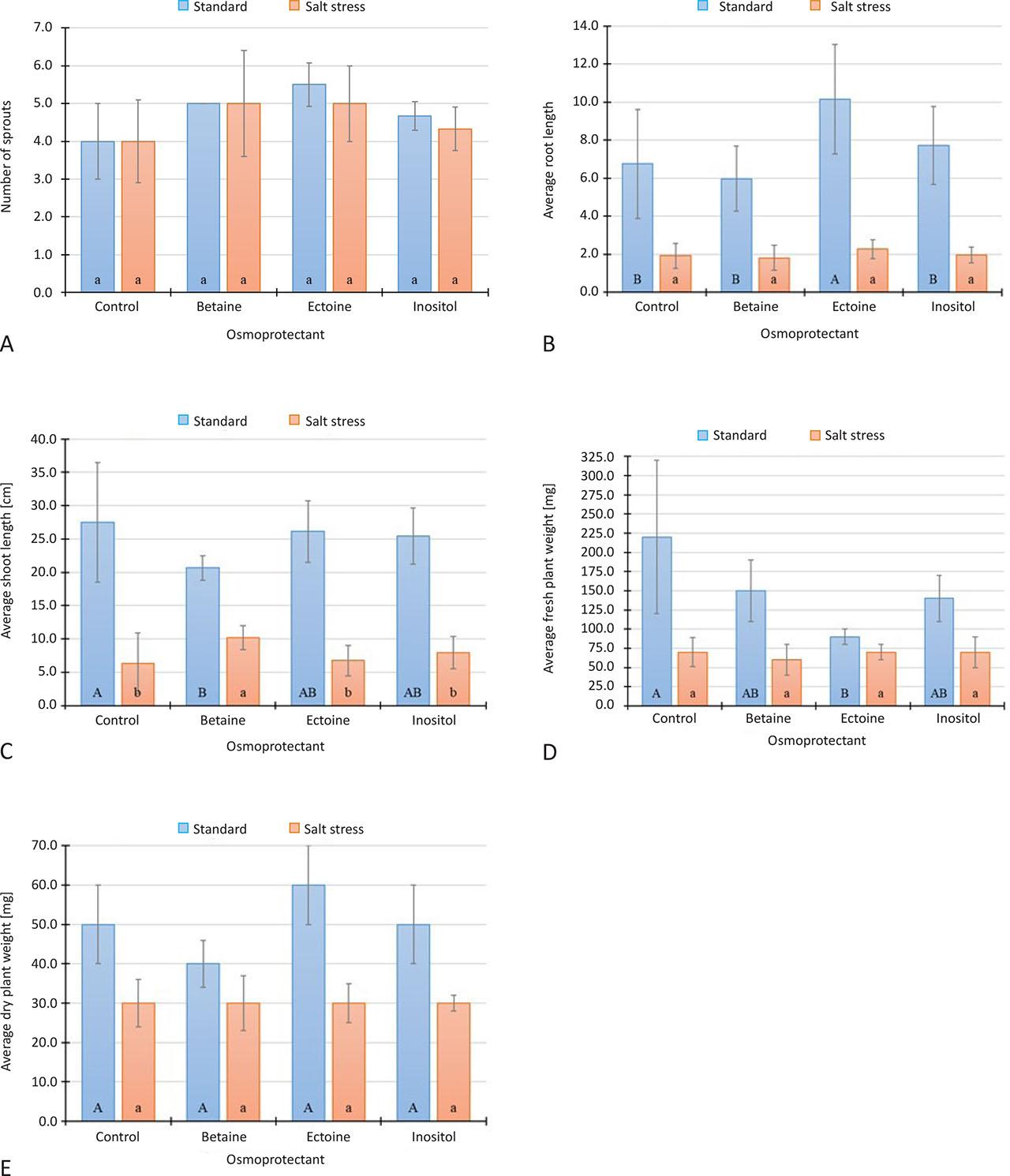
Plant parameters determined in the experiment after 17 days of growing plants on media with addition of selected osmoprotectants.
control – culture without addition of osmoprotectants; standard – Hoagland medium without NaCl; salt stress – Hoagland medium with NaCl (1% m/V); vertical lines indicate SE; Different lowercase letters ‘a–b’ indicate a significant difference for plants growing under salt stress conditions; different uppercase letters ‘A–B’ indicate a significant difference for plants growing on standard medium; both at p < 0.05, n = 10, Tukey HSD test, ANOVA.
The average root length of the plants showed a positive effect of the addition of ectoine and inositol (Figure 2B). The addition of ectoine to the medium resulted in 50.4% longer roots under optimum conditions and 18.7% longer roots under salt stress conditions. The effect of the addition of ectoine on wheat root length was statistically significant for plants grown on standard medium (at p < 0.05).
In the optimal conditions, the addition of betaine resulted in a statistically significantly lower values of shoots length compared to the control (Figure 2C). However, under the salt stress condition shoots were longer by an average of 3.8, 0.4, and 1.6 cm (i.e., 60.2, 6.0 and 25.0%) after the addition of betaine, ectoine and inositol, respectively, compared to the culture without osmoprotectants, but only for betaine the values were statistically significant.
For fresh weight, there was no positive effect of the addition of osmoprotectants (Figure 2D). Surprisingly, the fresh weight of the plants after application of ectoine was statistically significantly lower than the control (no added osmoprotectants). There were no statistically significant differences under salt stress conditions.
The addition of ectoine resulted in a higher dry weight of wheat under optimum conditions compared to the control sample (Figure 2E). This difference was 10 mg (i.e., 20%), but not statistically significant. No statistically significant differences were observed in the salt stress variant either.
Triticum aestivum L. is a staple crop for one third of the world's population and its production is of great economic and food security importance (Singh et al., 2021). Good quality wheat seed can have a germination capacity of up to 100% (Khaeim et al., 2022). In the present study under control conditions (without NaCl addition and without osmoprotectants) only 40% was obtained. Such a low result is surprising and may indicate the poor quality of the seeds used. Poor seed quality may also affect other plant growth parameters (Nuttall et al., 2017). Throughout the experiment, seeds from the same batch were used, which makes it possible to compare the results obtained.
Salt stress conditions simulated by adding NaCl to the medium (to end concentration 1% m/V) resulted in poorer plant growth, as indicated by lower values for root and shoot length, dry and fresh plant weight, compared to plants growing under optimum conditions. Wheat is generally sensitive to salinity, which inhibits plant growth and development and leads to low productivity (EL Sabagh et al., 2021). Four weeks of salinity stress (150 mM NaCl) caused significant changes in the physiological and agronomic characteristics of wheat – plant growth and biomass production – resulting in a significant reduction in yield (Tao et al., 2021). In presented study reductions in fresh and dry matter under salinity were also observed, by 68.2 and 40% respectively, compared to plants grown on medium without salt stress.
Betaine is an important plant osmoprotectant that enhances plant stress tolerance by improving leaf gas exchange, leaf chlorophyll content, activation of the antioxidant system and protection of cellular structures (Shemi et al., 2021). In this study, the addition of betaine increased the number of germinated seeds by 25% under salt stress and in optimal conditions, confirming the observations of other researchers using different test plants. For example, in experiments with maize, researchers confirmed that the addition of betaine increased germination (by 38.4%) and shoot-root biomass under salt stress conditions (Rhaman et al., 2024). Studies have shown that the addition of glycine betaine significantly increased seed germination and vigor of millet seeds (Lavanya, Amruthesh, 2017) and increased sunflower salt tolerance (Roychoudhury, Tripathi, 2020). Wheat growing with betaine added, carried out by Ahmed et al. (2021) resulted in less root reduction under water stress compared to the unstressed control. In addition, the researchers obtained 2.3% longer shoots from plants growth with betaine addition, and in the results discussed here, up to 60.2% longer shoots were obtained under salt stress conditions compared to plants without the osmoprotectants and the values were statistically significant (at p < 0.05). A surprising result is the lower value of fresh plant weight under optimum conditions obtained after betaine addition compared to control without addition. This is in contrast to the results obtained by other researchers, where the fresh weight of plants/shoots/roots was higher after betaine application than without betaine addition (Xing, Rajashekar, 1999; Lin et al., 2020). Also, in an experiment with Tartary buckwheat, the fresh weight of shoots was statistically significantly higher after the addition of 20 mM betaine hydrochloride compared to the control (Kim et al., 2020). However, the study also showed that the fresh weight of sprouts with the addition of 30 mM betaine was not significantly different from that of sprouts without the addition of the osmoprotectant. It can be assumed that the low fresh weight value may have been influenced by the quality of the seeds themselves or that the betaine concentration used (25 mM) was too high for this wheat cultivar and caused growth inhibition. This subject definitely deserves further investigation.
Ectoine protects proteins and cell membranes from dehydration and high salinity damage and helps maintain ionic homeostasis (Ma et al., 2022). In the present study, the addition of ectoine was found to have a positive effect on plant germination, root length and plant dry weight, with only root length being statistically significant compared to the unsupplemented control on standard non-stressed medium. A surprising result is the significantly statistically lower value of fresh plant weight under optimum conditions obtained after ectoine addition. This is in contrast to the results obtained by other researchers, where the positive effect of ectoine supplementation on plant growth has been reported. Addition of ectoine improved seed germination, photosynthetic rate and sodium accumulation under salt stress conditions in flax (Linum usitatissimum) (Elsakhawy et al., 2019). Other studies have shown that the addition of ectoine ensure efficient water management of maize plants under Cd stress conditions, which also induces osmotic stress (Ozfidan-Konakci et al., 2022). In the present study, a 25% higher number of germinated wheat seeds was recorded at a dose of 0.25 mM ectoine compared to the control under salt stress. Research conducted by Ma et al. (2023) showed that the addition of 0.25 mM ectoine under salt stress conditions recorded an 81.3% increase in cotton germination compared to the control. The researchers also used a dose of 0.50 mM ectoine, which resulted in 129.8% more germinated cotton plants compared to the control. Ectoine at both doses also contributed to higher root dry weight compared to the control without osmoprotectant, while in our study the addition of ectoine was associated with 18.7% longer wheat roots compared to the control, but the values were not statistically significant.
Inositol is a common small molecule found throughout the biological world and acts as an osmoprotectant, protecting plants from osmotic stress and oxidative damage and enhancing various metabolisms to adapt to NaCl stress (Li et al., 2023). Research conducted by Fatima et al. (2024) showed that the addition of inositol increased maize growth under saline conditions by improving chlorophyll content and photosynthesis compared to plants without inositol. The researchers recorded up to 72% higher root dry weight after application of 50 mg L−1 of inositol. In presented study, the wheat showed only 2.6% longer roots and 25% longer shoots after inositol addition compared to plants without the added osmoprotectant under saline conditions, but the values were not statistically significant. The study used 10 mM inositol showed an increase of 17.1 and 9.2% in shoot fresh and dry weight and 10.1 and 8.5% in quinoa (Chenopodium quinoa L.) root fresh and dry weight (Al-Mushhin et al., 2021).
The results show that exogenous application of selected osmoprotectants can mitigate the adverse effects of salinity and promote growth of spring wheat. The addition of each osmoprotectant was shown to have a positive effect on wheat seed germination under both optimal and salt stress conditions, although the differences were not statistically significant. Addition of ectoine (0.25 mM) showed a statistically significant positive effect on wheat root length under control conditions, and addition of betaine (25 mM) showed a statistically significant positive effect on shoot length under salt stress conditions.
Surprisingly, the addition of betaine, ectoine and inositol resulted in a lower fresh weight of wheat compared to the control under unstressed conditions. For ectoine this differences was statistically significant. This is even more unexpected as the number of germinated plants was higher after the addition of osmoprotectants and literature data do not indicate toxicity of these compounds at the concentrations used in the experiment.
The results highlight the need for further investigation into the use of exogenous osmoprotectants. The diversity of the test plant and the different concentration variants of the additives used deserve special attention in further research.