
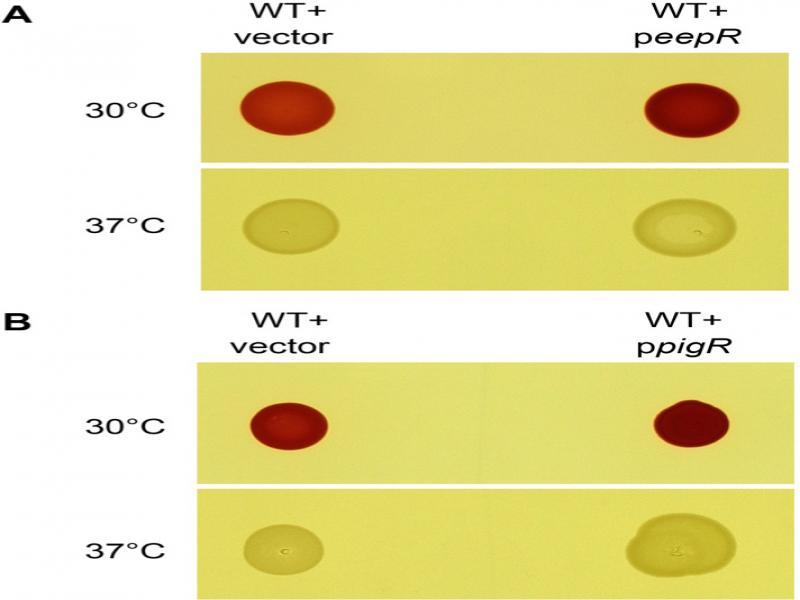
Fig. 1.
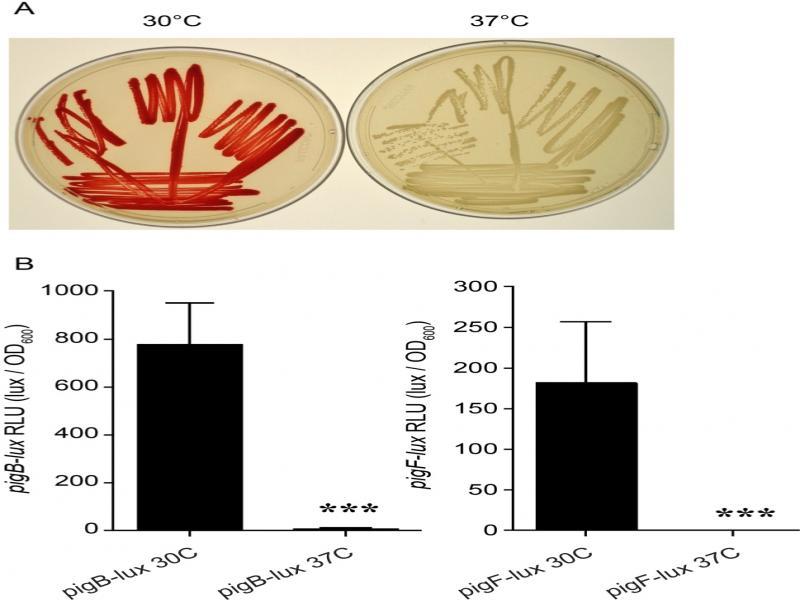
Fig. 2.
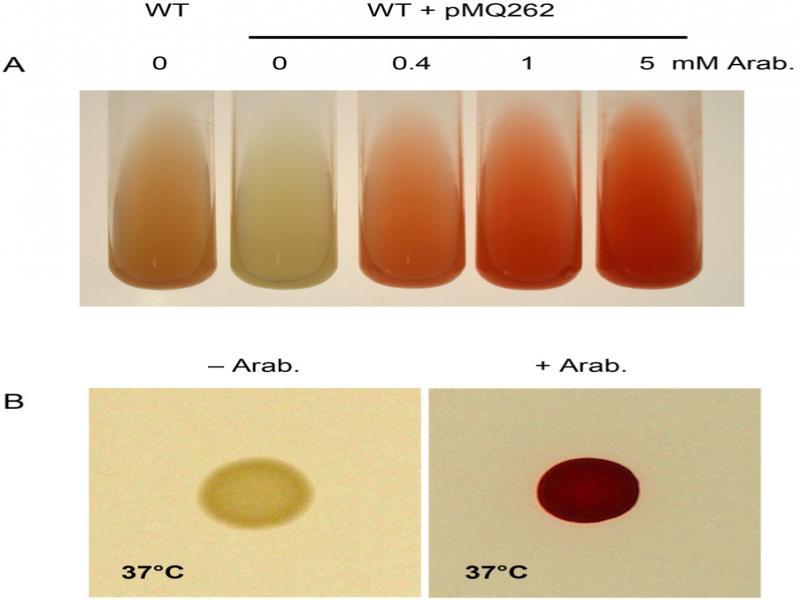
Fig. 3.
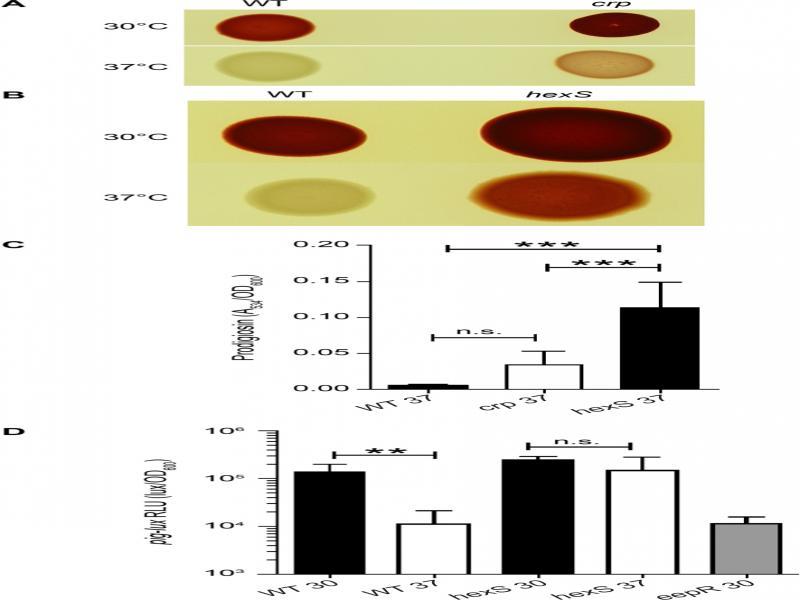
Fig. 4.
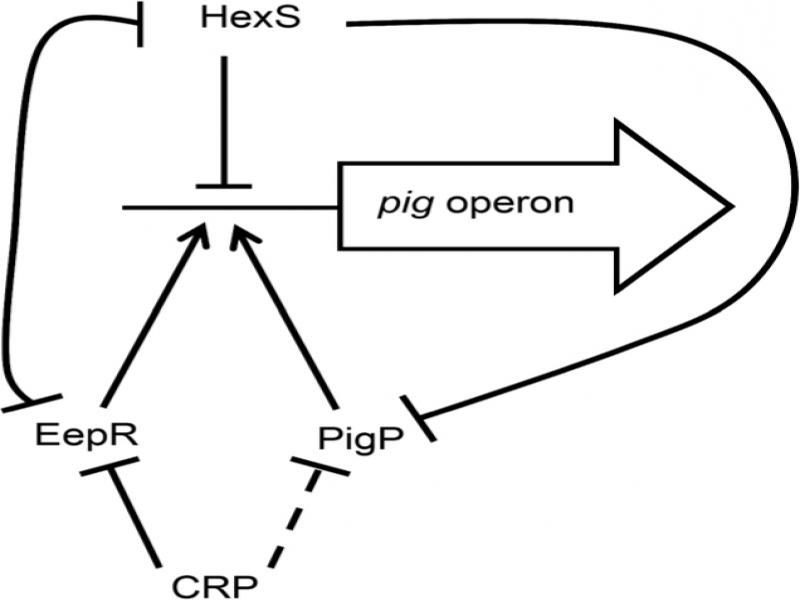
Fig. 5.
Strains and plasmids used in this study_
| Strain | Description | Source |
|---|---|---|
| PIC3611 | Wild-type Serratia marcescens | Presque Isle Cultures |
| CMS1687 | PIC3611 crp-Δ4 mutant | (Kalivoda et al. 2010) |
| CMS2097 | PIC3611 eepR mutant | (Stella et al. 2015) |
| CMS2922 | PIC3611 hexS mutant | (Shanks et al. 2013) |
| CMS4891 | PIC3611 pigB::tn-lux pigment operon reporter | this study |
| CMS4892 | PIC3611 pigF::tn-lux pigment operon reporter | this study |
| Plasmid | Relevant information | Source |
| pMQ99 | Plac-hcred shuttle vector | this study |
| pMQ132 | pBBR1-based shuttle vector | (Shanks et al. 2009) |
| pMQ200 | oriR6K-based suicide plasmid with PBAD | (Shanks et al. 2009) |
| pMQ221 | pMQ132 with pigP gene | (Shanks et al. 2013) |
| pMQ262 | pMQ200 with pigAB’ | (Kadouri and Shanks, 2013) |
| pMQ364 | pMQ132 with eepR gene | (Stella et al. 2015) |
| pMQ690 | promoterless-luxCDABE transposon delivery plasmid | this study |
| pMQ713 | pig promoter-luxCDABE transcriptional fusion | this study |