
Accumulating data have shown a pathophysiological association between inflammatory pathways and thrombosis. Venous thromboembolism (VTE), which includes deep vein thrombosis (DVT) and acute pulmonary embolism (APE), is a significant health burden (Konstantinides et al. 2019). Epidemiological data indicate that the prevalence of VTE in Europe is about 1–1.8 per 1000 inhabitants (Heit 2015). Despite significant advances in anticoagulation, 10%–25% of patients still experience recurrent VTE in 5-year period, indicating the limited management of pathogenic mechanisms (Kahn et al. 2014). Therefore, novel therapeutic options are warranted.
It is common knowledge that pathophysiology of APE is complex and multifactorial, leading to the concept of mosaic theory, as indicated in our previous review (Imiela et al. 2024). It involves not only hemodynamic disturbances due to the emboli occluding the pulmonary arteries, but also platelet activation, endothelial dysfunction, and “firing up” of the inflammatory cascade (Piazza 2015; Dzikowska-Diduch et al. 2017; Jackson et al. 2019; Mosquera and Diaz 2023; Imiela et al. 2024).
APE typically occurs when a thrombus enters the pulmonary circulation, usually originating from the lower extremities. The clot obstructs blood flow, increasing resistance in the pulmonary arteries, which elevates right ventricular pressure. This can ultimately lead to acute myocardial damage, necrosis, and hypotension. The lysis of monocytes in the RV promotes a pro-inflammatory response. In clinical practice, high-risk patients are hospitalized in the Emergency Department and evaluated by specialized Pulmonary Embolism Response Teams to provide immediate care and improve outcomes. APE can also result from the migration of fat, air, or more often from tumor cells (Poisner et al. 2018).
APE can lead to cardiac shock, resulting in hypoperfusion, hypoxia, and more exaggerated clot formation, which is why VTE is considered to be a nonspecific inflammatory state (Abuduhalike et al. 2020; Konstantinides et al. 2020). The interplay between inflammatory state and thrombosis was first reported in the early 1970s (Charkes et al. 1974). Generally, inflammation leads to the upregulation of platelet and fibrinogen reactivity, tissue factor (TF) formation, and membrane phospholipid activation (Mosquera and Diaz 2023). Conversely, inflammation inhibits fibrinolysis and downregulates modulin expression (Mosquera and Diaz 2023).
The physiological role of inflammation can be more exaggerated during infection when the host response, known as immunothrombosis, plays a protective role in limiting pathogen dissemination through an intricate interplay between the endothelium, platelets (PLT), and innate immunity (Engelmann and Massberg 2013; Kimball et al. 2016). However, excessive immunothrombosis usually leads to hypercoagulability, which is particularly evident during acute COVID-19 infections (Bonaventura et al. 2021; Machowski et al. 2024).
Another important player in immunothrombosis is neutrophils. Neutrophils are essential markers of inflammation and have been linked to VTE (Peng et al. 2023). Neutrophil levels are significantly higher in patients with intermediate- and high-risk APE compared to those with low-risk or non-APE (Peng et al. 2023). Neutrophil levels are positively correlated with the Pulmonary Embolism Severity Index (PESI), PAOI, D-dimer levels, and high-sensitive C-reactive protein (CRP), suggesting their strong association with both inflammation and thrombosis in APE (Kimball et al. 2016; Peng et al. 2023). In addition, neutrophil extracellular traps (NETs) are structures composed of DNA, histones, and antimicrobial proteins released by neutrophils (Natorska et al. 2023). This process, known as NETosis, is distinct from other forms of cell death such as necrosis and apoptosis. NETs play a crucial role in trapping and killing bacteria, but they also contribute to thrombosis by promoting platelet adhesion, activation, and aggregation (Natorska et al. 2023). Studies using murine models have shown that NETs contribute to thrombus formation by interacting with PLT and the coagulation cascade. The presence of NETs has been confirmed in human VTE, where they are associated with acute thrombotic events and may serve as potential biomarkers for diagnosis (Natorska et al. 2023).
Immunothrombosis refers to the process by which the immune system contributes to thrombosis, particularly in the context of infection or inflammation (Natorska et al. 2023). NETs play a dual role in this process: they help contain and eliminate pathogens but can also lead to pathological thrombosis if dysregulated (Kimball et al. 2016). There are potential therapeutic targets for preventing NET-mediated thrombosis, such as inhibiting peptidylarginine deimanse 4 (PAD4), an enzyme essential for NET formation, or using DNase to degrade NETs (Kimball et al. 2016; Natorska et al. 2023). Existing therapies like heparin and aspirin are also highlighted for their secondary effects on NETs, offering possible avenues for enhancing current treatments.
Despite studies on inflammation conducted in experimental models of APE, there is still a paucity of data from human studies (Imiela et al. 2024). Most therapeutic options for VTE target the coagulation cascade. In the clinical scenario, neither nonvitamin K antagonists nor direct oral anticoagulants (DOACs) can prevent recurrent VTE (Di Nisio et al. 2016).
The results of the study—Canakinumab Anti-Inflammatory Thrombosis Outcome Study (CANTOS trial) indicated that an anti-interleukin (IL)-1 specific antibody can reduce the risk of VTE in clinical settings, highlighting the need to address this therapeutic gap (Ridker et al. 2017). Additionally, other drugs, such as the nonselective neutrophil-to-lymphocyte ratio (NLR) family pyrin domain containing 3 (NLRP3) inhibitor colchicine and lipid-lowering drugs, reduce primary VTE events, creating a therapeutic milieu for new antithrombotic strategies (Potere et al. 2023).
In this review, we summarize clinical evidence on the emerging role of key immune system markers in APE pathophysiology.
Venous thrombus formation causes vein wall injury and can lead to thrombus dislocation, resulting in pulmonary artery (PA) embolism. This induces a robust inflammatory state not only in the PA wall, but also in the right ventricle (RV), as described in numerous animal models of APE (Zagorski et al. 2003, 2009; Zagorski and Kline 2016; Imiela et al. 2024). Traditionally, the clinical course of APE and its potential consequences, such as RV failure, are thought to be parallel to hemodynamic changes and embolism size. However, recent studies have revealed that APE pathogenesis is complex, involving not only hemodynamic aberrations but also thrombogenesis, endothelial cell dysfunction, and inflammation (Bontekoe et al. 2021; Darwish et al. 2022). All mentioned processes result finally in APE progression (Figure 1).
The process of thrombosis is due to the activation of numerous cells, mainly endothelial cells, and PLT, however, it can be postulated that leukocytes activation and inflammatory cytokine release, trigger the coagulation cascade (Darwish et al. 2022). Additionally, established embolisms have been shown to be a source of inflammation. While there are easily accessible biomarkers for APE stratification, such as high-sensitivity Troponin T levels or N-terminal pro-brain natriuretic peptide (NT-proBNP), which indicate RV failure, specific biomarkers dedicated to APE stratification are still lacking (Konstantinides et al. 2019).
Several clinical biomarkers can be considered for APE risk stratification and prognosis. In the mid-1980s, the prognostic role of white blood cell (WBC) count in mortality from coronary artery disease was reported (Grimm et al. 1985). However, the clinical utility of WBC concentration in VTE management remains limited. In humans, the systemic inflammatory state can also be evaluated using plasma levels of CRP and IL-6, which correlate with venous obstruction, thrombus extension, and clinical VTE complications such as postthrombotic syndrome, recurrent thromboembolism, worse quality of life, and functional impairment (Roumen-Klappe et al. 2009; Rabinovich et al. 2015; Potere et al. 2023).
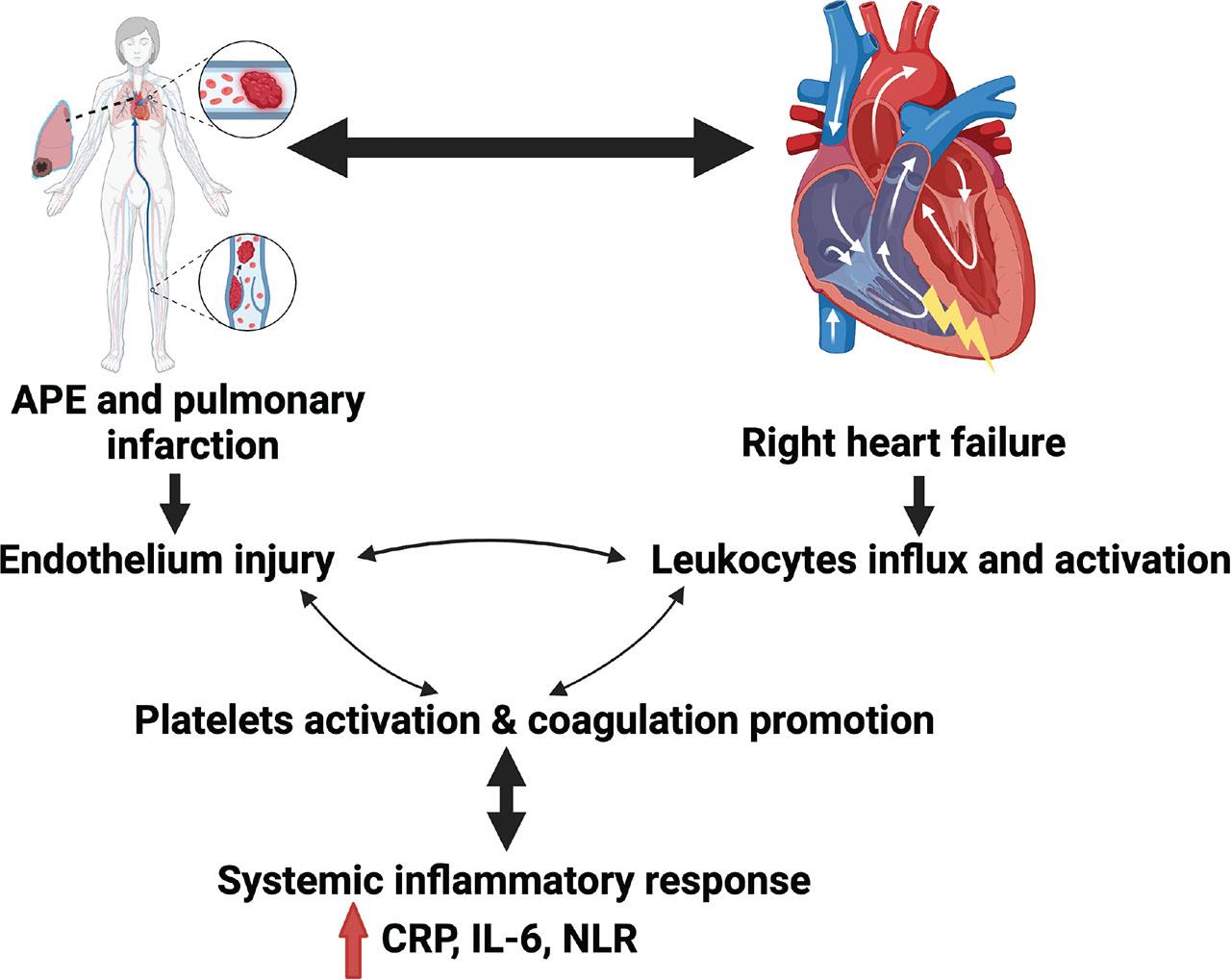
General view on APE pathogenesis. The figure presents the concept of the role of immune system and inflammation as a substrate in APE development and progression. APE, acute pulmonary embolism; CRP, C-reactive protein; IL-6, interleukin-6; NLR, neutrophil-to-lymphocyte ratio. This figure has been created with BioRender.com.
The exaggerated inflammatory state during postthrombotic syndrome aligns with severe alterations in endothelial function, such as activation of intercellular adhesion molecule (ICAM)-1 and E-selectin, as well as vascular proteolysis and fibrinolysis. Moreover, a hypercoagulable state, indicated by higher levels of von Willebrand factor (vWF) and factor VIII, is closely associated with the inflammatory response (Dzikowska-Diduch et al. 2017; Bittar et al. 2020; Potere et al. 2023). We aimed to describe the role of basic inflammatory markers in daily clinical practice. These markers can provide valuable insight into the interplay between thrombosis and inflammation, helping inform better management and treatment strategies (Figure 2).
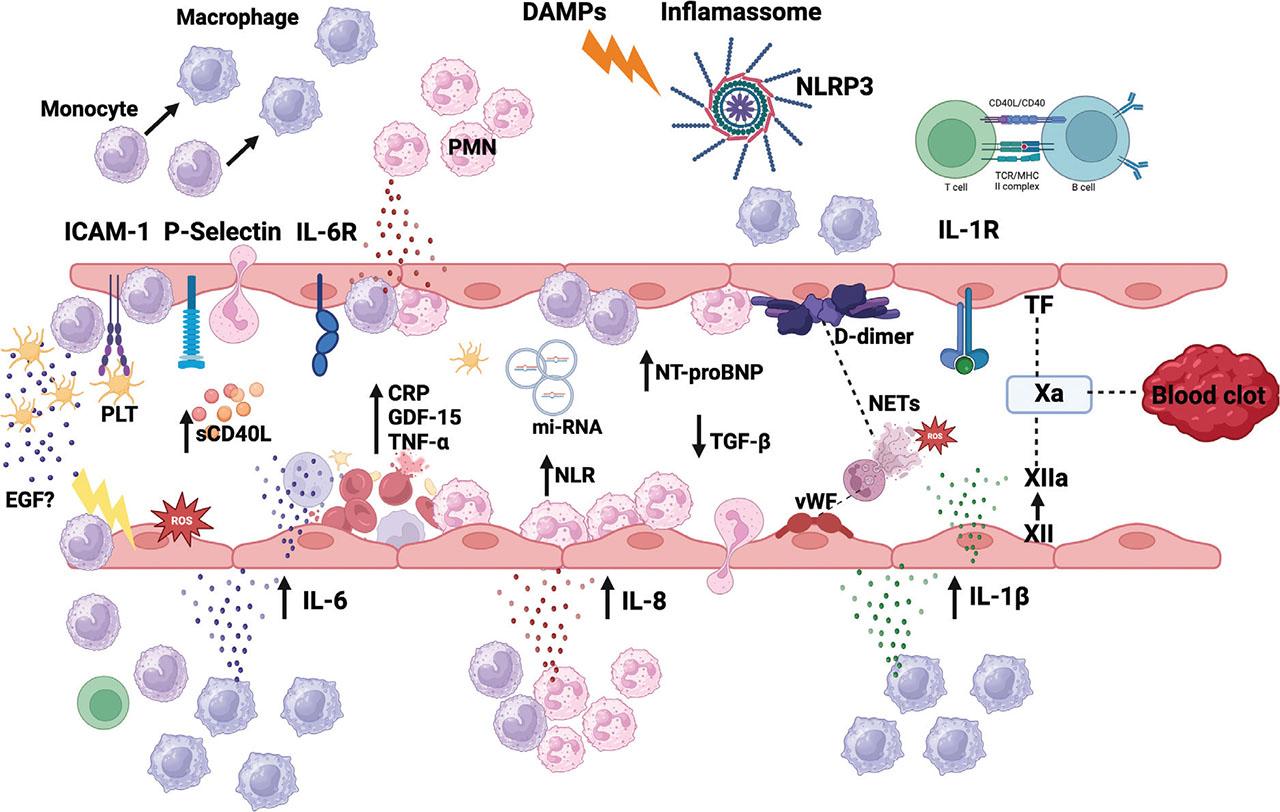
Schematic of immune reactive oxygen species (ROS) response in vessel during VTE. Early in thrombogenesis, adhesion molecules initiate leukocyte accumulation, and adhesion to endothelium for subsequent platelet accumulation. Endothelium injury releases DAMP. Proinflammatory cytokines and ROS, promote inflammasome activation in innate immune cells, primarily macrophages, and neutrophils. IL-1β promotes leukocytes adhesion as well as neutrophils and PLT recruitment. IL-6 is released by infiltrating leukocytes and monocytes and induced endothelial IL-1 production and expression of adhesion molecules and prothrombotic factors, facilitating further platelet recruitment. Systemic inflammatory state can be measured by higher concentration of circulating GDF-15, IL-6, TNF-α. As a response to proinflammatory state and higher IL6 level, CRP is synthesized by the liver. IL-6 also promotes lymphocytes T activation. DAMP and PAMPs are presented to lymphocytes T, resulting in higher sCD40 in bloodstream. Activation of platelet and macrophage NLRP-3 inflammasome induces exaggerated production of IL-1 and NETosis formation. The interaction between NETs DNA and vWF, leads to further thrombus growing. Inflammasome activation results in release of TF, which leads to thrombin generation and fibrin clot formation. Neutrophil express TF resulting in extrinsic pathway activation, however, NETs lead to factor XII activation and intrinsic pathway activation, finally contributing to thrombus development and growth. CRP, C-reactive protein; DAMP, danger-associated molecular pattern; EGF, epidermal growth factor; GDF-15, growth differentiation factor-15; ICAM, intercellular adhesion molecule; IL, interleukin; miRNAs, micro-RNAs; MHC, Major histocompatibility complex; NETs, neutrophil extracellular traps; NLR, neutrophil-to-lymphocyte ratio; NLRP-3, NOD-like receptor protein 3; NT-proBNP, N-terminal pro-brain natriuretic peptide; PAMPs, pathogen-associated molecular patterns; PLT, platelets; PMN, Polymorphonuclear cells; sCD40L, soluble CD40 ligand; TCR, T-cell receptor; TF, tissue factor; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α; VTE, Venous thromboembolism; vWF, von Willebrand factor. This figure has been created with BioRender.com.
First, WBC is a basic marker of peripheral blood morphology. Afzal et al. (1999) first described an independent elevation in WBC levels in subjects diagnosed with APE (Afzal et al. 1999; Trujillo-Santos et al. 2008). Leukocytosis is the classical marker of inflammation and is commonly used in clinical practice because of its low cost and easy accessibility. Leukocytosis has been shown to be related to the prognosis and severity of several cardiovascular diseases such as acute coronary syndromes (ACSs) and heart failure (Furman et al. 2004; Engström et al. 2009). Additionally, leukocytes contribute to increased local thrombogenic activity, and WBC levels are associated with factors VII and VIII and fibrinogen concentration (Bovill et al. 1996; Zarbock et al. 2007).
In the RIETE registry, WBC count has been shown to be related to VTE recurrence, major hemorrhage, and mortality in cancer patients (Trujillo-Santos et al. 2008). It has been demonstrated that an increased WBC count is more likely to be a prognostic tool than a diagnostic tool. Both low (<4 × 109/L) and high (>10 × 109/L) WBC levels are associated with higher 14-day mortality among patients with APE (Trujillo-Santos et al. 2008). Huang et al. (2011) found that WBC count >11,000/mm3 was an independent predictor of mortality among patients diagnosed with APE.
Among the WBC subtypes, neutrophils play a key role in modulating inflammatory responses in the atherosclerotic process (Afzal et al. 1999; Rosales 2018). The peripheral NLR has been proposed as a better indicator of inflammation than the WBC count (Zahorec 2001). According to the literature, the balance between neutrophils and lymphocytes is considered a systemic inflammation index (Jackson et al. 2019). An increased NLR has been shown to be related to coronary plaque disruption in ACS and higher in-hospital mortality. Neutrophils appear to modulate inflammatory responses during inflammatory processes (Falk et al. 1995).
In their retrospective study, Kayrak et al. (2014) first confirmed that NLR is an independent predictor of 30-day mortality, regardless of systolic blood pressure, creatine kinase MB (CK-MB), age, and diabetes mellitus type 2 among patients with APE. Previously, the prognostic value of NLR was confirmed in heart failure and ACS (Acanfora et al. 2001; Karabinos et al. 2009). Cytokines released from neutrophils may be responsible for the vascular injury. This clinical finding aligns with the results obtained in APE animal models, which showed neutrophil infiltration in the RV (Zagorski et al. 2003). A positive correlation between the CK-MB value, a marker of RV failure, and the neutrophil count was observed (Kayrak et al. 2014). Soylu et al. (2016) showed a significant correlation between a higher right-to-left ventricle (RV/LV) index, which is considered a negative prognostic marker in clinics, and the NLR. The higher NLR among APE patients and its positive predictive value for mortality have been confirmed in a series of further studies (Akgullu et al. 2015; Celik et al. 2015; Ma et al. 2016; Ates et al. 2017; Kurtipek et al. 2017; Galliazzo et al. 2018; Farah et al. 2020; Phan et al. 2020; Bi et al. 2021).
PLT play an important role in immunothrombosis (Balta et al. 2013). APE is associated with endothelial dysfunction and the release of CRP and pro-inflammatory cytokines (Bucek et al. 2002). As the thrombus develops, the arterial wall becomes damaged, leading to leukocyte influx and platelet aggregation. NLR and platelet to lymphocyte ratio are indicators of higher inflammation levels and predictors of disease severity and poor prognosis (Karatas et al. 2016).
CRP is a nonglycosylated protein synthesized in the liver during the acute phase and is induced by IL-6 downstream of IL-1β (Ridker 2016). The concentration of this nonspecific marker of inflammation increases dynamically during inflammatory diseases and infections (Clyne and Olshaker 1999). CRP is a well-known modulator of innate immunity stimulated mainly by IL-1, IL-6, and tumor necrosis factor (TNF) (Kolb-Bachofen 1991; Galeano-Valle et al. 2021). Peripheral blood monocytes stimulated by CRP can promote the release of TFs, contributing to thrombosis through the actions of IL-6 and monocyte chemoattractant protein-1 (MCP-1), which cause inflammation in the vascular wall (Figure 2).
Higher CRP levels in APE patients are associated with higher early (30 days) mortality, especially in patients diagnosed with infarction and pleural effusion (Buyuksirin et al. 2021). CRP test is an available and easy-access diagnostic parameter and it is routinely used in daily clinical practice. Under physiological conditions, the CRP levels were <1 mg/dL. However, during the acute phase, CRP levels increase rapidly within the first 6–8 h and reach their highest levels after 48 h (Mitchell et al. 2008; Galeano-Valle et al. 2021). There was a positive correlation between CRP levels and VTE among healthy subjects. When CRP rises from <1 mg/L to >3 mg/L, the risk of VTE doubles (Zacho et al. 2010) (Table 1).
Summary of the main studies in humans
| Marker | Author | Country | Year | Study design | Total patients | Main conclusion |
|---|---|---|---|---|---|---|
| Beckers et al. | Netherlands | 2010 | Prospective | 433 | The study demonstrated that four SNPs in inflammatory-related genes of IL-1α, IL-4, IL-6, IL-13 are related to VTE episodes. |
| IL-1 | Abuduhalike et al. | China | 2020 | Prospective | 284 | In patients with gene variant of IL-1 rs1800587 GG + GA are at higher risk of VTE than subjects with AA genotype (or 4.444, 95% CI = 1.466–13.470). |
| Zee et al. | USA | 2009 | Prospective | 22,413 | Gene polymorphism rs1143634 in the IL-1β gene was associated with a reduced risk of idiopathic VTE (hazard ratio = 0.59, 95% CI = 0.440.80, P = 0.0007, FDR = 0.02). Gene polymorphism rs1800872 in the IL-10 gene was associated with increased risk (hazard ratio = 1.42, 95% CI = 1.12–1.80, P = 0.004, FDR = 0.07). |
| NETs | Zabczyk et al. | Poland | 2020 | Prospective | 126 | Hypofibrinolysis is associated with enhanced NET formation and worst APE course. |
| IL-6 | Vormittag et al. | Austria | 2006a | Prospective | 355 | No relationship was found between IL-6 and its promoter polymorphism (–174) G > C and the risk of spontaneous VTE. |
| IL-4, IL-6, IL-8, IL-10, IL-1β, MCP-1, EGF | Bontekoe et al. | USA | 2021 | Prospective study | 157 | Cytokines IL-4, IL-6, IL-8, IL-10, IL-1β, MCP-1, and EGF demonstrated varying levels of significant increase (P < 0.05) in massive-risk PA patients compared to submassive- and low-risk PA patients. |
| CRP | Vormittag et al. | Austria | 2006b | Prospective | 355 | CRP + 1444TT genotype was significantly associated with increased CRP plasma levels in healthy individuals. CRP + 1444TT was more frequent (14%) among controls than DVT patients (9%, P = 0.26) or PE PA patients (6%, P = 0.05). |
| Stewart et al. | USA | 2017 | Prospective, double-blinded, randomized controlled trial, with 3-month follow-up | 60 | A significant reduction of IL-6, CRP, and MPO concentration was observed in the population of patients with APE after 3-month follow-up as compared to baseline (in group treated with Tenecteplase as well as placebo). | |
| Matos et al. | Brazil | 2011 | Prospective | 245 | VTE was related to higher IL-6 level and it remained significant independently of hs-CRP concentration and BMI. |
| EGF | Dirican et al. | Turkey | 2016 | Prospective | 57 | Higher SCUBE1 plasma concentration was observed in the APE than in the non-APE and controls. |
| EGF | Alqudah et al. | Jordan | 2017 | Prospective | 75 | Patients with VTE presented significantly lower EGF concentration in comparison with healthy subjects. |
| TNF-α | Mazetto et al. | Brazil | Human | Prospective | 2012 | Circulatory levels of TNF-α were significantly increased in patients with VTE compared to healthy controls. |
| TGF-β | Memon et al. | Sweden | 2014 | Prospective with 39 months follow-up | 126 | Reduced plasma concentration of TGF-β1 and TGF-β2 in patients with recurrent VTE. |
| Wang et al. | Sweden | 2019 | Prospective | 78 | 12 mRNAs: (miR-15b-5p, miR106a-5p, miR-197-3p, miR-652-3p, miR-361-5p, miR-222-3p, miR-26b-5p, miR-532-5p, miR-27b-3p, miR-21-5p, miR-103a-3p, and miR-30c-5p) were related to higher VTE reoccurrence. Eight miRNAs significantly correlated with circulating TGFβ1/2. | |
| sCD40L | Kaya et al. | Turkey | 2012 | Prospective | 84 | sCD40L concentration was significantly higher among APE patients in compare with healthy controls. |
| GDF | Lankeit et al. | Germany | 2008 | Prospective | 101 | GDF-15 is an independent predictor of a complicated 30-day outcome after APE. |
| Skowronska et al. | Poland | 2020 | Prospective | Plasma GDF‐15 concentration is a valuable biomarker in prediction of bleeding and hemodynamic destabilization in APE patients. |
APE, acute pulmonary embolism; BMI, body mass index; CI, confidence interval; CRP, C-reactive protein; DVT, deep vein thrombosis; EGF, epidermal growth factor; FDR, false discovery rate; GDF-15, growth differentiation factor-15; hs-CRP, high sensitive CRP; IL, interleukin; MCP-1, monocyte chemoattractant protein-1; miRNAs, micro-RNAs; MPO, myeloperoxidase; mRNA, messenger RNA; NETs, neutrophil extracellular traps; PA, pulmonary artery; PE, pulmonary embolism; sCD40L, soluble CD40 ligand; SNP, single nucleotide polymorphisms; TGF, transforming growth factor; TGF-β, transforming growth factor-β; TNF, tumor necrosis factor; VTE, venous thromboembolism.
CRP exerts prothrombotic effects, and chronically elevated CRP levels are associated with a higher risk of VTE among patients diagnosed with advanced cell lung cancer and among the obese population (Horvei et al. 2016; Shen et al. 2017). Moreover, regardless of the primary cause of hospitalization, CRP concentration correlates with VTE risk within a 90-day period (Grimnes et al. 2018). During the COVID-19 pandemic, elevated CRP levels at hospital admission were associated with higher in-hospital mortality and postdischarge VTE (Li et al. 2021; Smilowitz et al. 2021; Machowski et al. 2024). Prospective population-based studies demonstrated an almost 23% increase in VTE episodes per 5 mg/L increase in CRP level (Kunutsor et al. 2017).
A study conducted among 276 patients diagnosed with APE demonstrated that CRP values >5 mg/L on admission were associated with a higher 30-day bleeding risk and 30-day mortality (Demelo-Rodriguez et al. 2020). Another study conducted by Marchena Yglesias et al. (2006) indicated the potential role of CRP in predicting VTE complications and mortality during a 1-year follow-up.
As previously mentioned (Imiela et al. 2024), there is a link between RV dysfunction and inflammatory cell influx in experimental models of APE (Iwadate et al. 2003; Zagorski et al. 2007; Imiela et al. 2024). Acute RV pressure overload resulting in RV failure is one of the most severe consequences of APE and is related to a higher rate of mortality (Konstantinides et al. 2020).
A study by Abul et al. (2011) showed a positive correlation between mortality and CRP levels among patients with APE characterized by RV dysfunction. In another study by Najarro et al. (2024), conducted on 633 patients diagnosed with stable APE, elevated CRP levels were associated with significantly higher APE-related mortality and RV dysfunction. However, there was no correlation with 30-day all-cause mortality (Najarro et al. 2024) (Table 1). Moreover, a meta-analysis by Ding et al. (2023) demonstrated not only the relationship between elevated CRP and higher VTE risk but also suggested that CRP might act as a marker for pending thrombosis. Among patients with cancer, a CRP concentration >4.5 mg/L after withdrawal of anticoagulation therapy predicted a 10-fold increase in VTE recurrence (Jara-Palomares et al. 2018).
In contrast, other studies have reported different results regarding the predictive value of VTE. First, no significant correlations were found between a higher risk of VTE and CRP gene polymorphisms, resulting in higher CRP concentrations or platelet activation (Vormittag et al. 2006a; Chung et al. 2007). Additionally, no correlation was found between CRP levels and mortality (Hogg et al. 2013). Similarly, no significant correlation was found between RV dysfunction or mortality rate and CRP concentration among patients with a hemodynamically stable course of APE over 6 months of observation (Kline et al. 2008).
Although preclinical studies have indicated the role of inflammation in APE pathogenesis, primarily due to the development of RV dysfunction (Dewachter and Dewachter 2018), slightly elevated CRP or WBC plasma concentrations might mimic the biochemical state of ongoing infection in clinical practice. Procalcitonin (PCT) is another well-known marker of systemic inflammation that predicts outcomes in patients diagnosed with pulmonary infection. However, there is little data regarding the role of PCT in APE.
PCT is a peptide precursor of the hormone calcitonin, which is involved in calcium homeostasis (Vijayan et al. 2017). Under physiological conditions, PCT is produced by C-cells of the thyroid gland from the CALC-1 gene located on chromosome 11 (Vijayan et al. 2017). Normally, PCT concentrations do not exceed 0.05 ng/mL. However, in the systemic inflammatory state, PCT is produced in response to higher levels of TNF-α and IL-6 or directly due to lipopolysaccharide (LPS) stimulation (Vijayan et al. 2017). In daily practice, PCT is a crucial marker for sepsis diagnosis, enabling distinction between viral and bacterial infections.
As previously mentioned, biochemical changes in inflammatory biomarkers may suggest ongoing pulmonary infection. Additionally, APE diagnosis might be complicated by coexisting clinical symptoms, such as fever and chest pain, which can lead to misdiagnosis. One severe consequence of APE is pneumonia related to APE-induced pulmonary infarction; however, data on this clinical phenomenon remain limited (Kaptein et al. 2021; Eggers et al. 2023). Pulmonary infarction is potentially life-threatening due to distal pulmonic artery occlusion, leading to lung parenchymal necrosis (Kaptein et al. 2021). In the clinical setting, the prevalence of APE-induced infarction ranges from 10 up to 50% in all patients diagnosed with APE (Cha et al. 2012). A prospective study conducted by Eggers et al. (2023) in 749 patients with APE demonstrated that clinical infections during APE were common; however, only very high CRP plasma concentrations >124 mg/L showed 90% specificity for in-hospital adverse outcome prediction.
Although elevated PCT levels on admission were not frequently discovered, a higher PCT concentration (>0.25 mg/L) effectively predicted study outcomes such as hemodynamic insufficiency, defined as the necessity of catecholamine usage, cardiopulmonary resuscitation, and in-hospital all-cause mortality (Eggers et al. 2023). Additionally, significant infections treated with antibiotics were observed in 50% of all APE patients, exerting a similar impact as an increase in one risk class of the ESC risk-stratification algorithm (Eggers et al. 2023).
To summarize, elevated plasma concentrations of PCT and CRP are independently associated with higher risk of adverse outcomes. Numerous clinical studies have been conducted to elucidate the connection between VTE and CRP. Despite some conflicting results, the vast majority of data demonstrate that CRP could act as a potentially valuable biomarker, though more data is needed to confirm these findings. Furthermore, modest leukocytosis might be routinely observed during APE episodes. However, it should not delay proper APE diagnostics and treatment, nor should it prompt the rapid enrollment of antibiotics without thorough clinical evaluation and infection exclusion.
Cytokines are major components of the immune system and are the main players in the immune response. Until recently, the relationship between immunity and APE was not well understood. Modern technologies have demonstrated that various cytokines are involved in the initiation and progression of APE. Bontekoe et al. (2021) used biochip-array technology to quantify inflammatory biomarkers in patients with APE and compared them with healthy controls. Biochip technology is a high-throughput method for analyzing blood plasma samples, which enables the simultaneous measurement of plethora of biomarkers for potential risk stratification. In this study, cytokines such as IL-6, IL-8, and epidermal growth factor (EGF) showed the greatest increase among patients with APE compared to healthy controls (Bontekoe et al. 2021).
IL-1β is a key cytokine with a variety of functions (Sun et al. 2011). The IL-1 family consists of 11 members, with IL-1α and IL-1β being the most important members.
IL-1β is produced by activated macrophages, monocytes, lymphocytes, endothelial cells, smooth muscle cells, and keratinocytes. Binding to the IL-1β receptor activates kinases and leads to the transcription of proinflammatory pathways via the nuclear factor (NF)-κB signaling pathway (Dunne and O’Neill 2003). In addition, it triggers the transcription and translation of IL-1α and IL-1β precursors as well as inflammasome components (Swanson et al. 2019; Abbate et al. 2020) (Table 1).
Inflammasomes are multiprotein complexes in immune cells that activate inflammatory responses, leading to pyroptosis, a form of cell death that contributes to TF release and enhances the prothrombotic environment. The inflammasome requires two signals for full activation (Potere et al. 2023).
First, danger-associated molecular patterns (DAMPs), such as adenosine triphosphate (ATP), IL-1β, and IL-pro-1a, initiate cellular priming by activating Toll-like receptors and IL-1 receptors. This process leads to the translocation of NF-κB to the cell nucleus, where it promotes the transcription of proinflammatory molecules.
Next, additional DAMPs, including cathepsin B, reactive oxygen species, mitochondrial proteins, and extracellular ATP, further promote inflammasome activation through the purinergic P2 × 7R receptor. The resulting K+ efflux activates NLRP3, leading to its oligomerization and binding to procaspase-1 and the apoptosis-associated speck-like protein containing a caspase recruitment domain. This interaction triggers the autocatalytic conversion of procaspase-1 into its active form, caspase-1, which then cleaves IL-1β, IL-18, and gasdermin into their active forms.
During gasdermin activation, N-terminal fragments form pores in the cell membrane, causing the robust release of IL-1β and IL-18, which further promote pyroptotic cell death. IL-1β also facilitates leukocyte adhesion by upregulating endothelial adhesion molecule 1 and platelet-derived growth factor, contributing to an exaggerated inflammatory response.
Neutrophils, macrophages, and PLT require minimal priming only for robust cytokine release and inflammasome activation (Potere et al. 2023). Importantly, IL-1α derived from the damaged endothelium can promote the early recruitment and activation of macrophages, PLT, and neutrophils at the site of thrombus formation. Both IL-1β and IL-18, released by PLT and leukocytes, contribute to the expression of endothelial adhesion molecules, the release of additional proinflammatory mediators, and increased vascular wall permeability (Rider et al. 2011; Potere et al. 2023).
Venous thrombi are predominantly formed by erythrocytes, fibrin, and PLT (von Bruhl et al. 2012). Traditionally, neutrophils express TF, which can bind to injured endothelium and PLT, promoting platelet aggregation and thrombus formation (Figure 2) (Darbousset et al. 2012; Potere et al. 2023). Moreover, neutrophils generate reactive oxygen species and NETs (Stark and Massberg 2021; Natorska et al. 2023). Recently, it has become evident that NET formation is a crucial defense mechanism during immune response, involving the release of DNA in a web-like structure consisting of proteins, DNA core, and proteases (Schulz et al. 2013; Potere et al. 2023).
NETs can capture and present factor XII, promote thrombogenesis, and contribute to exaggerated fibrin and thrombin generation (Stark and Massberg 2021). NET DNA interacts with fibrin and vWF, promoting the formation of a stable thrombus (Stark and Massberg 2021). NETs and inflammasome activation are mutually related. Activation of the NLRP3 inflammasome is essential for NETs formation, and NETs further promote NLRP3 inflammasome activation, resulting in IL-1β and IL-18 release, contributing to exaggerated thromboinflammation (Kahlenberg et al. 2013; Munzer et al. 2021; Potere et al. 2023). Additionally, PLT exhibit NLRP3 inflammasome upon activation and produce IL-1α, IL-1β, and IL-18 (Figure 2) (Murthy et al. 2017). In humans exaggerated NET formation is related to hypofibrinolysis and the worst APE prognosis (Zabczyk et al. 2020).
A correlation study in a Chinese population indicated a relationship between the TT genotype of IL-1 rs2234650 and VTE occurrence (Beckers et al. 2010). Additionally, higher circulating levels of IL-1 were observed in the VTE group than in controls (Abuduhalike et al. 2020). Other studies in white populations, such as Women's Genome White Study and the Dutch population, showed an association between gene polymorphisms rs1143634 and rs1800587 and VTE (van Aken et al. 2000; Zee et al. 2009). A follow-up study performed by the same group of researchers evaluated the relationship between long-term (5 years) outcomes of VTE patients and mutations in inflammatory genes (Abuduhalike et al. 2020). Patients with a higher simplified PESI score and carriers of rs1800587 mutation genotype were characterized by higher mortality (Abuduhalike et al. 2020). The analysis of single nucleotide polymorphisms (SNP) is a relatively cost-effective but valuable method for analyzing the genetic background of VTE phenotypes. It can be postulated that there is a close link between VTE prognosis and inflammatory gene polymorphism.
Generally, there is a growing body of evidence demonstrating the key role of NETs in immunothrombosis. However, the vast majority of studies are conducted in experimental models of atherosclerosis and thrombosis. While a precise description of the exact role of NETs and inflammasome activation is beyond this review, it is essential to highlight the mechanism leading to IL-1 activation. In humans, NETosis is implicated in patients diagnosed with cancer-induced VTE (Schulz et al. 2013). Inflammation significantly contributes to VTE, with NLRP3 inflammasome and IL-1 being key factors. The NLRP3 inflammasome is a central player in thromboinflammation, promoting the release of IL-1β and IL-18, leading to VTE. Nonselective NLRP3 inhibitors, such as colchicine and IL-1 blockers like canakinumab show promise in reducing VTE incidence. Both preclinical and clinical studies support the effectiveness of inflammasome pathway blockade in managing VTE. This suggests promising therapeutic roles for colchicine and canakinumab in reducing VTE through inflammasome inhibition. Various studies have shown reduced thrombogenesis with inflammasome pathway inhibitors.
However, some clinical trials and studies have shown inconsistent results, with traditional anticoagulants like heparins and DOACs remaining more reliable for VTE management. Additionally, the cost and accessibility of novel inflammasome inhibitors may limit their practical application. It should be emphasized that there is a need for a more nuanced understanding of VTE pathogenesis and therapeutic approaches, considering both inflammation and other contributing factors.
IL-6 is a four-helix bundle cytokine that plays numerous roles, including coordinating innate and adaptive immune responses and promoting immune cell proliferation and differentiation. IL-6 is produced by activated monocytes and macrophages, endothelial cells, and T and B lymphocytes (Figure 2) (Najem et al. 2020). IL-6 is a major cytokine associated with DVT progression and postthrombotic syndrome development (Kerr et al. 2001). IL-6 function is closely related to endothelial cell and tissue damage, platelet activation, and the promotion of coagulation (Zhang et al. 2020). Elevated IL-6 concentrations in patients with APE contribute to thrombosis development by enhancing platelet responsiveness to thrombin (Du and Tan 2014). Notably, higher IL-6 concentrations are observed in patients with massive APE than in those with low and submassive APE (Bontekoe et al. 2021) (Table 1).
In a multicenter, double-blinded, randomized controlled trial performed by Stewart et al. (2017) involving 60 patients with submassive APE, a significant but temporary systemic inflammation was observed. Patients were treated with standard anticoagulation and divided into placebo and tenecteplase subgroups. Blood sample analyses at the start and after 3 months of follow-up showed a significant reduction in IL-6 concentration in both subgroups (Stewart et al. 2017).
These findings suggest that IL-6 may serve as a valuable marker for stratifying APE severity. However, studies by Vormittag et al. (2006b) found no relationship between the IL-6 promoter -174 G > C polymorphism and VTE occurrence. In contrast, a case-control study performed by Matos et al. (2011) indicated that higher IL-6 levels were significantly associated with VTE independent of high sensitive CRP (hs-CRP) concentration and body mass index (BMI).
IL-8, known as CXCL-8, is a chemokine responsible for the recruitment of monocytes and neutrophils (Figure 2). Monocytes and macrophages are the primary cellular sources of IL-8 (Apostolakis et al. 2009). IL-8 promotes procoagulant activity by enhancing integrin expression on the cell surface, thereby playing a critical role in the immune response and inflammation (Luster 1998). Data coming from previous studies have shown that elevated IL-8 levels correlate with the first thrombotic event. Bontekoe et al. (2021) demonstrated that patients with massive APE are characterized by higher IL-8 levels than patients with “submassive” APE risk. IL-8 elevation may be a potential indicator of thrombosis. Higher IL-8 concentrations were in line with data from animal models showing the key role of neutrophils in the APE-induced inflammatory state (Table 1).
EGF is a prominent protein released from activated PLT during thrombosis formation (Dirican et al. 2016). Bontekoe et al. (2021) indicated that EGF levels were higher in patients with APE compared to the healthy controls. However, this elevation did not show significant differences in APE risk classification. Previous studies have also shown that the concentration of EGF particles, such as SCUBE-1, is higher in APE patients than in non-PE patients and control subjects (Dirican et al. 2016). Alqudah et al. (2017) evaluated the potential role of EGF in VTE and found notably lower EGF concentrations in patients with VTE. The study revealed that the form of VTE-DVT did not affect the EGF levels. The decrease in EGF concentration might be related to the increased consumption of growth factors during vessel wall healing and recovery (Alqudah et al. 2017) (Table 1).
TNF-α is a key regulator of the immune response and plays a crucial role in orchestrating the cytokine network. TNF-α is primarily produced by T lymphocytes and macrophages (Figure 2) (Balkwill 2009). It exists in two forms: a soluble cytokine generated after cleavage by a disintegrin and A metalloproteinase 17, or a transmembrane protein. Two kinds of TNF-α receptors can be distinguished: TNFR1, which is expressed in the majority of cell types, and TNFR2, whose expression is preserved in hematopoietic cells. NF-κB is the main signaling pathway activated by TNF-α receptor stimulation. Studies have shown that circulatory levels of TNF-α are significantly higher in patients with VTE than in healthy controls (Mazetto et al. 2012). According to the literature, TNF-α may facilitate thrombus resolution by promoting collagen lysis, fibrinogen lysis, and neovascularization (Najem et al. 2020) (Table 1).
TGF-β is a potent pro-fibrotic factor that promotes organ sclerosis and inhibits the growth of lymphocytes and endothelial cells (Najem et al. 2020). In humans, TGF-β exists in three subtypes: TGF-β1, TGF-β2, and TGF-β3, and exerts anti-inflammatory effects in the vessel wall. TGF-β1 is the most crucial isoform of the cardiovascular system (Annes et al. 2003) (Table 1). It possesses antiatherogenic properties in atherosclerosis and antifibrinolytic features in hemostasis. Data obtained from patients with a first episode of unprovoked VTE enrolled in a prospective population-based study—Malmo Thrombophilia Study (MATS), confirmed an association between higher risks of recurrent VTE and lower levels of TGF-β1 and TGF-β2 (Memon et al. 2014). Furthermore, a continuation of MATS study demonstrated a positive correlation between certain micro-RNAs (miRNAs) and TGF-β1 expression. miRNAs are short, noncoding, single-stranded RNAs that inhibit translation and gene expression by promoting messenger RNA (mRNA) degradation. miRNAs regulate almost 90% of all human genes and play important roles in maintaining homeostasis. mRNA can be secreted from various cells into the bloodstream and is either associated with proteins or exosomes (Figure 2). According to the literature, miRNAs are valuable biomarkers for conditions such as heart failure, diabetes mellitus, and cancer. Specifically, miR-15b-5p, miR-106a-5p, miR-197-3p, miR-652-3p, miR-361-5p, miR-27b-3p, and miR-103a-3p are positively correlated with TGF-β1 expression (Wang et al. 2019).
Patients with a first episode of unprovoked VTE were characterized by significantly lower TGF-β concentrations, indicating that TGF-β may serve as a useful predictive marker for recurrence in these patients. Additionally, 12 plasma miRNAs have been identified as noninvasive and suitable biomarkers for predicting VTE recurrence.
The CD40 ligand (CD40L) is a transmembrane protein that is structurally related to the TNF-α family and is primarily found on the surface of T CD4+ T cells (Henn et al. 1998). Henn et al. (1998) demonstrated that the CD40L molecule is also present on the surface of PLT (Figure 2). It is well known that the CD40-CD40L interaction plays an important role in the pathogenesis of cardiovascular diseases, such as atherosclerosis, coronary artery disease (Garlichs et al. 2001), atrial fibrillation (Duygu et al. 2008), and peripheral artery disease (Blann et al. 2005) (Table 1).
CD40L has also been studied in patients with primary and secondary pulmonary arterial hypertension (PAH) (Damas et al. 2004). In a brilliant study, Damas et al. (2004) reported a higher CD40L concentration among subjects diagnosed with primary and secondary PAH. In vitro test showed that recombinant soluble CD40L (sCD40L) induced MCP-1 and IL-8 gene expression in endothelial cells. Plasma levels of these chemokines were elevated in all PAH groups and were significantly correlated with sCD40L levels and hemodynamic parameters (Damas et al. 2004).
Circulating CD40L molecules are predominantly produced by PLT and reflect their activity. Moreover, Damas et al. (2004) found increased plasma levels of CD40L in patients with PAH compared with healthy controls. Importantly, sCD40L concentrations are significantly higher in patients diagnosed with APE than in healthy controls (Kaya et al. 2012). However, RV dysfunction parameters were not associated with higher sCD40L levels (Kaya et al. 2012).
GDF-15 is a well-known member of the TGF-β cytokine superfamily and it plays a crucial role in cell growth, inflammation, and apoptosis (Paralkar et al. 1998). GDF-15 is a chemokine produced by macrophages upon activation and oxidative stress (Bootcov et al. 1997). Under physiological conditions, GDF production is low; however, it increases significantly during inflammation (Skowronska et al. 2020). GDF-15 has been identified as a predictor of cardiovascular death and mortality independent of CRP, cardiac troponin T, and NT-proBNP (Sharma et al. 2017). GDF-15 is produced in response to TNF-α and IL-1β during inflammatory states and tissue injury as well as in macrophages, smooth muscle cells, and endothelial cells (Figure 2) (Bermudez et al. 2008) (Table 1).
The influence of GDF-15 on thrombin/antithrombin and platelet activities was investigated. Rossaint et al. (2013) found that GDF-15 reduced platelet aggregation caused by thrombin, adenosine diphosphate (ADP), and thromboxane A2 analogs due to integrin inhibition in both mouse models and humans. However, another study showed contradictory findings regarding the association between GDF-15 and platelet activity (Lippi et al. 2017). In experimental models, GDF-15 knockout (GDF-15–/–) mice presented accelerated thrombus formation, enhanced neutrophil recruitment, and increased platelet aggregation (Kempf et al. 2011).
The role of GDF-15 in the APE has been extensively evaluated. Lankeit et al. (2008) conducted a prospective study of 123 consecutive patients with APE. The study found that those who experienced APE-related complications within the first 30 days had higher baseline GDF-15 levels (Lankeit et al. 2008). Additionally, a study by Duran et al. (2014) indicated that high serum GDF-15 could be more effective in mortality prediction than NT-proBNP in the population of normotensive subjects diagnosed with APE.
The predictive role of GFD-15 in DVT was examined. Liang et al. (2020) demonstrated that plasma GDF-15 concentrations were significantly higher in patients with DVT than in healthy controls. Furthermore, a significant positive correlation was found between GDF-15 levels and thrombus burden, as measured by the number of venous segments with thrombi. Patients with higher GDF-15 concentrations and more severe vein lesions are characterized by higher Geneva and Wells-APE scores (Liang et al. 2020). In vitro studies have shown that GDF-15 reduces platelet aggregation in a dose-dependent manner. Skowronska et al. (2020) performed a study focusing on the value of GDF-15 in predicting bleeding and serious adverse outcomes (death, need for catecholamine administration, reperfusion therapy, and cardiopulmonary resuscitation). The study concluded that higher GDF-15 concentrations were associated with increased occurrence of bleeding and serious adverse events (SAE) defined as death, need for cardiopulmonary resuscitation, need for urgent reperfusion therapy, need for catecholamine administration (Skowronska et al. 2020). A GDF-15 concentration >1680 ng/L was considered a threshold for predicting bleeding and SAE (Skowronska et al. 2020). In summary, GDF-15 is a promising marker for predicting hemodynamic destabilization and bleeding complications in APE subjects.
It is common knowledge that coagulation cascade consists of two major pathways: intrinsic and extrinsic. The extrinsic pathway initiates with the formation of factor VII and TF on the surface of monocytes or endothelial cells (Marcos-Jubilar et al. 2023). TF, a crucial glycoprotein, is expressed by neutrophils and peripheral blood mononuclear cells and is activated in response to tissue injury or infection (Ryan et al. 2022; Marcos-Jubilar et al. 2023). Once TF binds with factor VII, it activates factor VIIa, which triggers the extrinsic pathway. During infection, bacterial LPSs, also known as pathogen-associated molecular patterns (PAMPs), are presented to various cells, including endothelial cells, monocytes, and epithelial cells (Jackson et al. 2019). Upon PAMP presentation, NF-κB transcription leads to the expression of TF mRNA, which triggers changes in cell membrane conformation and the decryption of TF. This decryption is essential for TF's activation as a cofactor, enabling it to bind with factors X and VIIa. Pyroptosis, a form of programmed cell death, induces cell membrane injury, leading to the activation of the caspase pathway and inflammasome formation (as described in Section 3.1.). The binding of TF and activation of factor VIIa subsequently trigger the intrinsic coagulation pathway, resulting in the conversion of factor X to Xa and IX to IXa. Thrombin, generated in this process, converts fibrinogen to fibrin and is also responsible for platelet activation via PARs. Activated PLT and neutrophils contribute to the release of NETs through the process of NETosis (McDonald et al. 2017). These NETs exacerbate thrombosis by inducing endothelial dysfunction and further activating the intrinsic pathway. Additionally, activated PLT lead to TF alternative pathway activation (Jayarangaiah et al. 2020). Overall, TF plays a pivotal role in orchestrating the inflammatory response during the early stages of thrombus formation (Jayarangaiah et al. 2020).
vWF is an important glycoprotein which takes part in hemostasis and it is usually released by PLT and endothelium cells (Brehm 2017; Marcos-Jubilar et al. 2023). vWF is also responsible for leukocytes influx and it interacts with NETs in thrombosis process (Staessens et al. 2020). Animal studies showed that NETs are able to increase the levels of IL-6 and decrease a disintegrin and metalloproteinase with a thrombospondin type 1 motif, member 13 (ADAMTS13) activity. ADAMTS13 is a metalloproteinase which function is to regulate the size and activity of vWF by cleaving ultra-large vWF into smaller parts. In summary, vWF interaction with NETs leads to PLT and leukocytes recruitment, exaggerating immunothrombosis during pathological conditions such as COVID-19 illness, stroke, or angiopathy.
In daily clinical practice, DOACs or heparin are fundamental treatments for patients diagnosed with VTE. These medications are effective in resolving thrombi; however, their use is associated with an increased risk of hemorrhagic complications because fibrin, a crucial element of physiological hemostasis, is affected (Mackman et al. 2020). The primary mechanism of action for anticoagulants is the reduction of coagulation factor activity (Ryan and O’Neill 2022; Marcos-Jubilar et al. 2023). In our previous work, we discussed the potential anti-inflammatory effects of vitamin K antagonists and DOACs. Here, we will briefly outline selected therapeutic options for immunothrombosis, which have mainly been studied in preclinical models.
The vast majority of therapeutic strategies for VTE focus on targeting the coagulation cascade. In clinical scenarios, the main disadvantage is the risk of severe side effects, such as life-threatening hemorrhages, particularly in the central nervous system. Although anticoagulants are effective in preventing VTE recurrence (Di Nisio et al. 2016; Potere et al. 2023), they are not without risks. The anti-inflammatory effects of anticoagulants and statins have been previously described (Imiela et al. 2024).
New therapeutic strategies aim to target molecular pathways involved in immunothrombosis, such as by inhibiting inflammasomes, NETs, TF, and vWF, with the goal of reducing thrombotic risks without increasing bleeding complications. However, most of the available data come from animal studies, and large clinical trials are needed to confirm the safety and efficacy of these new drugs.
First, it can be postulated that inhibiting the inflammasome and pyroptosis might help bridge the gap in translating thromboinflammation research into clinical practice. Selective NLRP3 inflammasome inhibitors, such as dapansutril, have been studied in the treatment of systolic heart failure and gout (Kluck et al. 2020; Wohlford et al. 2020). Some nonspecific inflammasome inhibitors, like colchicine—which blocks microtubule polymerization and influences neutrophil degranulation and diapedesis—can also affect thrombosis. In an experimental model of VTE, the administration of colchicine reduces clot size and significantly decreases the required dose of heparin (by up to 20-fold) needed to prevent thrombosis, without affecting tail vein bleeding time (Abu-Fanne et al. 2019).
In clinical trials, the use of colchicine for atherosclerosis has shown promising results. The Low Dose Colchicine-2 Trial (LoDoCo) demonstrated that 0.5 mg colchicine once daily reduced the primary endpoint of cardiovascular death, myocardial infarction, and ischemic stroke in patients with chronic coronary artery disease by 31% (0.57–0.83; P < 0.001) (Nidorf et al. 2020). The LoDoCo proteomic substudy indicated that low-dose colchicine treatment reduced the concentrations of IL-6, IL-18, and IL-1RA, as well as neutrophil degranulation and inflammasome activation, suggesting a pleiotropic mechanism of action (Potere et al. 2023). The Colchicine Cardiovascular Outcomes Trial reported that low-dose colchicine reduced coronary and cerebral thrombotic events in patients after myocardial infarction (Tardif et al. 2019). As mentioned previously, anti-IL-1 monoclonal antibody exerted anti-inflammatory effects which was proved in an elegant CANTOS trial (Ridker et al. 2017; Imiela et al. 2024).
Lesser-known molecules are also capable of blocking inflammasome activation. For example, indole ureas, nitrofurans, and acrylamides inhibit TF activation in the endothelium during SARS-CoV-2 infection (Tardif et al. 2019). Another potential molecule, dimethyl fumarate, has shown effectiveness in sepsis treatment by inhibiting gasdermin and attenuating pyroptosis (Humphries et al. 2020). In a rat sepsis model, NLRP3 inhibition with MCC950 reduced platelet activation (Coll et al. 2019). Additionally, high-mobility group box 1 (HMGB1) has been shown to attenuate inflammatory processes during sepsis and disseminated intravascular coagulation (DIC) in experimental models (Yang et al. 2020).
Finally, HMGB1 protein inhibitors, such as metformin and thrombomodulin, have shown potential in preclinical studies for controlling inflammation and preventing conditions like DIC.
Next, Janus kinase inhibitors like baricitinib, ruxolitinib, and tofacitinib have been identified as potential therapeutic agents for reducing immunothrombosis by modulating the inflammatory pathways involved in arthritis and myeloproliferative neoplasms (Baldini et al. 2021).
PAD4 is essential in the formation of NETs. PAD4 inhibitors, such as GSK199 and GSK484, have shown promise in preclinical trials by specifically inhibiting PAD4, thereby reducing NET formation and the associated thrombotic risks (Lewis et al. 2015). It can be hypothesized that PAD4 inhibitors derived from benzol-arginine might also be effective (Lewis et al. 2015; Huang et al. 2023; Marcos-Jubilar et al. 2023). In addition, DNase enzymes, particularly DNase 1, break down the DNA in NETs, which can reduce thrombus formation. Literature indicates that DNase 1 is already used as a treatment for conditions like cystic fibrosis. The administration of DNases is also being explored for its potential in treating thrombotic conditions, particularly after thrombectomy procedures (Van Bruggen and Martinod 2023).
The complement system, part of the innate immune response, can contribute to thrombotic complications in infections. Eculizumab, a monoclonal antibody against complement component C5, has been used to treat thrombotic microangiopathy and is being explored as a potential treatment for COVID-19-related systemic thrombosis (Campbell and Kahwash 2020; Marcos-Jubilar et al. 2023).
Emerging anti-inflammatory therapies aimed at modulating the inflammatory response in acute APE are crucial for the future management of patients diagnosed with VTE. Drugs targeting the NLRP3 inflammasome, such as colchicine and canakinumab, show promise as future treatment strategies.
However, there is still a paucity of data from clinical trials on the potential of targeting the inflammasome and IL-1 signaling pathways as therapeutic strategies in VTE. This approach could complement traditional anticoagulation therapies, potentially reducing the risk of VTE recurrence and long-term complications like postthrombotic syndrome. Given the interconnected nature of inflammation and coagulation—particularly through the NLRP3 inflammasome, which is emerging as a crucial player in thromboinflammation—therapeutic strategies that target these inflammatory pathways could offer benefits beyond conventional anticoagulation. By addressing the underlying inflammatory mechanisms contributing to VTE, such strategies may reduce the risk of serious hemorrhages and improve overall outcomes.
Our review explores the intricate relationship between thrombosis, inflammation, and the immune system, with a particular focus on the concept of immunothrombosis, which describes how the immune response can influence thrombus formation. We emphasize the role of NETs in coagulation and the various mechanisms linking inflammation and thrombosis, such as cytokines, complement activation, and inflammasome formation.
The NLRP3 inflammasome plays a critical role at the intersection of inflammation and coagulation. Targeting this pathway offers a promising therapeutic strategy to address the inflammatory components of thrombotic diseases. This approach could potentially reduce the incidence and recurrence of conditions like VTE while minimizing the risks associated with traditional anticoagulant therapies.
It is crucial for clinicians managing VTE to understand these mechanisms, as novel anti-inflammatory drugs might enhance thrombus clearance, reduce vein scarring, and improve overall vascular function postthrombosis. This could protect patients from long-term complications and recurrent thromboembolism. However, more translational studies from bench to bedside are needed to confirm these hypotheses.